Онтогенетическая структура и жизненные стратегии ценопопуляций Delphinium pubiflorum

Автор: Шилова И.В., Кашин А.С., Богослов А.В., Пархоменко А.С.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Оригинальные статьи
Статья в выпуске: 4-1 т.27, 2018 года.
Бесплатный доступ
В статье приводятся данные по изучению онтогенетической структуры шести ценопопуляций Delphinium pubiflorum (DC.) Turcz. ex Huth: трех - из Саратовской, трех - из Ульяновской областей. Все популяции являются нормальными неполночленными, дефинитивными. Онтогенетические спектры всех изученных ценопопуляций симметричные, в большинстве случаев с одним максимумом на зрелых генеративных особях. Базовый спектр - центрированный, с максимумом на зрелых генеративных особях. Четыре из шести популяций D. pubiflorum оказались зрелыми, одна была зреющей, ещё одна со временем поменяла своё состояние со стареющей на зрелую. Ни одна из популяций в современных условиях не способна к самовосстановлению, что требует неотложных мер по их охране и восстановлению численности. Для морфометрических параметров отмечены следующие тактики и их сочетания: конвергентная тактика, дивергентно-конвергентная, конвергентно-дивергентная, тактика неопределённого варьирования. Для устойчивого существования в переменчивых условиях среды D. pubiflorum выработал стресс-толерантно-рудеральный тип стратегии.
Онтогенетическая структура, онтогенетические тактики, жизненные стратегии
Короткий адрес: https://sciup.org/148315174
IDR: 148315174 | УДК: 581.9 | DOI: 10.24411/2073-1035-2018-10089
Текст научной статьи Онтогенетическая структура и жизненные стратегии ценопопуляций Delphinium pubiflorum
Одной из главных диагностических характеристик в оценке общего состояния популяций является оценка их онтогенетического состояния. Важно выяснить, способны ли ценопопу-ляции (ЦП) редкого вида к самоподдержанию.
В последнее время активно изучаются тактики и стратегии жизни растений в условиях стресса (Злобин, 1989; Ишбирдин, Ишмуратова, 2004; Ишбирдин и др., 2005). Ю.А. Злобин (Злобин, 1989; Злобин и др., 2013) различает онтогенетические тактики структурных частей растения и растения в целом в течение онтогенеза. В то же время ряд авторов исследуют вариабельность морфологических признаков в популяциях различных видов растений при экологических и ценотических стрессах. Так А.Р. Ишбирдин и М.М. Ишмуратова (2004) рассматривают онтогенетические тактики структурных частей и растения в целом на экологическом градиенте.
Набор онтогенетических тактик является конкретным выражением параметров индивидуально-физиологического аппарата, поддерживающего популяцию вида в оптимальном для данного фитоценоза состоянии. Разнообразие форм онтогенетических тактик позволяет видам растений оптимизировать условия жизнедеятельности отдельных особей в одних эко-лого-ценотических условиях путём процесса их дифференциации, а в других – путём унификации (Злобин, 1989). Оценка онтогенетической тактики исключительно важна для понимания адаптируемости растений к условиям обитания. Использование концепции онтогенетических тактик для анализа редких видов растений дает полезную информацию для оценки устойчивости их популяций (Злобин и др., 2013).
Одной из ключевых задач популяционной ботаники является оценка стратегий жизни растений. Особенно важно изучение стратегий жизни редких видов растений.
Живокость пушистоцветковая ( Delphinium pubiflorum (DC.) Turcz. ex Huth) – редкий эндемичный вид Центрального (юго-восток Волжско-Донского подрайона) и Восточного (север Нижне-Донского и запад Заволжского подрайонов) флористических районов Восточной Европы (Цвелёв, 2001). Известны современные сборы из Ульяновской (Сенатор и др., 2016), Волгоградской (Ткаченко, 2017) и Саратовской (Шилова и др., 2016) областей.
Цель данной работы - определить состояние и перспективы существования ЦП D. Pubiflo-rum в Нижнем и Среднем Поволжье. Для этого были изучены: онтогенетическая структура ЦП, онтогенетические тактики морфометрических параметров, эколого-ценотическая и онтогенетическая стратегии данного вида.
МАТЕРИАЛЫ И МЕТОДЫ
Изучены шесть ЦП D. pubiflorum . Три из них произрастают на территории Саратовской обл.: в Красноармейском (Krm), Татищевском (Tat), Хвалынском (Hvl) р-нах, три – в Ульяновской обл.: две – в Радищевском (Gre, Bel), одна – в Новоспасском (Nov) р-нах. ЦП Tat изучалась с 2013 по 2017 гг., Hvl – с 2015 по 2017 гг., Krm – в 2015 и 2017 гг. Популяции из Ульяновской обл. исследованы в 2017 г.
Возрастные состояния выделялись по общепринятым методикам, исходя из размеров и числа вегетативных и генеративных органов без уничтожения растений (Заугольнова и др., 1988). В описании возрастных состояний использовали общепринятую классификацию возрастных групп: проростки (p), ювенильные (j), имматурные (im), виргинильные (v), молодые генеративные (g 1 ), зрелые генеративные (g 2 ), старые генеративные (g 3 ), субсенильные (старые вегетативные, ss), сенильные (отмирающие, s) (Злобин, 1989). Возрастные состояния генеративных растений выделяли согласно рекомендациям Н.И. Фёдорова (2003): g 1 – растения с одним цветущим побегом и ещё не пар-тикулировавшимся стеблекорнем; g2 – растения, имеющие только генеративные побеги и частично партикулировавшийся стеблекорень; g3 – растения с генеративными и вегетативными (на части отделяющихся партикул) побегами. Распределение особей по возрастным группам представлены в виде онтогенетических спектров, отражающих процентное участие особей различных возрастных групп в сложении популяции. На основе онтогенетических спектров ЦП, изученных за период с 2013 по 2017 гг., построен базовый спектр (Заугольнова и др., 1988).
Важной характеристикой динамичности или стабильности возрастной структуры ценопопу-ляций является индекс восстановления ( I восст ). Его определяли по формуле, рекомендованной Л.А. Жуковой (1987). I восст характеризует отношение совокупности всех особей прегенератив-ного периода к совокупности всех особей генеративного периода развития. Лишь при I восст >1 популяция способна к восстановлению. Кроме того, рассчитывали разработанные И.Н. Коваленко индексы, характеризующие участие отдельных онтогенетических групп особей в сложении популяции: индекс возобновляемости
( I возоб , характеризует участие совокупности всех особей прегенеративного периода), индекс гене-ративности ( I генер , участие совокупности всех особей генеративного периода развития), индекс старения ( I стар , участие совокупности всех растений от старых генеративных до сенильных), индекс общей возрастности ( I возр , характеризует отношение I стар / I возоб ) (Злобин и др., 2013).
Для характеристики онтогенетической структуры ЦП рассчитывали коэффициент воз-растности (Δ) (Уранов, 1975) и среднюю энергетическую эффективность популяции (ω). Тип ЦП определяли по классификации «дельтаомега» (Животовский, 2001).
Для того, чтобы определить онтогенетические тактики морфометрических параметров и стратегию D. pubiflorum у зрелых генеративных растений измерялись: высота (H) и диаметр (D) куста, длина побега (L), длина междоузлия (l), диаметр стебля (d) у его основания, длина (L Fol ) и ширина (W Fol ) листовой пластинки, длина (L Fl ) и ширина (W Fl ) цветка, длина шпорца (Cr), длина (L Pt ) и ширина (W Pt ) чашелистика; подсчитывалось число побегов (N) и число листьев (N Fol ) на побеге. По размерным спектрам составляющих ценопопуляции особей для оценки жизненности был рассчитан индекс виталитета популяций (IVC). Координация индекса IVC от минимального до максимального составила ряд улучшения условий произрастания растений (экоклин). На экоклине оценивалось изменение уровня варьирования каждого изученного нами признака (Ишбирдин и др. 2005). В этом ряду нами отмечены следующие тактики и их сочетания: конвергентная – коэффициент вариации признака (CV) при ухудшении условий (нарастании стресса) снижается; конвергентнодивергентная – при ухудшении условий CV вначале снижается, а при резком стрессе возрастает; дивергентно-конвергентная – при ухудшении условий CV вначале возрастает, а при резком стрессе снижается; тактика неопределённого варьирования, когда при нарастании стресса варьирование признака неоднократно то возрастает, то снижается.
По соотношению IVC max к IVC min , был определён индекс размерной пластичности: ISP= IVC max / IVC min (Ишбирдин и др., 2005)
Эколого-ценотическая стратегия D. pubiflorum определена по известной системе Раменского-Грайма (Миркин, Наумова, 1998), по которой стратегия вида определяется как место в системе независимых и определяющих выживание организмов факторов – стресса и нарушения. По соотношению потенциальных и реализованных значений морфометрических параметров растений в условиях максимального и минимального значения этих факторов выделяют три первичных типа стратегии – C (ви-оленты), S (патиенты) и R (эксплеренты), стоящих на вершинах «треугольника Грайма», а также переходные между ними.
Определение онтогенетической стратегии выживания вида проведено по характеру изменения морфологической целостности растений, оцениваемой по коэффициенту детерминации признаков (как среднего значения квадратов коэффициентов попарной корреляции всех признаков – R2m) на экоклине (Ишбирдин, Ишмуратова, 2004). По характеру изменения морфологической целостности растений в условиях нарастания стресса выделяют четыре типа онтогенетических стратегий растений:
-
1. защитная – с усилением стресса происходит усиление координации развития растений (повышается морфологическая целостность растения);
-
2. стрессовая – с усилением стресса ослабляется координация развития (снижается морфологическая целостность растения);
-
3. защитно-стрессовая – при нарастании стресса происходит сначала усиление, а затем ослабление координации развития растений (Ишбирдин, Ишмуратова, 2004).
-
4. стрессово-защитная – при нарастании стресса сначала происходит ослабление, а затем усиление координированности развития (чередование стрессовой и защитной компонент).
Для каждого параметра определялось среднее арифметическое значение, ошибка среднего арифметического, среднее квадратичное отклонение. Для расчётов применялись компьютерные программы: Microsoft Office Excel, Statistica 6.0, Past 3.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
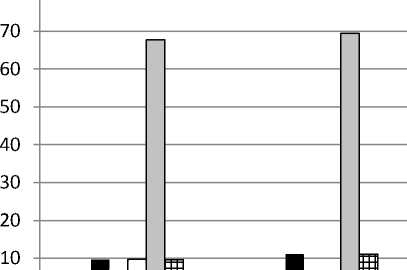
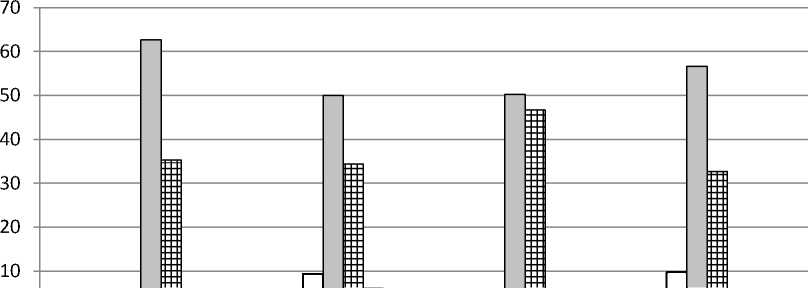
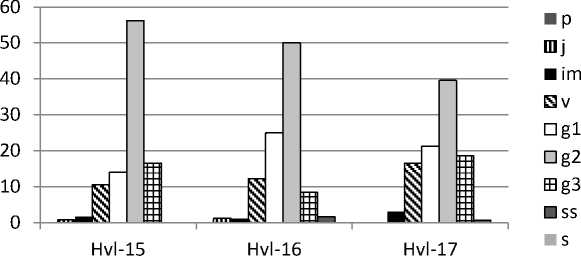
Онтогенетическая структура . Анализ соотношения групп особей разных онтогенетических состояний показал, что в большинстве изученных популяций D. pubiflorum постоянно преобладала группа зрелых генеративных растений (от 40 до 69%), особенно выраженная в ЦП Krm (рис. 1). В двух ЦП из Ульяновской обл. преобладали молодые генеративные особи. Второе место (33-47%) в онтогенетическом спектре в ЦП Tat с 2013 по 2016 гг. принадлежало группе старых генеративных особей. В этот период в данной ЦП молодые генеративные особи либо совсем отсутствовали, либо составляли от 3 до 10%. В 2017 г. в ЦП Tat
А
С
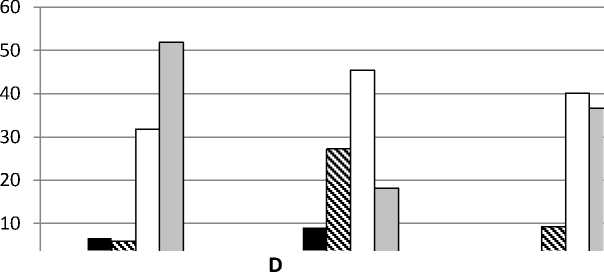
Рис. 1. Онтогенетические спектры ЦП Delphinium pubiflorum : A – ЦП из Красноармейского, B – ЦП из Татищевского, C – ЦП из Хвалынского р-нов Саратовской обл.; D – ЦП из Радищевского (Gre, Bel) и Новоспасского (Nov) р-нов Ульяновской обл. По оси абсцисс – условные обозначения ЦП и год исследования, по оси ординат – доля онтогенетической группы, %
резко снизилось доля старых генеративных и возросла доля молодых генеративных особей. В ЦП Krm в разные годы второе место делили группы имматурных, молодых и старых генеративных особей (по 10-11%). В ЦП Hvl в период с 2015 по 2017 гг. на второе место выходили группы то молодых, то старых генеративных особей, весьма заметное участие в онтогенетической структуре принимала группа вирги-нильных особей. В ЦП из Ульяновской обл. на втором месте были либо молодые (ЦП Gre), либо зрелые генеративные (ЦП Nov), либо вир-гинильные (ЦП Bel) особи. Группы особей пре-генеративного возрастного состояния практически отсутствовали в ЦП Tat, за исключением минимального участия виргинильных особей в 2016 и 2017 гг. В остальных изученных ЦП были отмечены более или менее значительные группы имматурных и виргинильных особей. Субсенильные особи в небольшом числе (1.56.2 %) обнаруживались в ЦП Tat, Hvl, Gre и Nov. Ювенильные особи наблюдались лишь в ЦП Hvl и Nov (около 1%). Сенильных особей и проростков зафиксировать не удалось. Возможно, отсутствие проростков связано с большой густотой травяного покрова в изученных сообществах, препятствующей прорастанию семян и развитию проростков. В пользу этого предположения свидетельствует то, что ювенильные и имматурные особи были обнаружены лишь на свободных от других травянистых растений небольших участках под кустарника- ми. Наиболее плотный травяной покров развился в сообществе с ЦП Tat. Именно в этой ЦП практически отсутствовали прегенератив-ные онтогенетические группы. Отсутствие проростков может быть связано и с тем, что популяции нами изучались в июле, и к этому времени проростки могли перейти в ювенильное состояние или погибнуть.
Все изученные популяции являются нормальными неполночленными. Популяции из Саратовской обл., изученные в течение нескольких лет, являются дефинитивными.
Онтогенетические спектры всех изученных ЦП симметричные, в большинстве случаев с одним максимумом на зрелых генеративных, в двух ЦП из Ульяновской обл. – на молодых генеративных особях (рис. 1). В ЦП Krm наблюдался ещё один небольшой пик на имма-турных особях. Базовый онтогенетический спектр (рис. 2) – центрированный симметричный, с максимумом на зрелых генеративных особях. Характер базового спектра определяется биологическими свойствами вида, а вариации в пределах зоны спектра – пластичностью реакции вида при воздействии различной экологической и ценотической обстановки (За-угольнова и др., 1988).
Участие отдельных онтогенетических групп в сложении популяции уточняют индексы, рассчитанные по рекомендациям И.Н. Коваленко (Злобин и др., 2013) и Л.А. Жуковой (Жукова, 1987) (таблица).
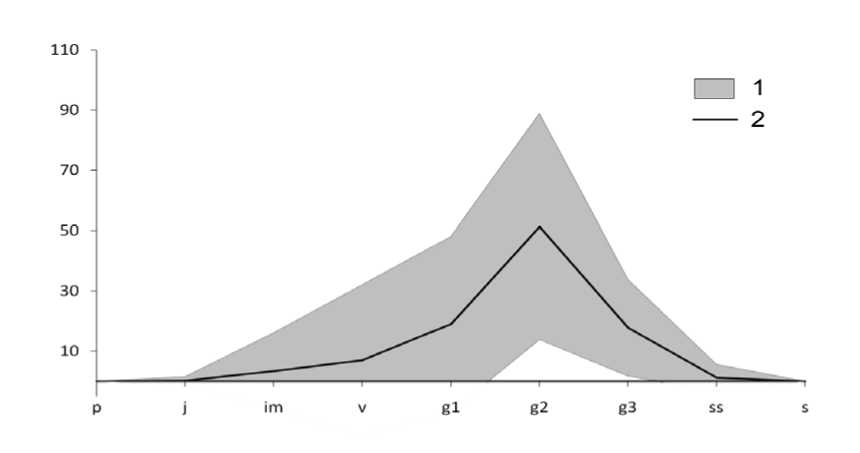
Рис. 2. Базовый онтогенетический спектр Delphinium pubiflorum : 1 – зона базового спектра;
2 – базовый спектр. По оси абсцисс – онтогенетические состояния особей: p – проростки, j – ювенильные, im – имматурные, v – виргинильные, g 1 – молодые генеративные, g 2 – зрелые генеративные, g 3 – старые генеративные, ss – субсенильные, s – сенильные растения.
По оси ординат – доля особей отдельных онтогенетических состояний, %
Данные, приведённые в таблице, свидетельствуют об очень высоком индексе генеративно-сти во всех изученных популяциях. Довольно высокий индекс старения отмечен в ЦП Tat, а в 2017 г. – в Hvl. Однако в ЦП Hvl в 2017 г. заметно повышались индекс возобновления и индекс восстановления. Выше эти индексы были только в ЦП Bel. Однако и эта ЦП не способна к самовосстановлению, о чём свидетельствует её I восст , не достигающий 1. Наиболее высоким
I возр отличалась ЦП Tat, низкие значения I возр были в ЦП Krm, Hvl, Gre, Nov. В ЦП Bel индекс общей возрастности равнялся нулю, поскольку старых особей здесь не обнаружено. По критерию «дельта-омега» большинство популяций D. pubiflorum оказались зрелыми (таблица). ЦП Bel была зреющей, а ЦП Tat со временем поменяла своё состояние со стареющей на зрелую.
Таблица
Индексы возрастного состояния и возрастность ценопопуляций Delphinium pubiflorum из Саратовской и Ульяновской областей
|
ЦП |
Год |
I возоб * |
I генер * |
I стар * |
I возр * |
I восст ** |
Δ*** |
ω **** |
Возрастность |
|
Krm |
2015 |
12.90 |
87.10 |
9.68 |
0.75 |
0.15 |
0.444 |
0.861 |
зрелая |
|
2017 |
13.89 |
86.11 |
11.11 |
0.80 |
0.16 |
0.452 |
0.857 |
зрелая |
|
|
Tat |
2013 |
0.001 |
98.04 |
37.25 |
37254.90 |
0.00 |
0.589 |
0.913 |
стареющая |
|
2014 |
0.001 |
93.75 |
40.62 |
40625.00 |
0.00 |
0.582 |
0.870 |
стареющая |
|
|
2015 |
0.001 |
100 |
46.71 |
46706.59 |
0.00 |
0.601 |
0.894 |
стареющая |
|
|
2016 |
0.88 |
99.12 |
32.74 |
37.00 |
0.01 |
0.549 |
0.904 |
зрелая |
|
|
2017 |
1.88 |
98.12 |
6.88 |
3.67 |
0.02 |
0.434 |
0.905 |
зрелая |
|
|
Hvl |
2015 |
13.16 |
86.84 |
16.67 |
1.27 |
0.15 |
0.454 |
0.851 |
зрелая |
|
2016 |
14.83 |
83.47 |
10.17 |
0.68 |
0.18 |
0.410 |
0.825 |
зрелая |
|
|
2017 |
19.77 |
79.46 |
19.38 |
0.98 |
0.25 |
0.419 |
0.788 |
зрелая |
|
|
Gre |
2017 |
12.59 |
85.92 |
3.70 |
0.29 |
0.15 |
0.385 |
0.823 |
зрелая |
|
Bel |
2017 |
36.36 |
63.64 |
0.00 |
0.00 |
0.57 |
0.250 |
0.670 |
зреющая |
|
Nov |
2017 |
11.05 |
85.46 |
12.21 |
1.10 |
0.13 |
0.398 |
0.806 |
зрелая |
Примечание: *– индексы по И.Н. Коваленко, ** – индекс по Л.А. Жуковой, *** – коэффициент возраст-ности по А.А. Уранову, **** – коэффициент эффективности и возрастность по Л.А. Животовскому, ***** – возрастность по критерию «дельта-омега».
Онтогенетические тактики . Как установила В.Н. Сулейманова (Злобин и др., 2013), разные структурные части растения могут реализовать разные типы онтогенетической тактики. Выявленная закономерность отражает модульную организацию растительных организмов с выраженной морфогенетической самостоятельностью отдельных модулей. Также разную направленность могут иметь тактики вегетативных и генеративных структур.
На экоклине, установленном по IVC, у D. pubiflorum нами отмечены следующие онтогенетические тактики и их сочетания. Конвергентная тактика отмечена лишь у W Fol (рис. 3 а); д ивергентно-конвергентная - у L Fl (рис. 3 б); конвергентно-дивергентная – у H, D, L, l, L Fol , W Fl , Cr (рис. 4). Тактика неопределённого варьирования выявлена у пяти параметров: N, d, N Fol , L Pt , W Pt (рис. 5).
При резком ухудшении условий особи становятся низкими, листья их – узкими. При этом варьирование ширины листа резко снижается, а 45
длины листа при умеренном стрессе увеличивается и лишь при резком ухудшении условий стабилизируется, чем обеспечивается устойчивость такого важного показателя, как площадь листа, и снабжение пластическими веществами генеративных органов. При нарастании стресса варьирование длины и толщины стебля, числа побегов и листьев, диаметра куста попеременно то усиливается, то ослабевает. Таким путём ведётся поиск ниши для выживания.
В генеративной сфере при умеренном стрессе с увеличением ширины цветка и чашелистика, длины чашелистика и шпорца вариабельность этих признаков вначале уменьшается, но при нарастании стресса варьирование ширины цветка резко возрастает, длины цветка резко снижается, а остальных признаков, увеличившись, в какой-то мере стабилизируется. Для насекомоопыляемого растения, каковым является дельфиниум, вполне объяснимо при стрессе увеличение параметров цветка, что делает его более заметным для опылителей. Увеличе- ние варьирования параметров цветка, в том числе шпорца, содержащего нектар, способствует привлечению как можно более разнообразных насекомых-опылителей. Сочетанием
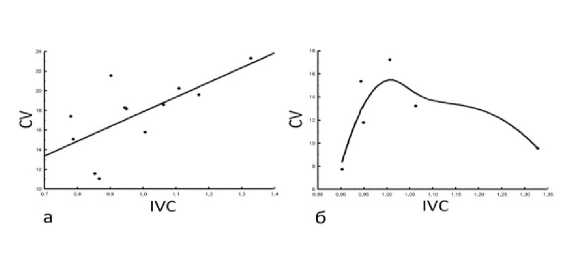
Рис. 3. Конвергентная (а) и дивергентно-конвергентная (б) онтогенетические тактики морфометрических параметров D. pubiflorum : а – ширины листовой пластинки, б – длины цветка
конвергентной и дивергентной тактик достигается цель завязать семена и дать потомство в стрессовых условиях.
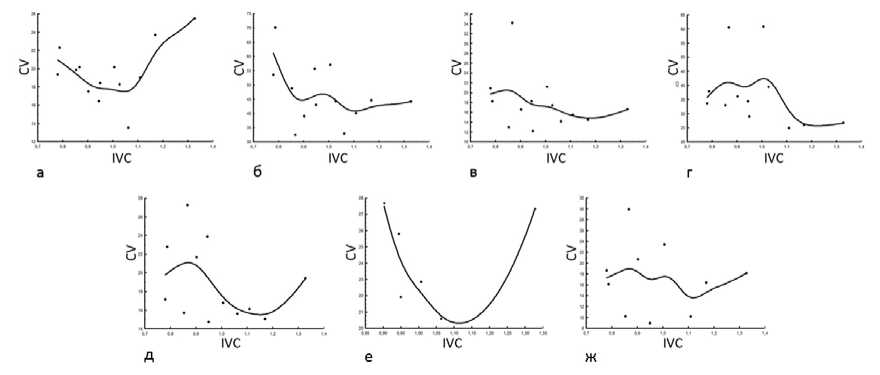
Рис. 4. Конвергентно-дивергентная онтогенетическая тактика морфометрических параметров D. pubiflorum : а – высоты растения, б – диаметра куста, в – длины генеративного побега, г – длины междоузлия, д – длины листовой пластинки, е – ширины цветка, ж – длины шпорца
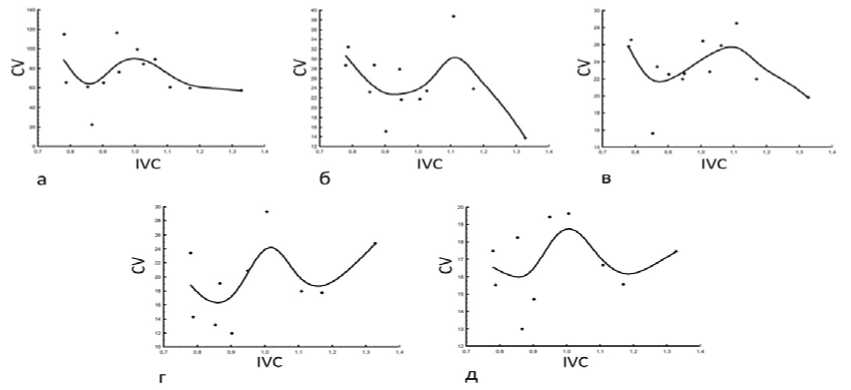
Рис. 5. Тактика неопределённого варьирования морфометрических параметров D. pubiflorum : а – числа генеративных побегов, б – диаметра стебля, в – числа листьев на побеге, г – длины чашелистика, д – ширины
чашелистика
Эколого-ценотическая стратегия D. pubiflorum . По нашим наблюдениям, D. pubiflorum присуща SR-стратегия (стресс-толерантно-рудеральная, или – патиентно-эксплерентная). Экотопическая и фитоценоти-ческая патиентность (S-стратегия) проявляется в избирательности субстрата (карбонатные почвы), тяготении к кустарниковым сообществам. При неблагоприятных условиях происходит уход от конкуренции за счёт перехода в покой. Так, в 2014 г. нами было отмечено, что часть растений в популяции Tat находилась в состоянии подземного покоя. Именно в 2014 г. в данной ЦП виталитет был самым низким (рис. 6). Уход в покой при сильном стрессе наблюдали и у других редких растений, в частности, орхидей (Ишбирдин и др., 2005).
R-стратегия просматривается в следующем. В благоприятных условиях, при увеличении параметров вегетативной сферы, происходит стабилизация (снижение варьирования) как вегетативной сферы (диаметра куста, длины и толщины стебля, числа листьев), так и генеративной сферы (числа генеративных побегов, их длины, включая и длину соцветия, длины цветка), тем самым создаются условия для увеличения семенной продуктивности. При произрастании D. pubiflorum в сообществах с густым травянистым покровом, молодые особи обнаруживались на прогалинках под кустарниками. В случае нарушения травяного покрова (пожар, порои животных и пр.) растения могут успешно засеять субстрат и дать потомство.
Как указано выше, конвергентнодивергентная тактика характерна для семи изученных признаков. Кроме того, в тактике неопределённого варьирования, отмеченной у пяти признаков, при резком стрессе явно преобладает дивергентная составляющая. Другие исследователи также отмечают, что в природных условиях преобладает тактика дивергенции, и подчёркивают, что особенно она характерна для R-стратегов. Это приводит к внутрипопуляцион-ному разнообразию растений по размеру, форме и свойствам и в конечном итоге обеспечивает более полное использование ресурсов, снижая конкуренцию между особями растений одной популяции. Полезна тактика дивергенции у растений с энтомофильными цветками, так как при расположении соцветий на разной высоте и при цветении растений в разное время снижается конкуренция за опылителей. В целом, тактика дивергенции повышает стабильность и устойчивость популяции (Злобин и др., 2013).
Онтогенетическая стратегия . У D. pubiflorum наблюдается чередование защитной и стрессовой компонент в онтогенетической стратегии (рис. 6).
Такой тип зависимости характерен для видов, проявляющих патиентность (S-стратеги). При нарастании стресса у D. pubiflorum происходит усиление детерминированности в развитии морфоструктур. Как отмечалось выше, в ЦП Tat, изучавшейся на протяжении пяти лет, наихудшее развитие растений было отмечено в 2014 г., в это время коэффициент детерминации в этой ЦП имел максимальное значение (рис. 6). При сильном стрессе ослабевает координированность развития. Так, ЦП Gre и Nov, имеющие самый низкий индекс виталитета популяции (рис. 6), были определены как депрессивные. Эти популяции находятся на северной границе ареала, а виды на границе ареала всегда испытывают стресс. Размерная пластичность у патиентов, как правило, невысокая. У D. pubiflorum ISP=1.70.
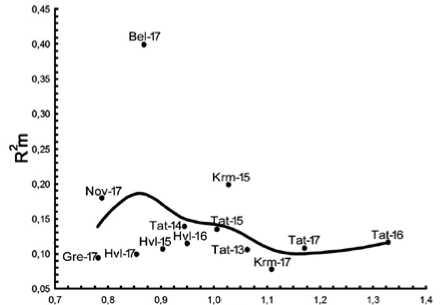
IVC
Рис. 6. Изменение морфологической целостности D. pubiflorum в ряду изменения эколого-ценотических условий. По оси абсцисс индекс виталитета ценопопуляций (IVC), по оси ординат – индекс морфологической целостности особей (коэффициент детерминации признаков, R2m)
ЗАКЛЮЧЕНИЕ
В подавляющем большинстве изученных ценопопуляций D. pubiflorum преобладает группа зрелых генеративных особей. Все популяции являются нормальными неполночленными и, судя по популяциям, изученным в течение нескольких лет, дефинитивными. Онтогенетические спектры всех изученных ЦП симметричные, в большинстве случаев с одним максимумом на зрелых генеративных особях. Базовый спектр – центрированный, с максимумом на зрелых генеративных особях. Четыре из шести популяций D. pubiflorum оказались зрелыми. Одна ЦП была зреющей, ещё одна ЦП со временем поменяла своё состояние со стареющей на зрелую.
Ни одна из популяций в современных условиях не способна к самовосстановлению, что требует неотложных мер по их охране и восстановлению численности.
Для морфометрических параметров отмечены следующие онтогенетические тактики и их сочетания. Конвергентная тактика отмечена лишь у ширины листовой пластинки; д ивергентно-конвергентная – у длины цветка; конвергентно-дивергентная – у высоты растения,
Список литературы Онтогенетическая структура и жизненные стратегии ценопопуляций Delphinium pubiflorum
- Животовский Л.А. Онтогенетические состояния, эффективная плотность и классификация популяций растений // Экология. 2001. № 1. С. 3-7.
- Жукова Л.А. Динамика ценопопуляций луговых растений в естественных фитоценозах / Динамика ценопопуляций травянистых растений. Киев: Наукова думка, 1987. С. 9-19.
- Заугольнова Л.Б., Жукова Л.А., Комаров А.С., Смирнова О.В. Ценопопуляции растений (очерки популяционной биологии). М., 1988. 184 с.
- Злобин Ю.А. Принципы и методы изучения ценотических популяций растений. Казань: Изд-во Казанского университета, 1989. 146 с.
- Злобин Ю.А., Скляр В.Г., Клименко А.А. Популяции редких видов растений: теоретические основы и методика изучения. Сумы: Университетская книга, 2013. 439 с.
