Онтогенез Eulophia streptopetalla Lindl. (Orchidaceae) в культуре
 в культуре")
Автор: Широков А.И., Сырова В.В., Максимов А.А., Макарова А.Е., Прокин А.М.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 3 т.19, 2025 года.
Бесплатный доступ
В работе представлены результаты исследования онтогенеза Eulophia streptopetalla Lindl. – редкого наземного вида орхидных, в условиях in vitro и ex situ. Выделены и подробно охарактеризованы стадии онтогенеза от протокорма до генеративного растения. Особое внимание уделено морфогенетическим особенностям ранних этапов развития и способности к вегетативному размножению на стадии проростка. Показано, что при культивировании на среде Harvais без добавления регуляторов роста возможно полноценное развитие растений, включая переход к цветению. После адаптации к условиям оранжереи сеянцы сохраняют характерные морфологические признаки и формируют цветоносы, что подтверждает завершение полного онтогенетического цикла. Результаты свидетельствуют о высоком потенциале данного вида для микроклонального размножения.
Eulophia streptopetalla, онтогенез, in vitro, ex situ, протокорм, орхидные, вегетативное размножение, адаптация, микроклональное размножение
Короткий адрес: https://sciup.org/148331560
IDR: 148331560 | УДК: 581.142:581.4 | DOI: 10.24412/2072-8816-2025-19-3-336-344
Ontogenesis of Eulophia streptopetalla Lindl. (Orchidaceae) in culture
This study presents the ontogenetic development of Eulophia streptopetalla Lindl., a rare terrestrial orchid species, under in vitro and ex situ conditions. The complete ontogenetic sequence – from protocorm to generative plant — is described in detail. Special emphasis is placed on the morphogenetic characteristics of early developmental stages and the ability of seedlings to undergo vegetative multiplication during the protocorm and seedling phases. It was demonstrated that cultivation on Harvais medium without growth regulators supports successful development, including flowering. After acclimatization in greenhouse conditions, plants retained typical morphological traits and produced inflorescences, confirming the completion of a full life cycle. The results highlight the high potential of this species for micropropagation.
Текст научной статьи Онтогенез Eulophia streptopetalla Lindl. (Orchidaceae) в культуре
Орхидные (Orchidaceae) представляют собой одно из самых многочисленных семейств цветковых растений, насчитывающее около 25–30 тысяч видов, большинство из которых обитает в тропических регионах (Chase et al., 2015; Vitt et al., 2023). Несмотря на широкое распространение, многие представители семейства находятся под угрозой исчезновения из-за утраты среды обитания, нелегального сбора и глобальных экологических изменений (Hussain et al., 2023). Представители рода Eulophia R.Br., насчитывающее более 200 видов (Chase et al., 2015), не являются исключением, они становятся редкостью, поскольку подвергаются чрезмерному сбору активно используются в традиционной медицине (Chinsamy et al., 2015; Nandgave et al., 2021).
Современные подходы к сохранению орхидных включают ex situ культивирование и размножение in vitro, позволяющие восстанавливать редкие виды и пополнять генофондовые коллекции ботанических садов (Tiwari et al., 2022). Для эффективного применения и оптимизации этих методов необходимы данные о стадиях онтогенеза, механизмах перехода между ними и условиях, способствующих успешной адаптации к нестерильной среде. Исследования по онтогенезу орхидей в асимбиотических условиях позволяют выявить характерные фазы морфогенеза и определить ключевые факторы, влияющие на развитие проростков и формирование генеративных особей.
Цель работы – выявление и описание стадий онтогенеза Eulophia streptopetalla в культуре (in vitro и ex situ).
М АТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Объект исследования.
Eulophia streptopetalla Lindl. – многолетнее наземное травянистое растение высотой 50–150 см. Псевдобульбы хорошо развиты, конически или цилиндрические, длиной 2,5–10 см, шириной – 1,7–2,5 см. Частично или полностью могут находиться под землей. Листья крупные в числе от 4 до 9, сидячие, ланцетные и ребристые, длиной 40–65 см, шириной – 4–8 см. Соцветие рыхлое многоцветковое 15–50 см высотой. Прицветники ланцетные или яйцевидные. Цветки, поникающие желто-коричневатых тонов. Губа трёхлопастная, 8–12 мм длиной и 5–11 мм шириной. Цветение с марта по май. Вид распространен по всей тропической и южной Африке, южной Аравии и в Йемене. Обычно встречается на опушках лесов, на каменистых известняковых склонах с редкими кустарниками, вдоль ручьев или на лугах от побережья до 2550 метров над уровнем моря (Demissew et al., 2004).
Согласно классификации форм роста, Е.С. Смирновой (Smirnova, 1990), E. streptopetalla относится к корневищному растению, на каждом побеге которого все междоузлия утолщены. У растений такого типа корневищный участок плохо выражен, такая особь воспринимается как группа тесно сомкнутых вертикальных побегов последовательных порядков ветвления. Большинство узлов побегов несут чешуевидные и влагалищные листья. Полноценные фотосинтезирующие листья располагаются в верхней части побегов и часто собираются в верхушечную розетку. Придаточные корни образуются исключительно в основании побегов.
Источником материала для исследования стали растения, полученные из семян, собранных в 2004 году в окрестностях города Морогоро (Танзания). Высота над уровнем моря около 1200 м. Популяция располагалась в долине горного ручья на каменной плите магматического происхождения, покрытой толстым слоем (более 20 см) сильно задерненного гумуса. Немногочисленные (не более 5 штук) куртины (включающие по 10–20 туберидиев) располагались среди дернины из лугового разнотравья. Зрелые семена были высеяны на стерильные питательные среды и через 8 лет были получены первые цветущие растения.
При произведении посевов in vitro использовались недозрелые коробочки от искусственного опыления, для стерилизации их погружали в 96% спирт на 2–3 минуты, затем прожигали в пламени. В стерильных условиях семена извлекали и помещали на питательную среду в колбы объемом 100 мл, предварительно стерилизованную автоклавированием при 120°C в течение 30 минут. Для предотвращения контаминации стаканы с посевами герметично закрывали пленкой. В качестве питательная среды использовалась среда Harvais (Harvais, 1982). Посевы содержались на стеллажах под люминесцентными лампами в условиях кондиционируемой камеры при температуре 20 ± 1°C, с фотопериодом 16 часов. После завершения выращивания растения высаживались из колб для адаптации в живой измельченный сфагнум. В течение 2–3 месяцев адаптация обычно проходит успешно и растения высаживаются в субстрат из смеси равных частей керамзитового песка, диатомита, древесного угля, агроперлита и сосновой коры. Все компоненты субстрата соответствуют фракции 0,5–0,7 см. Более взрослые растения пересаживаются в такой же по составу субстрат, но фракцию обычно увеличиваем до 0,8–1,0 см.
При описании онтогенеза нами использована популяционно-онтогенетическая концепция (Rabotnov, 1950; Uranov, 1967; Uranov,1974).
Р ЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
По результатам исследования нами были выделены следующие стадии онтогенеза E. streptopetalla :
Протокорм (pr). Под протокормом орхидных обычно понимают начальную стадию развития проростка, его формирование связано с прорастанием недифференцированного зародыша (Kulikov, 1998; Shirokov et al., 2011; Kolomeytseva et al., 2012). Понятие «прорастание» применительно к семенам орхидей весьма условно, так как в данном случае нет типичного «проклевывания» зародышевого корешка. Прорастание начинается с растрескивания семенной кожуры, небольшого высовывания набухшего тела зародыша наружу. Явное прорастание высеянных семян на питательной среде наблюдалось уже через 4-5 недель после посевов. Формирование полноценного протокорма (Рис. 1. pr) наблюдалось на 9 неделю развития. Он представляет собой шаровидное тело 0,2–0,3 см в диаметре. Протокорм E. streptopetalla c небольшим числом ризоидных волосков и окрашен в зеленый цвет, что свидетельствует о его способности к фотосинтезу.
Проросток (pl) . В литературе встречается много разногласий по вопросу о выделении ранних стадий онтоморфогенеза у орхидных (Kulikov, 1998; Shirokov et al., 2011; Kolomeytseva et al., 2012). Мы считаем началом стадии проростка закладка на верхушке протокорма почки с зачатками будущих листьев. Благодаря способности протокорма E. streptopetalla к фотосинтезу он начинает нарастать в горизонтальном направлении и формирует протокормоподобное тело зеленого цвета с выраженной верхушечной почкой (Рис. 1. pl (1). Именно так и выглядит проросток. В этом состоянии проросток затормаживает рост тела (размер не превышает 0,3–0,4 см в диаметре) и начинает продуцировать аналогичные протокормоподобные тела с выраженными верхушечными почками. В результате формируются агрегации (колонии по 5–12 шт.) протокормоподобных тел (Рис. 1. pl (2), которые разрастаются радиально. Затем мультипликация протокормоподобных тел останавливается, и они начинают увеличиваться в размерах, видимо, также за счет фотосинтеза. Их размер может достигать 1см (Рис. 1. pl (3). Формирование множества протокормоподобных тел на стадии проростка можно интерпретировать как раннюю форму вегетативного размножения. Аналогичные явления можно наблюдать и в условиях естественной среды (Shirokov et al., 2011; Kolomeytseva et al., 2012).
Ювенильное растение (j) . При культивировании in vitro сеянцы E. streptopetalla переходят в ювенильное состояние по прошествии 55-59 недель с начала культивирования. Переход в ювенильное состояние связан с началом автотрофного питания развития (Shirokov et al., 2011; Kolomeytseva et al., 2012) и характеризуется формированием на протокормоподобных телах зачатка вегетативного побега с первичным корнем. Такой побег, как правило, не превышает в длину 1,0–1,5 см и несет 2-4 чешуевидных, влагалищных листа (Рис. 1. j).
Ювенильно – имматурное растение (j-im) . Главным признаком перехода от ювенильного состояния к имматурному у орхидных в природных условиях является смена способа нарастания побега. Однако многолетние эксперименты показывают, что специфичные условия in vitro с выровненными температурным и световым режимами, а также особенности минерального питания могут изменять типичную картину онтогенеза орхидных и вызывать задержку развития (Shirokov et al., 2011; Kolomeytseva et al., 2012). Ювенильно-имматурное онтогенетическое состояние представляет собой переходный вариант, связанный с задержкой развития, по-видимому, связанную с сохранением связей с зародышевыми структурами, и продолжением мультипликации протокормоподобных тел особями, имеющими уже листостебельную форму. Переход от моноподиального нарастания побега в этом состоянии осуществился, но вместо свойственного виду членистого туберидия закладываются новые пртокормоподобные тела. При этом такие особи формируются, как правило, через 70 недель после посева и при дальнейшем культивировании их на стерильных питательных средах могут существовать неограниченно долго, особенно при внесении в среду гормональных веществ.
В j-im онтогенетическом состоянии (рис. 1. j-im) растение имеет один более-менее развитый влагалищный лист, с листовой пластинкой длиной 1,0–1,5 см и 2–4 чешуевидных, влагалищных листа. Трубка влагалища может достигать 3,0–3,5 см. Растущий побег имеет один хорошо развитый корень, запасающего типа, длиной 1,0–1,5 см.
Имматурное растение (im1) всё же может сформироваться на среде in vitro, после длительного пребывания в ювенильно-имматурном онтогенетическом состоянии. Такое растение имеет, как правило, развитое «членистое корневище» (ч.к.) из крупных протокормоподобных тел (по-видимому, это недоразвитые туберидии). На таком корневище хорошо заметны зачаточные, спящие вегетативные (в.п.), единичные корни, запасающего типа, достигающие в длину до 5,0 см. Сформировавшийся, в текущий период роста, побег несет два влагалищных листа с листовыми пластинками длиной 4,0–5,0 см и 4-5 чешуевидными влагалищными листьями. Размер растения (при позволяющем объеме лабораторной посуды – 0,3–0,5 л) может достигать в высоту более 10,0 см.
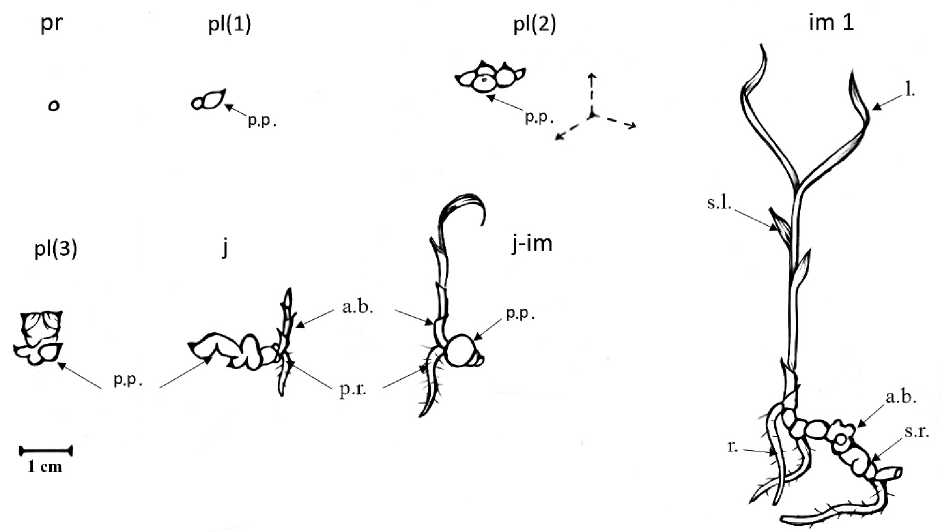
Рис. 1. Ранние стадии онтогенеза E. streptopetalla in vitro
Условные обозначения: pr – протокорм, pl – проросток, j – ювенильное растение, j-im – ювениьно-имматурное растение, im 1 – имматурное растение, p.p. – проткормоподобное тело, p.r. – первичный корень, a.b. – верхушечная почка, s.l. – чешуевидный лист, l. – лист, r. – корень, s.r. – членистое корневище
Fig. 1 . Early stages of ontogenesis of E. streptopetalla in vitro
Legend: pr – protocorm, pl – seedling, j – juvenile plant, j-im – juvenile-immature plant, im 1 – immature plant, p.p. – protocorm-like body, p.r. – primary root, a.b. – apical bud, s.l. – scale leaf, l. – leaf, r. – root, s.r. – segmented rhizome
Дальнейшее описание онтогенеза производилось нами после извлечения растений из асептических условий и адаптации сеянцев в условиях тропической оранжереи Ботанического сада Института биологии и биомедицины Нижегородского государственного университета им. Лобачевского.
Имматурное растение (im2). Растение имеет небольшие (до 4,0 см длиной и до 1,0 см в диаметре) сформированные членистые (2–4 узла) туберидии. Нижние листья, в числе 1–2, чешуевидные влагалищные, а верхние (2–3 листа) – так же влагалищные, имеют хорошо развитую, фотосинтезирующую листовую пластинку до 8.0 см длиной. Розетка верхушечных листьев выражена слабо из-за их небольшого числа. В этом состоянии наблюдается типичное для взрослого растения симподиальное нарастание побега, при котором развитие нового туберидия наблюдается из нижнего узла предыдущего туберидия, закрытого влагалищем нижнего чешуевидного листа. Из этого же узла развиваются почти не ветвящиеся белые придаточные корни с развитым слоем веламена.
Виргинильное растение (v) – имеет практически взрослый облик. Членистые туберидии несут от 3 до 6 узлов. Верхушечные ланцетовидные, влагалищные листья, в числе 3–5, имеют дину до 25,0 см и образуют ярко выраженную розетку. Нижние чешуевидные влагалищные листья в числе 1–2. Толстые, извилистые придаточные корни имеют почти белую окраску из-за хорошо выраженного веламена и образуются из нижнего узла побега.
Генеративное растение (g). Переход в генеративное состояние определяется началом цветения и плодоношения растения. Растения становятся более мощными. Членистые туберидии несут от 5 до 9 узлов. Верхушечные ланцетовидные, влагалищные листья, в числе 5– 8, имеют длину до 50,0 см (ширина до 5,0 см) и образуют крупную розетку. Нижние чешуевидные влагалищные листья в числе 2–4. Из нижнего узла побега, разрывая влагалище чешуевидного листа появляется многоцветковый цветонос, достигающий длины до 1,0 м. Цветонос несет до 30 желто-коричневых цветков до 1,5-2,5 см в диаметре. По мере разрастания особи и образования новых побегов у зрелых генеративных особей, в условиях оранжереи, образовывалось до 5 цветоносов.
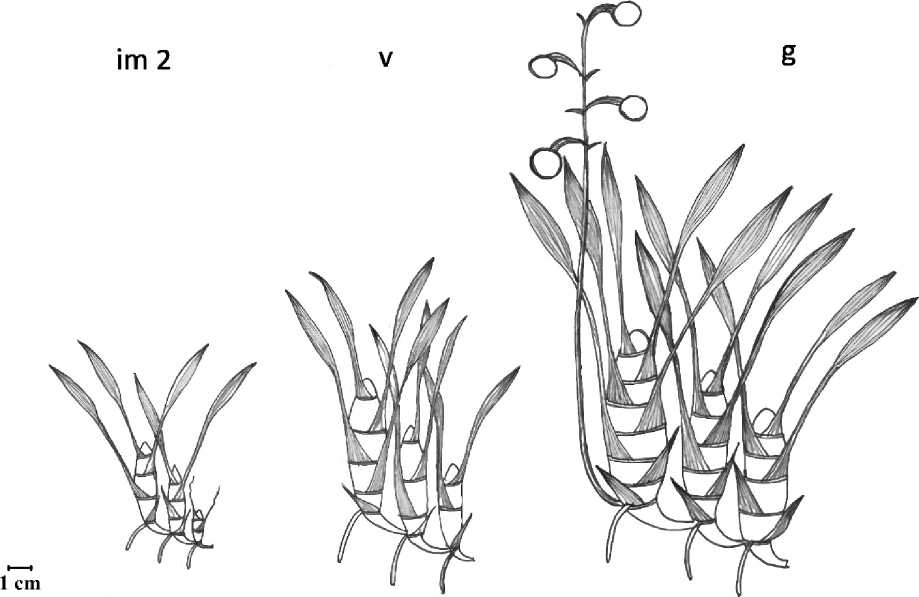
Рис. 2. Стадии онтогенеза E. streptopetalla в условиях оранжереи
Условные обозначения: im 2 – имматурное растение, v – виргинильное растение, g – генеративное растение
Fig. 2 . Stages of ontogenesis of E. streptopetalla under greenhouse conditions
Legend: im 2 – immature plant, v – virginal plant, g – generative plant
Выявленные особенности протокорма E. streptopetalla , в частности его фотосинтетическая активность, маленькое число ризоидов на стадии протокорма и формирование устойчивых агрегаций протокормоподобных тел на стадии проростка, что может рассматриваться как ранняя форма вегетативного размножения и указывает на способность переходить к автотрофному питанию уже на ранних этапах развития. Аналогичные структуры были отмечены у других представителей рода Eulophia , таких как E. flava (Lindl.) Hook.f. и E. alta (L.) Fawc. & Rendle, при культивировании in vitro (Vasupen et al., 2023; Johnson et al., 2007), а также у других орхидных (Yamazaki, 2006; Stewart, 2006). Эти особенности могут играть роль в повышении выживаемости и адаптации к асимбиотическим условиям культивирования in vitro.
Длительное пребывание E. streptopetalla в стадии j-im может быть объяснено отсутствием эндогенных стимулов или отсутствием определённых веществ в питательной среде, регулирующих переход к следующей стадии. Это явление описано и у других родов на разных стадиях, в частности Cymbidium и Paphiopedilum, где задержка развития наблюдалась в условиях дополнительной гормональной стимуляции (Deb, 2011; Xu et al., 2020). Однако устойчивость таких «застрявших» форм, продолжающих мультипликацию, может быть полезна для целей массового микроклонального размножения.
Формирование членистого корневища из протокормоподобных тел также наблюдалось у E. promensis Lindt., E. flava и E. bicallosa (D.Don) P.F.Hunt & Summerh, где такие структуры успешно приживались при пересадке в нестерильные условия (Hossain, 2015; Wongsa et al., 2025). Это указывает на то, что морфологически специфичные структуры in vitro могут обладать высокой способностью к прорастанию и дальнейшему развитию. Переход E. streptopetalla к генеративному состоянию после адаптации ex vitro, с формированием цветоносов и цветков, произошел позднее по сравнению с некоторыми другими видами рода, где генеративные растения получались в течение 1-3 лет (Hossain, 2015; Chang et al., 2015; Panwar, 2022).
З АКЛЮЧЕНИЕ
-
E. streptopetalla проходит в условиях культуры полный цикл развития, от посева семян до формирования генеративных особей, формирующих цветки и плоды. Особенностью онтогенеза является формирование фотосинтетически активных агрегаций протокормоподобных тел на, что свидетельствует о высокой адаптивности и способности к вегетативному размножению. Эти структуры сохраняют потенциал даже при длительном пребывании на переходной стадии (j-im), что делает их перспективными для микроклонального размножения.
Развитие растений после пересадки в условия оранжереи показало сохранение морфологических признаков, характерных для природной формы. Способность растений формировать до пяти цветоносов открывает перспективы как для коммерческого культивирования, так и для участия в восстановительных программах.
В дальнейшем представляется целесообразным оптимизировать состав питательной среды для более эффективного и быстрого развития на первых стадиях, а также исследовать влияние фитогормонов на онтогенез. Результаты могут быть использованы при разработке устойчивых протоколов in vitro для размножения редких орхидей и введения в культуру других видов рода Eulophia .
