Онтогенез и динамика популяций остролодочника колосистого (Oxytropiss picata (Pall.) O. et B. Fedtsch., Fabaceae) в Самарской области
 O. et B. Fedtsch., Fabaceae) в Самарской области")
Автор: Ильина В.Н.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 2 т.26, 2017 года.
Бесплатный доступ
В статье приведены многолетние данные по онтогенезу и структуре природных популяций редкого в самарской флоре Oxytropis spicata (Pall.) O. et B. Fedtsch. ( Fabaceae ). Дано предварительное описание стадий онтогенеза и определена их длительность. Проанализированы данные по распространению вида в области.
Природоохранный статус, онтогенез, популяция, численность, возобновление, виталитет, антропогенные факторы, красная книга, самарская область
Короткий адрес: https://sciup.org/148315046
IDR: 148315046 | УДК: 581.
Текст краткого сообщения Онтогенез и динамика популяций остролодочника колосистого (Oxytropiss picata (Pall.) O. et B. Fedtsch., Fabaceae) в Самарской области
Остролодочник колосистый ( Oxytropis spicata (Pall.) O. et B. Fedtsch., Fabaceae ) – субэндемик бассейна р. Волги (Васюков и др., 2015), представляет собой травянистый стержнекорневой бесстебельный многолетник из секции Orobia . Семена обладают твердой водонепроницаемой кожурой и могут длительно сохранять всхожесть (Ели-зарьева, Кунакасова, 2013). В связи с этим в почве возможно накопление банка семян, что важно для возобновления популяций и переживания неблагоприятных условий существования в латентном периоде.
Он был включен в Красную книгу Самарской области (далее СО) со статусом редкости (3/А) – крайне редкий вид, резко снижающий свою численность (Красная книга Самарской области, 2007). В сопредельных регионах O. spicata занесен в списки охраняемых растений в Республики Татарстан и Саратовской области.
В СО вид отмечен в Заволжье. В последние годы ботаниками региона вид зафиксирован в различных районах области. Алексеевский : Родник истока р. Съезжей (Реестр …, 2010). Большечерниговский : ПП «Грызлы» (Реестр …, 2010), ПП «Попов сад» (Реестр …, 2010), ПП «Балка Кладовая» (Реестр …, 2010); ПП «Истоки реки Большой Иргиз» (Реестр …, 2010), ПП «Кошкинская балка» (Реестр …, 2010; Ильина, Митрошенкова, 2014, 2015); ПП «Сестринские окаменелости» (Реестр …, 2010), ПП «Урочище Мулин дол» (Реестр …, 2010); ПП «Участок типчаково-ковыльной степи» (Реестр …, 2010). Елховский : ПП «Гора Зеленая» (Реестр …, 2010). Исаклинский : окр. с. Исаклы (Иванова, Елкина, 2008; Саксонов, Сенатор и др., 2008); ПП «Исаклин-ская нагорная лесостепь» (Моисеева, Ильина, 2009; Реестр …, 2010); гора Пионерка (Ильина, 2006 б, 2010, 2014, 2015а; Шаронова, Курочкин, 2014; Ильина, Митрошен-кова, 2015); окр. с. Сухарь Матак, левый берег р. Шунгут (Шаронова, Курочкин, 2015). Камышлинский : ПП «Новоусмановская сероводородная вода» (Корчиков и др., 2011); окр. с. Стар. Ермаково (Шаронова, Курочкин, 2015); Овраг Куркульный (Шаронова, Курочкин, 2015). Кинельский : ПП «Алакаевско-Чубовская каменистая степь» (Реестр …, 2010; Ильина, Митрошенкова, 2015), ПП «Чубовская степь» (Красная книга…, 2007; Реестр …, 2010; Ильина, Митрошенкова, 2015). Клявлинский : окр. с. Новые Сосны (Ильина, 2004); окр. дер. Воскресенка, склоны к р. Байтермишка (Раков и др., 2008; Сенатор, Саксонов, 2013); окр. с. Русское Добрино, междуречье рр. Лесной Шешмы и Хуторской (Шаронова, Курочкин, 2015); окр. с. Иваново-Подбельское, на правом берегу р. Шешмы (Шаронова, Курочкин, 2015); окр. с. Кляв-лино, левый берег р. Средней и правый берег р. Бол. Черемшан (Шаронова, Курочкин, 2015); окр. с. Софьино (Шаронова, Курочкин, 2015). Сергиевский : ПП «Гора Высокая» (Ильина, 2006 б, 2010 б, 2014, 2015 а; Ильина, Ильина, 2009; Реестр …, 2010; Ильина, Митрошенкова, 2015); Чекалинские высоты, в левобережной части р. Елхи, в 4 км западнее с. Чекалино (Митрошенкова и др., 2015); ПП «Серноводский шихан» (Ильина, 2006 б, 2007 в, 2010 б, 2014, 2015 а; Саксонов, Васюков и др., 2013; Сенатор, Саксонов, 2013); Коренной берег р. Шунгут в окр. с. Ст. Якушкино (Шаронова, Курочкин, 2014); Успенская шишка (Ильина, 2006 б, 2007 в, 2010 б, 2014, 2015 а; Ильина и др., 2008), окр. с. Гундоровка (Ильина, 2006б, 2010, 2014, 2015 а), правый берег р. Большой Кандабулак, в окр. с. Кандабулак; правый берег р. Липовка, в окр. с. Седовка; правый берег р. Елховка, в окр. с. Новая Елховка. Похвистневский : ПП «Гора Копейка» (Ильина, 2003, 2010б, 2014, 2015а; Ильина и др., 2005; Красная книга…, 2007; Кудашкина, Плаксина, 2009; Реестр …, 2010; Ильина, Митрошенкова, 2015), ПП «Лесостепь в верховьях р. Аманак» (Ильина, 2010б, 2014, 2015а; Реестр …, 2010). Челновершинский : окр. с. Челно-Вершины, Кондурчинские Яры ( Ильина, 2009, 2014, 2015а); гора Маяк (Митрошенкова, 2014). Шенталинский : Нов. Кувак
(Иванова, Елкина, 2008; Саксонов, Сенатор и др., 2008); окр. с. Крепость Кондурча, Кондурчинские яры (Ильина, 2009).
На территории Высокого Заволжья СО отмечено несколько местообитаний, где, по нашему мнению, целесообразным является проведение ценопопуляционных исследований вида. В ходе работ нами использовались ставшие традиционными популяционно-онтогенетические методы сбора и обработки данных, основанные на дискретном описании онтогенеза модельных представителей и определении виталитета особей и их природных популяций (Работнов, 1950; Уранов, 1975; Ермакова, 1976; Ценопопуляции растений, 1976, 1977, 1988; Воронцова, Заугольнова, 1979; Жукова, 1995; Злобин и др., 2013).
Исследования ценопопуляций O. spicata проводились нами в 2000-2013 гг. на территории Самарского Высокого Заволжья (Похвистневский район – памятники природы «Гора Копейка», «Лесостепь в верховьях р. Аманак», Сергиевский район – памятники природы «Серноводский шихан», «Гора Высокая», Успенская шишка, окр. с. Гундоровка, Исаклинский район – Гора Пионерка, Челновершинский район – Кон-дурчинские Яры). Некоторые результаты работы были предварительно изложены в публикациях (Ильина, 2006а, б, 2007а, б, в, 2010б, 2014, 2015а, 2015б).
В большинстве местообитаний численность особей вида имеет флуктуационную динамику, однако в целом характерна тенденция к её снижению. В обследованных местообитаниях отмечаются далеко отстоящие друг от друга небольшие скопления особей по 5-10 особей. Нередко в популяциях таких агрегаций немного и общее количество генеративных растений в сумме составляет менее 100 экземпляров. Уменьшается в Самарской области и число местообитаний O. spicata . Например, в Чубовской луговой степи (Кинельский район СО) в последние годы вид не зафиксирован. В Предволжье на Губинско-Троекуровских высотах (Шигонский район) указан по результатам исследований 2005-2009 гг. (Ильина, Ильина, 2010), но в последующие годы остролодочник колосистый не отмечался.
В ходе полевых исследований нами установлено, что численность особей O. spi-cata в местообитаниях с незначительной антропогенной нагрузкой в целом остается неизменной, но при длительном воздействии или увеличении антропогенного пресса она снижается, в некоторых местообитаниях фиксируются только единичные экземпляры с низким уровнем виталитета; лимитируют развитие популяций перевыпас, степные палы, распашка территории; многие популяции вида в Самарской области – стареющие неполночленные, с высоким уровнем генеративных особей и средним уровнем жизненности, возобновление особей в популяциях недостаточное для ее нормального развития (Ильина, 2014, 2015а, б).
Оптимальные местообитания остролодочника характеризуются разреженным травостоем с проективным покрытием почвы 10-35%. В этом случае виды могут стабильно удерживать территорию, проявляя высокую конкурентную способность и ви-олентные черты. В угнетенных ценопопуляциях (с низкой численностью и плотностью особей, чаще при значительной антропогенной нагрузке) отмечен патиентный тип жизненной стратегии (Ильина, 2006а).
По нашему мнению, следует уделить особое внимание вопросу нерегулируемых степных палов на растительный покров. Этот вопрос неоднократно поднимается исследователями почвенно-растительного покрова в степной зоне (Родин, 1981; Малышева, Малаховский, 2000; Рябцов, Сафонов, 2002; Опарин, Опарина, 2003; Попов, 2004; Калмыкова, 2006; Рябцов, 2006; Тишков, 2009; Ильина, 2010а, 2011; Дубинин и др., 2012; Колчин и др., 2012; Смелянский, 2012; Павлейчик, 2015, 2016а, б; Павлей- чик и др., 2016). Неоднократно нами фиксировалось негативное влияние степных палов на структуру и состояние популяций вида в Самарском Заволжье.
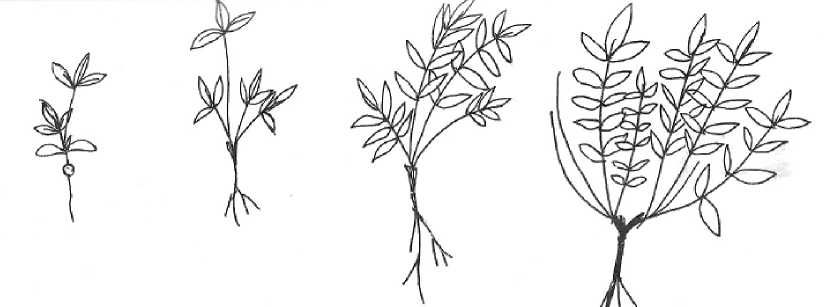
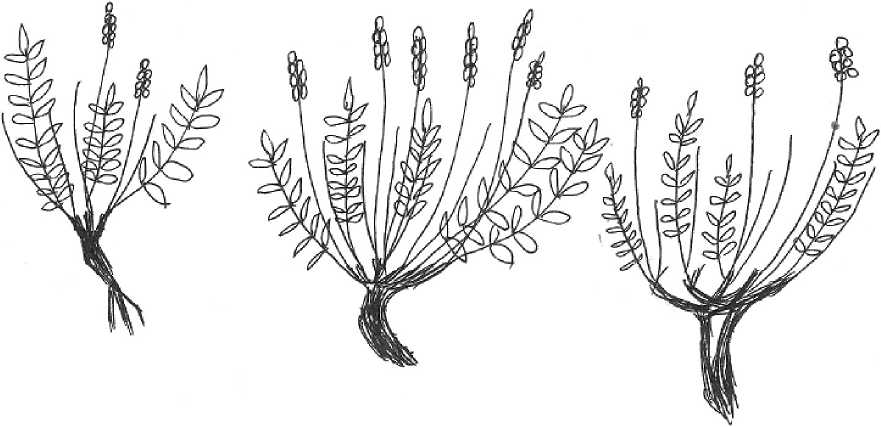
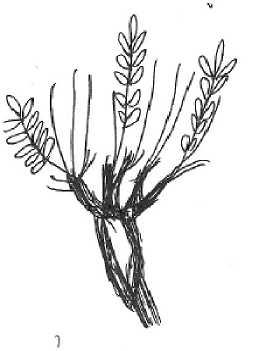
Ранее онтогенез вида описан не был. Изучение особей O. spicata на стационарных участках позволили описать и предварительно оценить длительность основных стадий онтогенеза. Онтогенетические стадии представлены на рисунке.
Латентный период . Возобновление O. spicata осуществляется только семенным путем. Семена созревают в конце июля – в августе, плоды опадают недалеко от материнского растения, что обусловливает агрегацию особей в популяциях. Бобы яйцевидные, 13-15 мм длины, 6-7 мм ширины, бархатисто оттопыренно опушенные, тесно прижаты друг к другу, с узкой (около 1 мм ширины) перегородкой по брюшному шву. Семена в течение достаточно продолжительного срока могут сохраняться в почве и прорастать через несколько лет, поскольку для них, как и для многих других видов сем. Fabaceae характерна твердосемянность. В почве может накапливаться значительный банк семян. Лимитируют сохранность банка семян степные палы. Cвежесо-бранные семена твердосемянностью не обладают (лабораторная всхожесть около 90 %). При хранении увеличивается доля твердых семян: твердосемянность через 6 месяцев хранения составляет 35 %, через 12 месяцев хранения – около 67%, через 18 месяцев хранения – 84%.
Прегенеративный период . Проростки (p) в природных условиях появляются после схода снега и прогревания почвы. Основная масса проростков O. spicata появляется в середине мая-начале июня в зависимости от погодных условий и условий местообитания. Семядоли овальные и зеленые, длина 4-9 мм, ширина 2-4 мм. Первый лист тройчатосложный, листочки продолговато-яйцевидные, цельнокрайние, сверху опушенные. Пластинки длиной 0,5-0,7 см, шириной 0,4-0,7 см, длина черешка – 0,92,4 см. После появления 2-3 листа семядоли отмирают. Характерна катастрофическая гибель проростков. Длительность состояния 1-2 недели.
Ювенильное состояние (j). Растения однопобеговые, 3,8-7,0 см высотой, несущие 3-5(7) листьев ювенильного типа – тройчатые, в их пазухах закладываются боковые почки. Начинают появляться единичные непарноперистосложные листья с 2-3 парами листочков. Уязвимы и не обладают устойчивостью в сообществах. Длительность состояния 2-4 недели.
Имматурное состояние (im). Растение представлено первичным побегом, достигает высоты 5,5-12,0 см, имеет 5-7(11) непарноперистосложных листьев с 3-6(8) парами листочков. Первые тройчатосложные листья отмирают, на особях сохраняются до 6-10(12) ювенильных листьев. Начинается формирование каудекса (пока одноглавый). При окончательном формировании данного онтогенетического состояния пазушные почки на базальной части главной оси побега, находящиеся в почве, прорастают и образуют 2-4 розеточных побега. Длительность состояния – 1-3 месяца.
Виргинильное состояние (v). В связи с появлением новых побегов у особей O. spicata начинается процесс кущения. Виргинильные растения имеют 1-4(6) побегов 8,5-18,2 см высотой с 6-18(24) листьями взрослого типа. Листья 10-20 см дл., с 14-20 парами листочков, черешки, как и оси, довольно густо опушены оттопыренными и полуоттопыренными, относительно короткими (меньше поперечника черешка) волосками; листочки продолговато-яйцевидные, сверху опушенные полуоотпыренными, рыхло прилегающими, тусклыми волосками, снизу довольно густо рыхло прилегающими и полуоттопыренными волосками. Продолжается формирование каудекса, пока расположенного под землей компактно (базальные части побегов с расположенными на них почками возобновления в числе 3-12(15) штук). Растение приобретает вид взрослой нецветущей особи, имеющий 2-3(5) розеточных побега. Длительность состояния - 1-12 месяцев, иногда до трех лет.
Генеративный период. Молодые генеративные растения (g1) характеризуются началом формирования многоглавого (2-4 главки) каудекса в результате развития боковых розеточных побегов; имеют 2-8(11) побегов (0-4(6) вегетативных и 1-3(5) генеративных побегов). Высота розетки листьев 8,4-22,8 см, диаметр розетки 11,6-23,5 см. Из пазух некоторых листьев розетки отходят 1-3(8) цветоносов (стрелки), они длиннее листьев, бороздчатые, б.м. густо (часто бархатисто) опушенные оттопыренными волосками. Кисти 5-10 см дл., весьма густые, 25-40-цветковые, при плодах не рыхлеющие. Распускание цветков приходится на начало июня, массовое цветение -на середину июня (что зависит от метеоусловий, экспозиции склона, вытаптывания при неумеренном выпасе крупного рогатого скота). В июле на растениях можно наблюдать соцветия с бутонами, цветками, с зелеными и зрелыми плодами, что свидетельствует о проявлении ритмологической поливариантности, что указывается и для других видов остролодочников (Маслова и др., 2011). Прицветники яйцевидноланцетные, 5-8 мм длины; чашечка трубчато-колокольчатая, 9-12 мм длины, более или менее густо опушена полуоттопыренными и оттопыренными белыми волосками, иногда с примесью чёрных, зубцы её в 4-5 раз короче трубки; венчик светлопурпурный, розовый, голубовато-розовый, в гербарии голубой, 15-16 мм длины, 7-10 мм ширины, с пластиной обратнояйцевидной на верхушке слегка выемчатой. Крылья 12-13 мм дл., 4-6 мм ширины. Лодочка чуть короче крыльев, с носиком 1 мм длиной. Длительность состояния - 1-3(5) лет.
Средневозрастные генеративные растения (g2) характеризуются наибольшей мощностью и составляют ядро популяции в условиях существования, близких к оптимальным. Высота с соцветием до 28-45(60) см. У растений этого возрастного состояния число побегов (4-10(12) шт. из них 0-3(5) вегетативных и 4-10(14) генеративных побегов), листьев (16-78), соцветий (4-7(10)) и цветков (13-30(56)) в соцветии, размеры листьев (6-18(27) см длиной и 1,5-4,3(6,6) см шириной) и число пар листовых пластинок (5-21(28)) достигают максимума, каудекс становится многоглавым (3-4(8) главок). Длительность состояния - 1-6 и более лет.
Старые генеративные растения (g3) характеризуются уменьшением размеров растений (высота особей достигает 25-35(51) см, число побегов 5-12(16), из них вегетативных 0-4(6), генеративных 1-6(12)), биомассы, сокращением элементов в репродуктивной сфере (кроме числа генеративных побегов уменьшается число соцветий (14(8) и число цветков (6-19(25)). Особи отличаются крупными размерами каудекса с явными признаками партикуляции и распада на главки. С возрастом усиливается процесс отмирания тканей (отмерших тканей более 50 %). Длительность состояния - 1-2 и более лет.
Постгенеративный период. Субсенильное состояние (ss). Растение имеет 37(9) розеточных вегетативных побегов, высота до 10-15 см, расположенных они в основном по периферии каудекса. Число побегов и листьев уменьшается по мере отмирания отдельных глав каудекса. Происходит сильная общая партикуляция. При выраженном антропогенном прессе доля субсенильных растений в популяциях достигает 30%. Длительность состояния - 3-6 месяцев, иногда до 3 лет.
Сенильное состояние (s). Растения со слабыми побегами, несущими 2-5 листа с несколькими парами листочков. Иногда остается одиночный розеточный вегетативный побег с 3-4 некрупными листьями. Отмершие главки каудекса сохраняются частично, на живых главках регистрируется много мертвых участков, и они сильно раз- 105
рушены. Подобные особи регистрируются редко. Длительность состояния – 1-6 меся- цев.
Проросток Ювенильное Имматурное Виргинильное
состояние состояние состояние
Молодое генеративное со-
Зрелое генеративное со-
Старое генеративное со- стояние
стояние
стояние

Субсенильное состояние
Сенильное состояние
Рис. Онтогенетические стадии Oxytropis spicata (Pall.) O. et B. Fedtsch. ( Fabaceae ) (рисунок автора)
Таблица. Соотношение особей разных онтогенетических групп
|
№ п/п |
Местообитание, год исследования и число ценопо-пуляций |
Демографические характеристики ценопопуляций (%) |
||||||
|
p-v |
g1-g3 |
ss-s |
Индекс замещения |
Индекс восстановления |
Индекс старения |
Стабильность |
||
|
1 |
Гора Копейка – 2002 (6) |
17,1 |
82,9 |
0 |
0,21 |
0,21 |
0 |
82,9 |
|
2 |
Гора Копейка – 2005 (8) |
11,2 |
74,0 |
14,8 |
0,13 |
0,15 |
0,17 |
88,8 |
|
3 |
Гора Копейка – 2007 (7) |
10,8 |
74,1 |
15,1 |
0,12 |
0,15 |
0,18 |
89,2 |
|
4 |
Гора Копейка – 2009 (4) |
11,8 |
76,2 |
12 |
0,13 |
0,16 |
0,14 |
88,2 |
|
5 |
Гора Копейка – 2010 (9) |
8,4 |
86,3 |
5,3 |
0,09 |
0,10 |
0,06 |
91,6 |
|
6 |
Гора Копейка – 2011 (7) |
15,4 |
79,5 |
5,1 |
0,18 |
0,19 |
0,05 |
84,6 |
|
7 |
Гора Копейка – 2012 (5) |
9,6 |
85,9 |
4,5 |
0,10 |
0,11 |
0,05 |
90,4 |
|
8 |
Гора Копейка – 2013 (11) |
6,9 |
83,6 |
9,5 |
0,07 |
0,08 |
0,11 |
93,1 |
|
9 |
Серноводский шихан – 2004 (4) |
7,6 |
87,3 |
5,1 |
0,08 |
0,09 |
0,05 |
92,4 |
|
10 |
Серноводский шихан – 2005 (4) |
7,6 |
87,0 |
5,4 |
0,08 |
0,09 |
0,06 |
92,4 |
|
11 |
Серноводский шихан – 2007 (7) |
8,2 |
90,2 |
1,6 |
0,09 |
0,09 |
0,02 |
91,8 |
|
12 |
Серноводский шихан – 2009 (6) |
5,6 |
89,0 |
5,4 |
0,06 |
0,06 |
0,06 |
94,4 |
|
13 |
Серноводский шихан – 2010 (10) |
4,5 |
90,8 |
4,7 |
0,05 |
0,05 |
0,05 |
95,5 |
|
14 |
Серноводский шихан – 2012 (10) |
5,6 |
87,0 |
7,4 |
0,06 |
0,06 |
0,08 |
94,4 |
|
15 |
Гора Высокая – 2004 (3) |
15,6 |
79,2 |
5,2 |
0,19 |
0,19 |
0,05 |
84,4 |
|
16 |
Гора Высокая – 2005 (4) |
16,6 |
78,1 |
5,3 |
0,20 |
0,21 |
0,06 |
83,4 |
|
17 |
Гора Высокая – 2007 (6) |
10,3 |
84,4 |
5,3 |
0,12 |
0,12 |
0,06 |
89,7 |
|
18 |
Гора Высокая – 2009 (10) |
17,6 |
80,1 |
2,3 |
0,21 |
0,22 |
0,02 |
82,4 |
|
19 |
Гора Высокая – 2010 (10) |
30,3 |
68,2 |
1,5 |
0,43 |
0,44 |
0,02 |
69,7 |
|
20 |
Гора Высокая – 2012 (10) |
10,6 |
86,0 |
3,4 |
0,12 |
0,12 |
0,04 |
89,4 |
|
21 |
Успенская шишка – 2004 (3) |
17,1 |
80,5 |
2,4 |
0,21 |
0,21 |
0,03 |
82,9 |
|
22 |
Успенская шишка – 2005 (3) |
18,3 |
81,7 |
0 |
0,22 |
0,22 |
0 |
81,7 |
|
23 |
Успенская шишка – 2007 (5) |
7,6 |
87,0 |
5,4 |
0,08 |
0,09 |
0,06 |
92,4 |
|
24 |
Успенская шишка – 2009 (5) |
7,6 |
87,0 |
5,4 |
0,08 |
0,08 |
0,05 |
92,4 |
Продолжение таблицы
|
№ п/п |
Местообитание, год исследования и число ценопо-пуляций |
Демографические характеристики ценопопуляций (%) |
||||||
|
p-v |
g1-g3 |
ss-s |
Индекс замещения |
Индекс восстановления |
Индекс старения |
Стабильность |
||
|
25 |
Успенская шишка – 2010 (5) |
6,8 |
88,8 |
4,4 |
0,07 |
0,07 |
0,04 |
93,2 |
|
26 |
Успенская шишка – 2012 (5) |
7,0 |
88,7 |
4,3 |
0,08 |
0,08 |
0,04 |
93,0 |
|
27 |
Гундоровка – 2005 (2) |
17,2 |
69,0 |
13,8 |
0,21 |
0,25 |
0,16 |
82,8 |
|
28 |
Гундоровка – 2010 (5) |
10,3 |
85,6 |
4,1 |
0,11 |
0,12 |
0,04 |
89,7 |
|
29 |
Гора Пионерка – 2005 (3) |
20,7 |
75,6 |
3,7 |
0,26 |
0,27 |
0,03 |
79,3 |
|
30 |
Гора Пионерка – 2010 (5) |
20,9 |
72,9 |
6,2 |
0,26 |
0,29 |
0,07 |
79,1 |
|
31 |
Гора Пионерка – 2012 (6) |
20,3 |
72,9 |
6,8 |
0,26 |
0,28 |
0,07 |
79,7 |
|
32 |
Лесостепь в верховьях р. Аманак – 2010 (6) |
28,0 |
70,0 |
2,0 |
0,39 |
0,40 |
0,02 |
72,0 |
|
33 |
Лесостепь в верховьях р. Аманак – 2012 (7) |
17,0 |
78,7 |
4,3 |
0,21 |
0,22 |
0,05 |
83,0 |
|
34 |
Лесостепь в верховьях р. Аманак – 2013 (12) |
17,9 |
74,9 |
7,2 |
0,22 |
0,24 |
0,08 |
82,1 |
|
35 |
Кондурчинские Яры (окр. с. Челно-Вершины) – 2008 (9) |
20,5 |
75,9 |
3,6 |
0,26 |
0,27 |
0,04 |
79,5 |
|
36 |
Кондурчинские Яры (окр. с. Челно-Вершины) – 2011 (7) |
20,9 |
75,9 |
3,2 |
0,26 |
0,28 |
0,03 |
79,1 |
|
Среднее значение |
13,6 |
81,0 |
5,4 |
0,15 |
0,16 |
0,06 |
86,4 |
|
В таблице приводятся основные параметры, характеризующие динамику популяций O. spicata (доля прегенеративных (p-v), генеративных (g1-g3) и постгенеративных (ss-s) особей, индексы восстановления популяций, замещения особей, старения и показатель стабильности популяций).
Установлено, что генеративная фракция особей преобладает в популяциях (81%), прегенеративные особи составляют около 13,6% от численности, постгенеративные – 5,4%. Лишь в двух случаях (гора Высокая, 2010 г; Лесостепь в верховьях реки Аманак, 2010) доля виргинильных растений достигала почти 30%. Высокий процент сенильных растений отмечен трижды (гора Копейка, 2005, 2007; окр. с. Гундоровка, 2005) – около 14-15%. Максимальный индекс замещения отмечен на территории памятника природы «Лесостепь в верховьях р. Аманак» (2010) – 0,389%, минимальный - на территории памятника природы «Серноводский шихан» (2010) – 0,047%. Индекс восстановления имеет максимальное и минимальное значение в тех же пунктах – 0,40% и 0,05% соответственно. Минимальным оказалось старение популяций на горе Копейке (2002) и Успенской шишке (2005) – 0%. Максимально популяции «состарились» на горе Копейке (2005, 2007), в окр. с. Гундоровка (2005) – 16-18% особей. Однако в большинстве случаев популяциям свойственна стабильность численности и возрастного спектра – в среднем 86,4%
Определение виталитетного состояния особей проводили на основе анализа высоты экземпляров, количества побегов, числа цветоносов, диаметра каудек-са, длины листьев, размеров листочков, количества цветков в соцветии и некоторых других признаков. После проведения статистической обработки полученных данных было выделено три класса виталитета. Установлено, что в ценопопуля-циях преобладают особи второго (среднего) уровня жизненности.
Таким образом, популяции остролодочника колосистого в СО обычно характеризуются низкой численностью, групповым размещением особей, средним уровнем виталитета, флуктуационной динамикой онтогенетической структуры. Вид, проявляя черты патиента, выпадает из сообществ на территориях, подверженных значительному антропогенному прессу. Длительность онтогенеза оценивается в 5-20 и возможно более лет.
Список литературы Онтогенез и динамика популяций остролодочника колосистого (Oxytropiss picata (Pall.) O. et B. Fedtsch., Fabaceae) в Самарской области
- Абрамова Л.М., Баширова Р.М., Муртазина Ф.К., Усманов И.Ю. Характеристика ценопопуляций Glycyrrhiza korchinskyi Grig. на юго-востоке Республики Башкортостан//Растительные ресурсы. 2001. Т. 37. № 2. С. 24-29.
- Абрамова Л.М., Каримова О.А., Андреева И.З. Структура и состояние популяций Althaea officinalis (Malvaceae) на юге Предуралья (Республика Башкортостан)//Растительные ресурсы. 2010. Т. 46. № 4. С. 47-54.
- Васюков В.М., Саксонов С.В., Сенатор С.А. Эндемичные растения бассейна Волги//Фиторазнообразие Восточной Европы. 2015. Т. IX. Вып. 3. С. 27-43.
- Воронцова Л.И., Заугольнова Л.Б. О подходах к изучению ценопопуляций растений//Бот. журнал. 1979. Т. 61. № 9. С. 1296-1306.
- Головлёв А.А. О популяциях Scabiosa isetensis L. в Сокольих горах//Теоретические проблемы экологии и эволюции: Шестые Любищевские чтения, 11-й Всероссийский популяционный семинар и Всероссийский семинар «Гомеостатические механизмы биологических систем» с общей темой «Проблемы популяционной экологии»/Под ред. Г.С. Розенберга. 2015. С. 94-96.
