Онтогенез и структура популяции Fritillaria camschatcensis (Liliaceae) в северной части ареала вида (Магаданская область)
 в северной части ареала вида (Магаданская область)")
Автор: Кондакова Д.А.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 3 т.19, 2025 года.
Бесплатный доступ
Проведено исследование онтогенеза рябчика камчатского и структуры его популяции в северной части ареала на примере популяции разнотравного луга приморского склона, где вид доминирует в мае-июне. Приведены сведения об онтогенезе рябчика камчатского. Выявлены онтогенетические состояния растений и приведено морфологическое описание семени, проростка, ювенильного, имматурного, виргинильного, молодого генеративного, зрелого генеративного, временно нецветущего растений. Даны ключи и диагнозы онтогенетических состояний. Приведены сведения о вегетативном размножении рябчика камчатского. Определен тип онтогенеза. Проанализированы гендерная и онтогенетическая структуры популяции. Проведено сравнение с популяциями из южной части ареала на примере японских популяций по литературным данным.
Fritillaria camschatcensis, онтогенетическое состояние, проросток, имматурное растение, онтогенез, структура популяции
Короткий адрес: https://sciup.org/148331547
IDR: 148331547 | УДК: 582.572.2(571.65):581.15 | DOI: 10.24412/2072-8816-2025-19-3-175-188
Ontogenesis and population structure of Fritillaria camschatcensis (Liliaceae) in the northern part of the species range (Magadan region)
The population of Fritillaria camschatcensis, which dominates in spring on the seaside slope within the forb meadow, is represented by sprouts, juvenile plants, immature, virginal, young and mature generative, temporarily non-flowering plants. Plants in the immature, virginal, young and mature generative states are characterized by the formation of up to two stolons with particles rejuvenated to the state of an immature plant. According to Zhukova's classification, the life cycle is classified as type D. The gender structure of the population is close to the ratio of 1:1. The ontogenetic spectrum of the population is normal, a large group of immature plants imparts to the spectrum a pointed peak and positive asymmetry, characteristic of the species and explained by the features of the life cycle. Compared with the southern boundary of the range, considered using the example of Japanese populations, the northern Okhotsk population is represented by a larger proportion of mature generative plants, a group of seedlings, the population density is several times lower than the density of the Japanese population, which is explained by a significant slope (on average 31°) and the scattering of seeds along the slope.
Текст научной статьи Онтогенез и структура популяции Fritillaria camschatcensis (Liliaceae) в северной части ареала вида (Магаданская область)
Fritillaria camschatcensis (L.) Ker Gawl. (рябчик камчатский) – столонообразующий луковичный многолетник, вегетативно подвижное растение. Это декоративное (Otani, Shimada, 1997) и лекарственное растение (Iguchi et al, 2022). Ареал вида – Япония, Корея, северо-запад Северной Америки, Алеутские острова, в России – Охотское побережье в пределах Магаданской области и Корякии, Камчатка, Алеутские острова, Сахалин, Курилы, Амурская область, Приморье, юг Хабаровского края. Растение встречается на разнотравных и приморских лугах, опушках каменноберезовых лесов и зарослей ольхового стланика, на закустаренных луговинах в лиственничниках (Barkalov, 1985; GBIF, 2025; POWO, 2025).
Целью исследования было изучить онтогенез и возрастную структуру популяции рябчика камчатского. Были поставлены следующие задачи:
-
1. Собрать коробочки с семенами в период плодоношения до раскрытия коробочек, собрать растения рябчика камчатского весной и в период цветения.
-
2. Исследовать полученные образцы, выявить качественные признаки, присущие растениям, установить количественное выражение этих признаков (длину, ширину, вес, цвет, количество).
-
3. Дифференцировать растения по наличию у них разных качественных признаков и ранжировать их в порядке появления и исчезновения у них качественных признаков.
-
4. На основании полученных признаков онтогенетических состояний произвести учет особей разных возрастных состояний на учетных площадках.
-
5. Учесть процент особей в популяции с обоеполым цветком.
Популяции рябчика камчатского, как и его онтогенез, ранее изучались в Японии (Shimizu et al., 1998, Nogami et al., 2008), но методика японских исследователей значительно отличается от принятой в российской популяционно-онтогенетической школе.
М АТЕРИАЛЫ И МЕТОДЫ
Сбор растений разных онтогенетических состояний, закладка пробных площадей и учет возрастных состояний проводились с апреля по июль 2025 г. на приморском склоне (уклоном 32°) вблизи пос. Нюкля, в 4 км к юго–западу от поселка Ола, Ольского района Магаданской области (N 59°34′21.85″, E 151°13′07.42″, h=16 m) (Рис 1). Пробные площади заложены на склоне в пределах мозаичного фитоценоза. В пределах пробных площадей (рис. 1) были выделены участки разнотравного луга. Осыпи, обнажения, заросли кустарников, отдельно стоящие деревья, ложбины, вытаптываемые участки были исключены из исследования. Сбор растений проводили на отдельных участках вне учетных площадок. Выполнено описание фитоценоза.
Для изучения диагностических признаков и внешних отличий онтогенетических состояний на склонах близ пос. Нюкля растения выкапывали вместе с фрагментами грунта 10 х 10 см глубиной 5 – 7 см. Всего собрано 123 растения рябчика камчатского в разных онтогенетических состояниях.
В качестве определения онтогенетических состояний использовалась классификация Т.А. Работнова и критерии выделения возрастных состояний, предложенные А.А. Урановым (Smirnova et al., 1976). Для поиска растений предгенеративного периода развития учитывались морфологические признаки, отмеченные М. В. Барановой у разновозрастных сеянцев рябчика камчатского (Baranova, 1999). Тип жизненного цикла определяли по Жуковой (1988).
Сбор коробочек с семенами проводился 13.08.2024 г. на участке склонового разнотравного луга на побережье в 4 км к западу от поселка Ола (N 59°34′19.57″, E 151°13′23.37″, h=24 m). Всего было собрано 32 побега с 1–3 коробочками. Проростки и ювенильные растения собирались 18.06.2025 г. на участке разнотравного приморского луга, на приморском склоне южной экспозиции с уклоном 18ᴼ, в 5 км к западу от поселка Ола (близ пос. Нюкля), (N 59°34′23.80″, E 151°12′58.00″, h=3 m). Позднее (28.06.2025 г.), там же проведено исследование линейных размеров листа имматурных растений.
Морфометрические показатели растений измерялись в камеральных условиях. Для 30 семян, случайным образом отобранных из 30 конвертов с семенами (по одному от каждого растения), определяли: высоту и ширину семени с семенной оболочкой (мм), длину и ширину эндосперма с зародышем (мм), толщину семени (мм). Для проростков с семенем на апикальном конце семядоли измерялись длина и толщина семядоли (см), а для проростков с семядольным листом, утративших семенную оболочку – длина, ширина и толщина семядольного листа в верхней уплощенной и нижней частях листа (см), диаметр и высота семядоли (см). У ювенильных растений измеряли высоту растения (см), длину и ширину листовой пластинки (см), диаметр и высоту листового влагалища зеленого листа (см), число жилок на листе, число листовых влагалищ, число луковичных чешуек, число листьев, для имматурных – длина и ширина листовой пластинки (см), число жилок на листе. У виргинильных и нецветущих растений измеряли высоту побега (см), число зеленых листьев, число мутовок, длину и ширину нижнего листа нижней мутовки (см), дину и ширину верхнего самого маленького листа верхней мутовки (см), высоту и диаметр луковицы (см), число запасающих чешуй, образовавших верхнюю поверхность луковицы и сильно сросшихся с донцем. Также отмечали наличие побега предыдущего года и его онтогенетическое состояние по форме луковицы и наличию в ее составе остатков стебля. Морфометрические показатели имматурных и зрелых генеративных растений, включая размеры луковиц имматурных растений, опубликованы ранее (Kondakova, Sinelnikova, 2025).
Полученные результаты обрабатывали с помощью программы MS Office Exсel с надстройкой «Анализ данных». Вычисляли минимальное ( ^min,) максимальное ( ^max ) и среднее арифметическое (X) значение переменной в выборке, моду (Ivanter, Korosov, 2010) Для выявления изменчивости морфологических признаков использовали коэффициент вариации (CV, %), его оценивали по шкале Мамаева, где приняты следующие уровни изменчивости: очень низкий (меньше 7 %), низкий (8–12 %), средний (13–20 %), повышенный (21–30 %), высокий (31–40 %), очень высокий (больше 40 %) (Mamaev, 1972). Критерий надежности t=2, порог вероятности безошибочных прогнозов β=0.95. Для характеристики онтогенетического спектра вычисляли эксцесс и коэффициент асимметрии распределения (Plokhinskiy, 1970).
При изучении структуры популяции использован эколого-демографический подход (Kashin et al., 2015). Для изучения онтогенетической структуры популяции использована рамка со стороной 1 м2, размеченная шнурами на ячейки по 10 см2. Изучено 33 учетные площадки, онтогенетическое состояние определено у 9099 растений популяции разнотравного луга на приморском склоне близ пос. Нюкля в период с 18.05.25 г. по 28.06.25 г.
Гендерная структура популяции выявлена с помощью оценки 200 растений на предмет наличия хотя бы одного обоеполого цветка. Оценку проводили в пределах пробной площади, поднимаясь по склону на 15 метров вверх, оценивая по пять растений в пределах метра слева от себя, затем справа от себя, продвигаясь вверх с шагом 2–3 метра, всегда пропуская растения, выпавшие из поля зрения. Подъем повторяли со смещением с востока на запад на 5 метров.
Р ЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
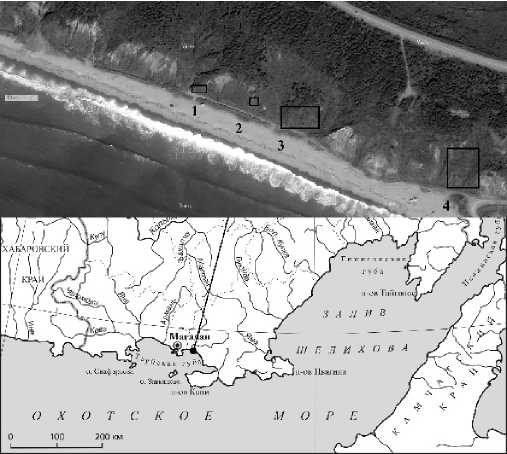

Рис. 1. Расположение пробных площадей. 1 – место сбора проростков, исследования имматурных растений, место размещения учетных площадок, 2, 3 – места размещения учетных площадок, 4 – место сбора коробочек с семенами.
Fig. 1. Location of the test areas. 1 – the place of collection of seedlings, research of immature plants, the location of accounting sites, 2, 3 – the locations of accounting sites, 4 – the place of collection of seed boxes.
Рис. 2. Разнотравный луг на приморском склоне южной экспозиции близ пос. Нюкля (пробная площадь № 3).
Fig. 2. A grass meadow on the seaside slope of the southern exposure near the Nyklya settlement (trial area No. 3)
В мае 2025 года на приморских склонах доминировали Potentilla fragiformis Willd. ex D.F.K. Schltdl., Fritillaria camschatcensis (L.) Ker Gawl., Ligusticum scothicum L., субдоминанты Corydalis magadanica A.P. Khokhr., Myosotis asiatica (Vestergr.) Schischk. et Serg. Остальные растения находились в фазах отрастания и развертывания листьев. В первой декаде июня доминировали Potentilla fragiformis Willd. ex D.F.K. Schltdl., Fritillaria camschatcensis (L.) Ker Gawl., Ligusticum scothicum L, Artemisia arctica Less., субдоминанты Parasenecio hastatus (L.) H. Koyama, Angelica gmelinii (DC.) Pimenov, Myosotis asiatica (Vestergr.) Schischk, Rubus arcticus L., редко, Aruncus dioicus (Walter) Fernald, Lysimachia europaea (L.) U. Manns et Anderb.
Онтогенез
Состояние покоя. Семена
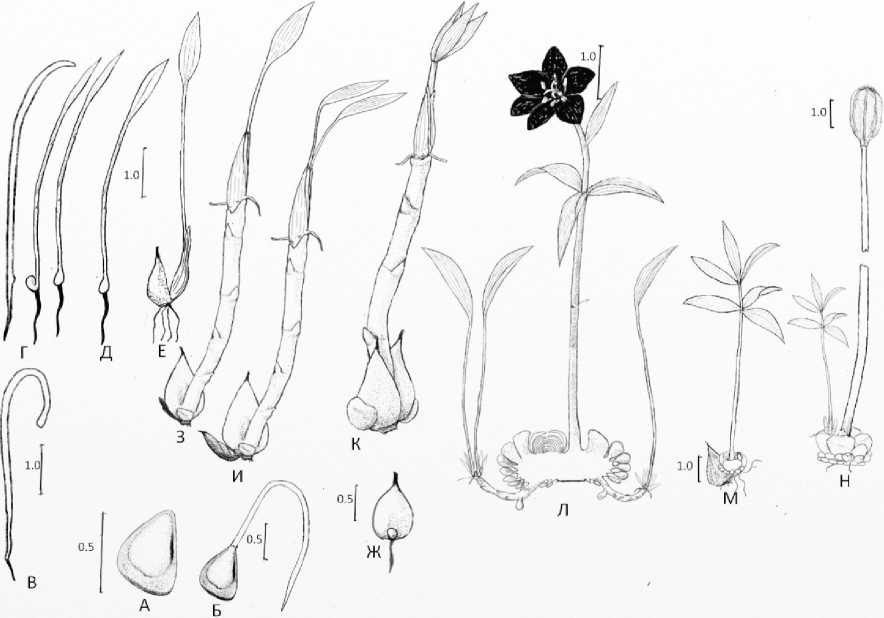
Рис. 3. Онтогенетические состояния Fritillaria camschatcensis : А – семя, Б – проросток с семенем в апреле-мае, В – проросток с семядолей в мае-июне, Г – проросток с семядолей и образующимся семядольным влагалищем в июне (три растения), Д – ювенильные растение с линейно-ланцетным листом, E – ювенильное растение с тремя жилками на листе, Ж – ювенильное растение в состоянии покоя, З – имматурное растение на вершине столона, растущего из почки в пазухе листового влагалища ювенильного растения, И – имматурное растение с двумя листьями на столоне, К – имматурное растение с двумя листьями, давшее столон с виргинильным растением, Л – молодое генеративное растение, образовавшее два клона на столонах, М – виргинильное растение, Н – временно нецветущее растение.
Fig. 3. Ontogenetic states of Fritillaria camschatcensis : А – seed, Б – seedling with seed in April-May, В – seedling with cotyledons in May-June, Г – seedling with cotyledons and the resulting cotyledon sheath in June (three plants), Д – juvenile plant with linear-lanceolate leaf, Е – juvenile plant with with three veins on a leaf, Ж – is a juvenile plant at rest, З – is an immature plant on top of a stolon growing from a bud in the axil of the leaf sheath of a juvenile plant, И – is an immature plant with two leaves on a stolon, К – an immature plant with two leaves that produced a stolon with a virginal plant, Л – is a young generative plant that formed two clones on stolons, М – is a virginal plant, Н – is a temporarily non-flowering plant.
Рябчик камчатский образует коробочки с тремя камерами, в каждой камере расположены две стопки семян, отгороженные друг от друга перегородкой коробочки. Морфологические признаки семян приведены в таблице 1. Осенью коробочки с семенами раскрываются, и семена попадают на подстилку. По нашим наблюдениям, часть семян не высыпается из коробочек и остается внутри коробочки до следующего лета. Семена каплевидные, семенная кожура образует крыло (рис. 1). Семенная кожура охристого цвета, глянцевая, гладкая (рис. 3, А). При попадании в подстилку острый конец семени с микропиле часто устремлен вниз, а крыло вверх.
Число семяпочек рябчика камчатского определяется биологическими особенностями вида. Число нормальных семян и процент семинификации показывают высокую неоднородность условий произрастания.
Таблица 1. Морфологические признаки семян Fritillaria camschatcensis (t=2)
Table 1. Morphological characteristics of seeds of Fritillaria camschatcensis (t=2)
|
Признак |
^771 ГП |
^ max |
% ± Д |
m |
CV, % |
Оценка CV |
|
Высота крыла, мм |
4.5 |
6.4 |
5.5±0.037 |
0.019 |
8.34 |
низкий |
|
Ширина крыла, мм |
3.3 |
4.7 |
3.9±0.026 |
0.013 |
9.77 |
низкий |
|
Высота семени, мм |
3.2 |
4.5 |
3.8±0.025 |
0.013 |
8.60 |
низкий |
|
Ширина семени, мм |
1.7 |
2.8 |
2.2±0.015 |
0.008 |
11.52 |
низкий |
|
Толщина семени с оболочкой, мм |
0.2 |
0.4 |
0.3±0.0002 |
0.0009 |
22.89 |
повышенный |
Состояние проростка
Весной семена поверхностно прорастают. В 2025 году на разнотравном лугу на приморском склоне близ пос. Нюкля начало прорастания семян отмечено 26 апреля. Из семени появляется слабоокрашенный в желтоватый или салатовый цвет недифференцированный проросток (рис 3, Б). Со временем проросток удлиняется и устремляется в субстрат, формируя зародышевый корень, наземная часть проростка приобретает яркую зеленую окраску и удлиняется, приподнимая семенную оболочку над поверхностью субстрата – формируется семядольный лист. Семенная оболочка при этом часто отпадает (рис. 3, В). Семядольный лист удлиняется, его верхняя пятая часть формирует линейно-ланцетную листовую пластинку без жилок. А в основании семядольного листа в виде бугорка, а затем в виде сферического утолщения формируется влагалище семядоли (рис. 3, Г). Листовая пластинка и верхняя треть побега ярко зеленые, глянцевые, средняя часть побега окрашена антоцианом, нижняя не окрашена. Морфологические признаки проростков приведены в таблице 2.
Таблица 2. Морфологические признаки проростков Fritillaria camschatcensis (t=2)
Table 2. Morphological features of Fritillaria camschatcensis seedlings (t=2)
|
Признак |
i±A |
m |
CV,% |
^mm |
Оценка CV |
|
|
Семядольный лист |
||||||
|
Длина, см |
8.85±0.571 |
0.285 |
18.33 |
5.2 |
11.9 |
средний |
|
Длина зеленой части, см |
4.86±0.316 |
0.158 |
22.77 |
2.5 |
8.2 |
повышенный |
|
Наибольшая ширина, см |
0.09±0.006 |
0.003 |
33.82 |
0.02 |
0.14 |
высокий |
|
Толщина листовой пластинки, см |
0.03±0.0021 |
0.001 |
27.46 |
0.02 |
0.05 |
повышенный |
|
Диаметр черешковой части, см |
0.03±0.002 |
0.001 |
29.08 |
0.02 |
0.05 |
повышенный |
|
Влагалище семядоли |
||||||
|
Диаметр, см |
0.17±0.011 |
0.006 |
24.49 |
0.1 |
0.32 |
повышенный |
|
Высота, см |
0.24±0.016 |
0.008 |
18.23 |
0.1 |
0.32 |
средний |
Ювенильное состояние.
На второй год жизни растение формирует лист с узколанцетной листовой пластинкой с 1 – 3 жилками на длинном черешке, основание черешка растет из пазухи перезимовавшего влагалища семядоли и в процессе вегетации формирует собственное листовое влагалище (рис. 3, Е, Ж, таблица 3). С образованием первого столона с зачатком замещающей луковицы стадия ювенильного растения завершается. Число жилок, ширина и длина листовой пластинки, высота побега и ширина луковицы у ювенильных растений слабо изменчивы.
Таблица 3. Морфологические признаки Fritillaria camschatcensis в ювенильном состоянии (t=2)
Table 3. Morphological features of Fritillaria camschatcensis in the juvenile state (t=2)
|
Признак |
х±Л |
m |
МОДА |
CV, % |
Оценка CV |
|
Число жилок |
3±0.25 |
0.12 |
3 |
17.8 |
средний |
|
Ширина листовой пластинки, мм |
3.4±0.32 |
0.16 |
2.5 |
24.21 |
повышенный |
|
Длина листовой пластинки, см |
1.6±0.15 |
0.08 |
1.6 |
24.86 |
повышенный |
|
Высота побега см |
6.5±0.60 |
0.30 |
6.9 |
20.43 |
средний |
|
Возраст луковицы, год |
2±0.17 |
0.08 |
2 |
21.71 |
повышенный |
|
Чешуи луковицы |
1±0.11 |
0.05 |
0 |
112.48 |
очень высокий |
|
Ширина луковицы, мм |
2.1±0.20 |
0.10 |
1.5 |
36.25 |
высокий |
Имматурное состояние . Образование нового органа – столона знаменует наступление стадии имматурного растения. На вершине столона образуется растение, несущее зачаток побега, в составе которого множество пленчатых чешуй, в течение вегетации образующих мясистые чешуи луковицы. Центральное положение в верхней части луковицы занимает коническое мясистое листовое влагалище зеленого листа. А в пазухе этого листового влагалища расположена почка возобновления. Зеленый лист у имматурного растения часто один, редко два. Внешним отличительным признаком этой стадии является появление более трех жилок на листе растения. В апреле можно обнаружить перезимовавшие луковицы, дающие массивный коленчатый столон, на вершине которого располагаются молодые луковицы замещения. Донца этих луковиц окружены пленчатыми чешуями (низовыми листьями), укрывающими зеленую часть побега, листовое влагалище которого не разрослось. В области подземной розеточной части побега различаются корни и зачатки корней. Форма листа имматурного растения узколанцетная, ланцетная, овальная. Размеры листа представлены в таблице 4. В течение жизни имматурное растение значительно увеличивается в размерах. Одновременно идет увеличение числа и размеров чешуй луковицы. Чешуи год от года накапливают все больше питательных веществ, формируя в итоге луковицу, характерную для генеративного периода. Имматурные растения рябчика камчатского способны к вегетативному размножению. Отмечено, что луковицы имматурных растений образуют два столона с молодыми имматурными растениями (1–2 листа у растения) и зачатком побега. Отмечено, что имматурные растения рябчика камчатского производят до двух клонов на плагиотропных столонах длиной до 2 см.
Таблица 4. Морфологические признаки листа Fritillaria camschatcensis в имматурном состоянии (t=2)
Table 4. Morphological features of Fritillaria camschatcensis leaf in an immature state (t=2)
|
Признак |
X |
m |
^min |
^max |
CV, % |
|
Длина листовой пластинки, см |
6.0 ±0.26 |
0.13 |
3.0 |
11.5 |
33.8 |
|
Ширина листовой пластинки, см |
1.9 ±0.09 |
0.04 |
0.9 |
4.1 |
41.5 |
|
Число жилок |
6.6 ±0.29 |
0.14 |
4.0 |
12.0 |
28.8 |
Виргинильное состояние. Наступление этого состояния определяется появлением побега, имеющего стебель с мутовками листьев – сменой типа побега. Зеленый побег имеет 2 мутовки листьев, верхний лист значительно меньше остальных. По окончании вегетации имматурное растение производит столон с почкой возобновления. Весной 2025 года обнаружены имматурные растения рябчика камчатского с луковицей, образованной двумя листовыми влагалищами конической формы и столоном, на конце которого располагался молодой побег, в основании которого обнаружилась почка, сам побег содержал луковицу и мутовки листьев, но не имел бутона (рис. 3, И). Верхняя часть луковицы виргинильного растения сложена тремя крупными чешуями, сильно сросшимися основаниями с донцем (рис. 3, М).
Молодое генеративное состояние . Молодое генеративное растение имеет единственный цветок вне зависимости от пола цветка. Побег его состоит из стебля с мутовками листьев (рис. 3, Л).
Зрелое генеративное состояние . Морфологическое описание зрелых генеративных растений рябчика камчатского из этой популяции приведено нами ранее (Kondakova, Sinelnikova, 2024). Зрелое генеративное растение имеет соцветие вне зависимости от пола цветка. Число крупных, сильно сросшихся с донцем чешуй, образующих верхнюю поверхность луковицы, от 5 до 17 штук в среднем и чаще всего 9.
Cтарые генеративные растения . Данных о существовании растений имевших в прошлом году соцветие, а в нынешнем году единственный цветок нет.
Состояние временно не цветущего растения Gd, субсенильное и сенильное растения
В популяции обнаружены растения, имеющие в составе побега весной луковицу прошлого года (с остовом стебля прошлого года), из почки возобновления которой развился нецветущий побег (Рис. 3, Н). Также встречаются в популяции растения с абортивным цветком. Верхнюю поверхность луковицы временно нецветущих растений слагают от 2 до 6 луковичных чешуй, а сама луковица образуется непосредственно из почки возобновления, находящейся в пазухе верхней запасающей чешуи. Остаток цветоноса прошлого года всегда находится в составе отмирающей луковицы, он прижат к молодой луковице сбоку. В мае 2025 года было обнаружено растение, имевшее перезимовавшую луковицу, состоящую из трех сильно сросшихся верхних чешуй, в пазухе верхней запасающей чешуи располагался остаток стебля, а центральное положение занимал коленчатый столон, вынесший почку возобновления за пределы местонахождения материнской луковицы. На столоне было молодое имматурное растение. Израсходовав значительную часть энергии на цветение, растение может сформировать побег с мужским цветком, либо побег с абортивным цветком, нецветущий побег, а через год после формирования нецветущего побега упростить свою организацию до состояния имматурного растения, сохранив столон и луковичные чешуи. Таким образом, морфологические признаки обнаруженных растений соответствуют более молодым онтогенетическим состояниям, накопление отмерших частей растения (луковицы прошлого года столона, остова цветоноса) проследить трудно, характерного субсенильного и сенильного состояний в онтогенезе вида предположительно нет, вместо этого после обильного плодоношения у растения либо не развивается женская половая система, либо в достаточной степени не развивается цветок (абортивный цветок), либо отрастает нецветущий побег. Нецветущий побег в свою очередь производит столон с еще более омоложенным (имматурным растением), что является процессом омоложения.
Вегетативное размножение . В популяции обнаружены растения рябчика камчатского в имматурном, временно не цветущем, молодом и зрелом генеративном состояниях, имеющие по 2 плагиотропных столона с партикулами. Одна из партикул несла один ланцетный лист на длинном черешке, другая – два листа. Жилок на листьях партикул больше трех, а луковица расположена на столоне и сложена ранней весной пленчатыми луковичными чешуями, верхнюю поверхность луковицы формируют одно или два листовых влагалища, в пазухе которых находится почка возобновления (представляющая собой молодой столон).
Тип онтогенеза. Онтогенез рябчика камчатского, таким образом, осуществляется в ряду поколений особей. Начиная с процесса отмирания влагалища семядоли и образования листа ювенильного растения ежегодно весной (с апреля по середину июня) происходят процессы отмирания побега прошлого года и развития молодого побега из почки возобновления. Полный онтогенез происходит в ряде поколений вегетативно возникших особей (Zhukova, 1995), при этом особи, возникшие из почек, закладывающихся в пазухе верхней запасающией чешуи (V, G1,G2,G3, Gd), или пазухи листового влагалища (Im) год от года состариваются вплоть до состояния временно нецветущего растения (Gd), а затем омолаживаются до состояния имматурного растения. Столон может ветвиться. Столоны, образованные из почек на нижней поверхности донца луковицы (G1, G2, G3, Gd) или из пазухи листового влагалища (Im), несут глубоко омоложенные до имматурного состояния партикулы. В течение лета происходит дезинтеграция клона. Эти факты позволяют отнести жизненный цикл рябчика камчатского к типу Д.
Гендерная структура популяции. Рябчик камчатский – андромоноэцидное растение – соцветия его могут содержать как тычиночные, так и обоеполые цветки, или только тычиночные, либо только обоеполые цветки. Растения с единственным цветком имеют либо тычиночный, либо обоеполый цветок. Растения, имеющие хотя бы один обоеполый цветок, составляют от 45% до 51% цветущих растений. Соотношение растений с пестичными цветками к обоеполым в изученном фитоценозе близко к базовому генетическому закону (1:1) (Zlobin, 2009). Стоит с осторожностью предположить, что условия окружающей среды не оказывают значительного негативного влияния на продуктивность растений.
Онтогенетическая структура популяции .
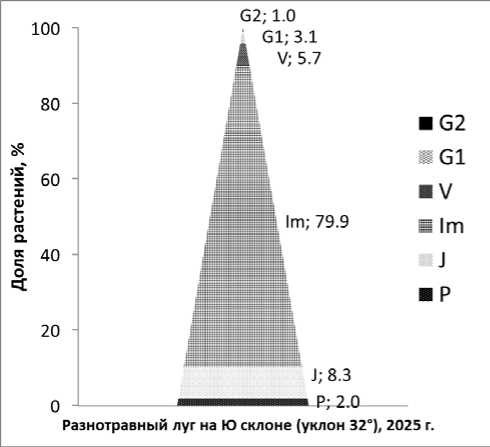
Рис. 4. Онтогенетический спектр популяции Fritillaria camschatcensis разнотравного луга на приморском склоне с уклоном 32°.
Fig. 4. Ontogenetic spectrum of the Fritillaria camschatcensis population of a grassland on a coastal slope with a slope of 32°.
Для вычисления долей от общей численности использовали не среднее арифметическое, а сумму растений на 33 м2 из-за высоких значений коэффициента вариации (табл. 5). Популяция представлена всеми онтогенетическими группами – полночленная (рис. 4). Поскольку сумма долей численности проростков и ювенильных растений уравновешивается суммой долей численности виргинильных, временно нецветущих и генеративных растений популяция отнесена к категории нормальных (Zlobin, 2009). Пик численности приходится на имматурные растения (эксцесс распределения равен 5.8 – распределение остроконечное), что делает эту группу ключевой для исследования продуктивности сообществ, в которых доминирует рябчик камчатский (весной). Коэффициент ассимметрии распределения особей по классам содержаний (онтогенетическим состояниям) 2,4. Высокие значения эксцесса и асимметрия распределения обусловлены спецификой онтогенеза рябчика камчатского.
Поскольку группа имматурных растений образована растениями разного происхождения и по большей части состоит из клонов, которые легко посчитать не за два, а за три растения, популяция выглядит как инвазионная, но таковой на самом деле не является. Поэтому для установления типа популяции предлагаю сравнивать процентное соотношение суммы проростков и ювенильных растений (P+J) с суммой виргинильных, временно нецветущих, молодых и зрелых генеративных растений (V+G1+G2+Gd). В случае если сумма растений P+J будет больше чем, V+G1+G2+Gd, предлагаю считать популяцию инвазионной, в случае равенства двух групп нормальной, в случае, если группа V+G1+G2+Gd составит большую долю численности, чем группа P+J необходимо предположить, что это регрессивная популяция.
Распределение рябчика камчатского по лугу неравномерное, о чем свидетельствует очень высокий коэффициент вариации, как общего числа особей, так и каждой возрастной группы растений (табл. 5). Проростки рябчика на площадках с большим уклоном (около 30°) встречаются единично, на учетных площадках с меньшим уклоном (18°) проростки встречаются гораздо чаще. Семена рассеиваются по склону с большим уклоном, а на выположенных участках семена остаются вблизи рассеявших их коробочек.
Таблица 5. Онтогенетическая структура популяции Fritillaria camschatcensis в 2025 г.
Table 5. Ontogenetic structure of the Fritillaria camschatcensis population in 2025
|
Онтогенетическое состояние |
Число особей на 33 м2 |
я, на 1 м2 |
CV, % |
|
P |
179 |
5 |
175 |
|
J |
751 |
23 |
88 |
|
Im |
7270 |
220 |
56 |
|
V+Gd |
523 |
16 |
64 |
|
G1 |
281 |
9 |
79 |
|
G2 |
95 |
3 |
93 |
|
Общее число особей |
9099 |
276 |
54 |
Онтогенез рябчика камчатского изучен слабо. Первые сведения о морфологических особенностях его сеянцев разного возраста в условиях ботанического сада получены М.В. Барановой (Baranova, 1981). Группа японских исследователей (Nogami et al., 2008й, изучая структуру популяций рябчика камчатского в горах Японии, выделили шесть стадий жизненного цикла: проросток, «однолистное растение», растение со стерильным «воздушным стеблем», растения с мужским цветком, растения с обоеполым цветком, растения с двумя цветками). При подсчете особей на учетных площадках за счетную единицу они приняли рамету – особь вегетативного происхождения. Использование морфобиометрического метода в рамках популяционно-онтогенетического подхода (Smirnova et al., 1976, Zlobin, 1989, Zhukova, 1995) позволило сформулировать диагнозы и ключи для определения онтогенетических состояний рябчика камчатского (табл. 6). Было выявлено, что имматурные растения представляют собой сборную группу, в которую входят:
-
1. Растущие из почек возобновления, образованных ювенильными растениями;
-
2. Омоложенные партикулы, образовавшиеся в процессе вегетативного размножения из почек возобновления, расположенных на нижней поверхности донца луковицы растений (Im, G1, G2, Gd);
-
3. Глубоко омоложенные партикулы, образовавшиеся из почки возобновления, расположенной в пазухе верхней запасающей чешуи временно нецветущего растения.
Нецветущий побег не является признаком единственного онтогенетического состояния. Нецветущий побег свойственен виргинильным и временно нецветущим растениям. За нецветущий побег можно принять побег с абортивным цветком или побег с уничтоженным цветком (обнаружены следы поедания цветков гусеницами). Поэтому, при выявлении онтогенетической структуры популяции виргинильные, временно нецветущие растения и растения с побегом, на котором не было цветка и не было заметно, что это механическое повреждение учтены как виргинильные.
Таблица 6. Диагнозы и ключи онтогенетических состояний Fritillaria camschatcensis
Table 6. Diagnoses and keys of ontogenetic states of Fritillaria camschatcensis
|
Диагноз |
Критерии по А. А. Уранову (кратко) |
Ключи онтогенетических состояний рябчика камчатского |
|
P |
Морфологическая связь с семенем или семядолей |
Игловидная семядоля, имеющая связь с семенем. Линейно-ланцетная семядоля, влагалище семядоли. Жилок нет. |
|
J |
Нет связи с семядолей. Простая организация, отличная от взрослого растения. |
Луковица представлена влагалищем зеленого листа, редко с 1-2 пленчатыми чешуями. Лист узколанцетный, ланцетный на длинном черешке с 1 – 3 жилками. |
Продолжение таблицы 6
|
Диагноз |
Критерии по А. А. Уранову (кратко) |
Ключи онтогенетических состояний рябчика камчатского |
|
IM |
Наличие признаков и свойств, переходных от ювенильных к взрослым: развитие листьев и корневой системы переходного типа |
Лист ланцетный, широколанцетный, овальный с 4 – и более жилками, Луковица закладывается на столоне и образована более чем двумя луковичными чешуйками, центральную часть луковицы образует мясистое листовое влагалище листа, либо два листовых влагалища вегетирующих листьев. Невозможно различить имматурное растение семенного и вегетативного происхождения. Имматурное растение с двумя листьями легко принять за два растения. |
|
V |
Появление основных черт жизненной формы, генеративные органы отсутствуют. |
Побег представлен стеблем с мутовками листьев, верхнюю мутовку венчает 1 мелкий лист, луковица на столоне образована сильно сросшимися тремя верхними чешуйками и многочисленными мелкими чешуями, формирующими боковые и нижнюю поверхности луковицы. Весной и в начале июня побег сохраняет связь с характерной луковицей имматурного растения (Рис 3, М). |
|
G1 |
Появление генеративных органов. |
Появление на побеге единственного цветка. |
|
G2 |
Максимальная семенная продуктивность |
Появление на побеге соцветия. |
|
Gd |
Преобладание процессов отмирания, снижение генеративной функции |
После цветения и плодоношения растения могут сформировать абортивный, рудиментарный цветок. Накоплением отмерших частей является наличие отмершей луковицы с остатком побега прошлого года Побег текущего года имеет облик виргинильного растения, развивается из почки на ранее цветущем, плодоносящем растении, либо на столоне, образовавшимся из луковицы, сложенной лишь тремя сильно сросшимися чешуями. |
Динамику структуры популяций рябчика камчатского изучали в Японии на альпийских лугах на склоне горы Хокусан с 1992 г. по 2003 г. (Nogami et al., 2008). Японские исследователи приводят данные своих многолетних исследований на участке размером 8 м2, где они подсчитывали количество растений разных габитусов, считая их разными онтогенетическими состояниями. Так называемую «однолистную стадию» не подсчитывали. Проростком считалось растение, имеющее на апикальном конце семядоли оболочку семени. Чтобы сравнить данные, полученные нами с данными японских авторов, числа особей с единственными мужским и обоеполым цветками суммированы (табл. 7). В таблице приведены данные за 1994, 1997, 2001, 2003 года и средние значения за 12 лет исследования Японских популяций.
Плотность растений рябчика камчатского в популяции с горы Хокусан в среднем выше в два раза, также популяция отличается в десять раз меньшей долей зрелых генеративных растений.
Другая группа японских исследователей (Shimizu et al., 1998) представила онтогенетический спектр популяций рябчика камчатского с гор Мидзуядзири и Нанрю (табл. 8). Популяции рябчика камчатского с г. Мидзуядзири имеют в два раза более высокую плотность, с г. Нанрю в три раза более высокую плотность растений. Популяция с нюклинских склонов также отличается более высокой долей зрелых генеративных растений, наличием в онтогенетическом спектре группы проростков. Ярко выраженное доминирование имматурных растений свойственно как нюклинской, так и японским популяциям рябчика камчатского.
Таблица 7. Сравнение популяции Fritillaria camschatcensis в северной части ареала вида с популяциями юго-западной границы ареала.
Table 7. Comparison of the Fritillaria camschatcensis population in the northern part of the species' range with the populations of the southwestern border of the range
|
Место, год |
«Нет цветка» / V+Gd, % |
G1, % |
«Два цветка»/ G2, % |
Сумма |
|
Альпийский луг на горе Хокусан, Япония (Nogami et al., 2008) |
||||
|
1994 |
62.3 |
36.8 |
0.3 |
– |
|
1997 |
61.3 |
33.3 |
2.6 |
– |
|
1999 |
60.9 |
37.6 |
1.5 |
– |
|
2001 |
52.4 |
45.3 |
2.4 |
– |
|
2003 |
62.2 |
36.9 |
0.0 |
– |
|
Среднее число за 12 лет ± SD |
247.1 ± 72.4 |
123.4 ± 41.6 38.9 ± 21.3 |
4.5 ± 4.2 |
417.7 ± 99.8 |
|
Средняя доля за 12 лет, % |
58.7 |
39.2 |
1.0 |
– |
|
Разнотравный луг на приморском склоне близ пос. Нюкля, Магаданская обл, Россия |
||||
|
2025 |
58.3 |
31.2 |
10.6 |
– |
Таблица 8. Сравнение популяции Fritillaria camschatcensis в северной части ареала вида с популяциями юго-западной границы ареала.
Table 8. Comparison of the Fritillaria camschatcensis population in the northern part of the species' range with the populations of the southwestern border of the range
|
Стадия |
(Shimizu et al., 1998) |
Магаданская обл. |
||||
|
Мидзуядзири |
Нанрю |
Нюкля |
||||
|
1994 |
1995 |
2025 |
||||
|
n |
% |
n |
% |
n |
% |
|
|
S /P |
0 |
0 |
0 |
0 |
43 |
2.0 |
|
L /J+Im |
4437 /– |
92.8 /– |
6439 /– |
96.2 /– |
1944 /182+1762 |
88.2 /8.3+79.9 |
|
A/V+Gd |
209 |
4.4 |
164 |
2.4 |
127 |
5.7 |
|
G1 |
127 |
2.6 |
72 |
1 |
68 |
3.1 |
|
T/G2 |
1 |
0 |
2 |
0 |
23 |
1.0 |
|
Всего |
4779 |
6690 |
2206 |
|||
В ЫВОДЫ
Исследован жизненный цикл рябчика камчатского в естественных условиях обитания. Популяция разнотравного луга на приморском склоне представлена проростками, ювенильными растениями, имматурными, виргинильными, молодыми и зрелыми генеративными, временно нецветущими растениями. Образование партикул – продуктов вегетативного размножения, свойственно растениям в имматурном, виргинильном, молодом и зрелом генеративном состояниях. Растения образуют до двух столонов с партикулами, омоложенным до имматурного состояния. Виргинильные и временно нецветущие растения при подсчете на учетных площадках необходимо объединять в группу виргинильных растений, из-за высокого риска ошибок при определении онтогенетического состояния растения. В соответствии с классификацией Жуковой, жизненный цикл отнесен к типу Д. Гендерная структура популяции близка к соотношению 1:1. Онтогенетический спектр популяции нормальный, многочисленная группа имматурных растений придает спектру островершинность и положительную ассимметрию, характерную для вида и объясняющуюся особенностями жизненного цикла. По сравнению с южной границей ареала, рассмотренной на примере японских популяций, североохотоморская популяция представлена большей долей зрелых генеративных растений, группой проростков, плотность популяции в разы ниже, чем плотность японских популяции, что объясняется значительным уклоном (в среднем 31°) и рассеянием семян по склону.
Автор благодарит с.н.с. лаборатории ботаники к.б.н. Сазанову Нину Александровну за предоставленную информацию, руководителя лаборатории ботаники ИБПС ДВО РАН к.б.н. Хореву Марию Геннадьевну за помощь в организации исследования и полезные замечания по содержанию рукописи, в.н.с. лаборатории ботаники д.б.н. Синельникову Надежду Вячеславовну за помощь в организации исследования и полезные замечания по содержанию рукописи. Работа выполнена по плановой теме НИР лаборатории ботаники ИБПС ДВО РАН (123032000015–3).
