Онтогенез эфиромасличного растения Monarda didyma L. в условиях Омской области

Автор: Крикливая А.Н., Чермушкина В.А., Шорин Н.В.
Журнал: Вестник Омского государственного аграрного университета @vestnik-omgau
Рубрика: Биологические науки
Статья в выпуске: 4 (24), 2016 года.
Бесплатный доступ
Впервые в условиях Западной Сибири изучены жизненная форма и особенности онтогенеза перспективного эфиромасличного вида монарды двойчатой ( Monarda didyma L. ). Выделены и описаны 4 онтогенетических периода (латентный, прегенеративный, генеративный, постгенеративный) и 10 состояний (покоящиеся семена, проростки, ювенильные, имматурные, виргинильные, молодые генеративные, средневозрастные генеративные, старые генеративные, субсенильные, сенильные). Даны авторские рисунки 4 онтогенетических периодов. В первый год жизни особи M. didyma проходят два онтогенетических периода (латентный и прегенеративный) и 4 состояния (проростки, ювенильные, имматурные, виргинильные). Продолжительность жизни растений в первый год 165-174 дн. Генеративный период длится 3-5 лет. Синфлоресценция - закрытая кисть из 3-9 фрондозно-брактеозных головковидных или колосовидных тирсов. Тирс состоит из супротивно расположенных многоярусных дихазиев. Молодые генеративные, средневозрастные генеративные особи представлены куртиной. В старом генеративном состоянии на 3-4-й год жизни происходит полная партикуляция, образуется клон. У особей, пребывающих в постгенеративном периоде в течение последних двух-трех лет, наблюдается заметное старение. Установлено, что общая продолжительность жизни особей M. didyma в Омской области составляет 7-9 лет. В ходе индивидуального развития в данных почвенно-климатических условиях растения проходят полный онтогенез и формируют длиннокорневищно-стержнекорневую жизненную форму, ее основная структурная единица - разветвленный дициклический удлиненный ортотропный побег с полным и неполным циклом развития.
Монарда двойчатая (monarda didyma l.), онтогенез, длиннокорневищно-стержнекорневая жизненная форма
Короткий адрес: https://sciup.org/142199305
IDR: 142199305 | УДК: 633.81:582.929(571.13)
Ontogeny essential oil plant Monarda didyma L. in the conditions of Omsk region
This article examines the life form and ontogeny specifics of the advanced essential oil variety of Oswego tea ( Monarda didyma L. ) in conditions of West Siberia. The manuscript defines and characterises four ontogeny periods (latent, pre-generative, generative, and post-generative) and ten conditions of the plant (dormant seeds, seedlings, juvenile, immature, virginal, young generative, middle-generative, generative old, sub-senile, senile), also includes illustrations of those periods. During the first year of live varieties of Oswego tea completes two ontogeny periods (latent, pre-generative) and has four conditions (seedlings, juvenile, immature, virginal). First-year plant life varies from 165 to 174 days; the generative period is 3-5 years. Synflorescence of the plant is a closed truss type with 3-9 frondose-bract form capituliform or spicigerous thyrsus, which has a shape of sub-opposite multi-layered dichasial cyme. Young and middle-generative varieties represented by a flower bed. The third and the fourth year of the generative old condition of the plant can be characterised by full particulation and as a period when the slope is emerging. Over the final two or three years of the post-generative period, variety starts dramatically ageing. It was found that the average plant life of M. didym in Omsk region is 7-9 years. The research has revealed that in the course of the individual development in the regional soil and climate conditions the plant covers full ontogeny and builds elongaterhizome-taproot life form, the main structural unit of which is a forked dicyclic elongate orthotropic stem with a full and semi-full development cycle.
Текст научной статьи Онтогенез эфиромасличного растения Monarda didyma L. в условиях Омской области
Мировая флора эфиромасличных растений в настоящее время оценивается в пределах 2500–3000 тыс. видов. Сохранение и расширение ее генофонда, в том числе вовлечение в культуру полезных в практическом отношении видов растений из природных экосистем, бесценно. Флора Северной Америки – весьма богата, это полезный источник интродуцентов из Семейства Lamiaceae для Западной Сибири, представители которой способны в новых экологических условиях проходить полный жизненный цикл, сохранять способность к семенному, вегетативному воспроизводству и формировать высокие урожаи качественного эфиромасличного сырья.
На территории России в естественных условиях вид M. didyma не произрастает. Во всем мире она ценится за эфирное масло, широко используемое в пищевой, парфюмерной промышленности и народной медицине. За последние десятилетия российскими и зарубежными учеными [1 ; 2; 3] проведены комплексные исследования эфирного масла M. didyma , в результате выявлено высокое содержание в его составе тимола, карвакрола и других ценных компонентов, характеризующих M. didyma как перспективное пряно-ароматическое и лекарственное растение, обладающее тонизирующим, антиоксидантным, антисептическим, противовоспалительным, иммуномодулирующим, акарицидным, противопаразитным и противогрибковым действием. Кроме того, растения M. didyma высоко декоративны и популярны во флористике и ландшафтном строительстве. По мнению ряда авторов [4; 5; 6], это прекрасный медонос. В связи с тем что компонентный состав эфирного масла хорошо изучен, исследователи из Польши [7] счи-
тают: в первую очередь следует сосредоточить исследования на селекционной работе по созданию перспективных сортов, пригодных к возделыванию в промышленном производстве.
По литературным данным, сведения о жизненной форме и онтогенезе M. didyma единичны, в Омской области и других регионах России отсутствуют. Следовательно, комплексное изучение нового вида в конкретных почвенно-климатических условиях актуально.
Цель исследования – определить жизненную форму M. didyma , изучить особенности онтогенеза в условиях Омской области.
Объекты и методы
Материал по онтогенезу собран в культуре на экспериментальном участке Ботанического сада ФГБОУ ВО Омский ГАУ (Омская область) с 2007 по 2015 г. Морфологический и статистический анализ проведены не менее чем на 27 модельных разновозрастных особях, полученных из семян местной репродукции. Индивидуальное развитие особей M. didyma изучено согласно концепции дискретного описания онтогенеза по Т.А. Работнову [8]. В соответствии с эколого-морфологической классификацией жизненных форм по особям, находившимся в средневозрастном генеративном состоянии, определена жизненная форма [9]. При характеристике побеговых систем использована терминология, предложенная ведущими российскими учеными-морфологами: И.Г. Серебряковым, Л.Е. Гатцук, Е.Л. Нухимовским [10–13]. Соцветия описаны согласно терминологии Т.В. Кузнецовой и W. Troll [14; 15]. Количественные данные обработаны с помощью пакета программ Microsoft Excel.
Результаты исследований
Онтогенез M. didyma в условиях Омской области представлен на рис. 1.
Семена (эремы) M. didyma вскрывающиеся, формируются в ценобиях. Ценобии четырехкамерные, состоят из 4 эремов овально-продолговатой формы темно-коричневого цвета. Семена с хорошо выраженным зародышем, окруженным толстым слоем эндосперма (80–85%) и тонкой семенной кожурой, неплотно прилегающей к эндосперму. Длина семян 1,51–1,64 мм, ширина 0,77–0,81 мм. Масса 1000 семян – 0,49 г. Оптимальная температура их прорастания 18–20°С [16]. Плодоношение растянуто на 30– 60 дн. Массовое созревание происходит во 2-й половине сентября. Семена не имеют периода покоя, могут прорастать в год созревания.
Прорастание семян надземное. Высота вегетативного побега 1,2–4,4 см. На первичном удлиненном побеге проростка (p) развертываются два семядольных листа и 1–3 пары настоящих листьев на коротких черешках. Листорасположение накрест супротивное. Верхушечная почка открытого типа. Гипокотиль хорошо выражен, 0,3– 2,1 см длиной, на нем образуются 2–6 зачатков придаточных корней. Семядольный лист округлой формы с выемчатой верхушкой и округлым основанием, имеющим стреловидные выступы до 0,1 см. Верхняя сторона семядольного листа светло-зеленая, нижняя антоцианового цвета, что характерно для большинства видов Семейства Lamiaceae . Лист с обеих сторон опушен железистыми волосками. Длина и ширина листовой пластинки семядоли 0,2–0,5 см, длина черешка 0,1–0,4 см. Эпикотиль длиной 0,4–1,4 см. Листовая пластинка настоящих листьев первого метамера длиной 0,4– 1,3 см, шириной 0,4–1,1 см, имеет широкояйцевидную форму, заостренную верхушку, округлое основание и волнисто-зубчатый край. Длина черешка 0,3–0,8 см. В пазухах всех листьев закладывается по 1 пазушной почке открытого типа. Главный корень ветвится и формирует 8–14 боковых корней I–II порядка. Глубина залегания главного корня – 1,9–6,4 см. Длительность состояния проростка – 5–20 дн.
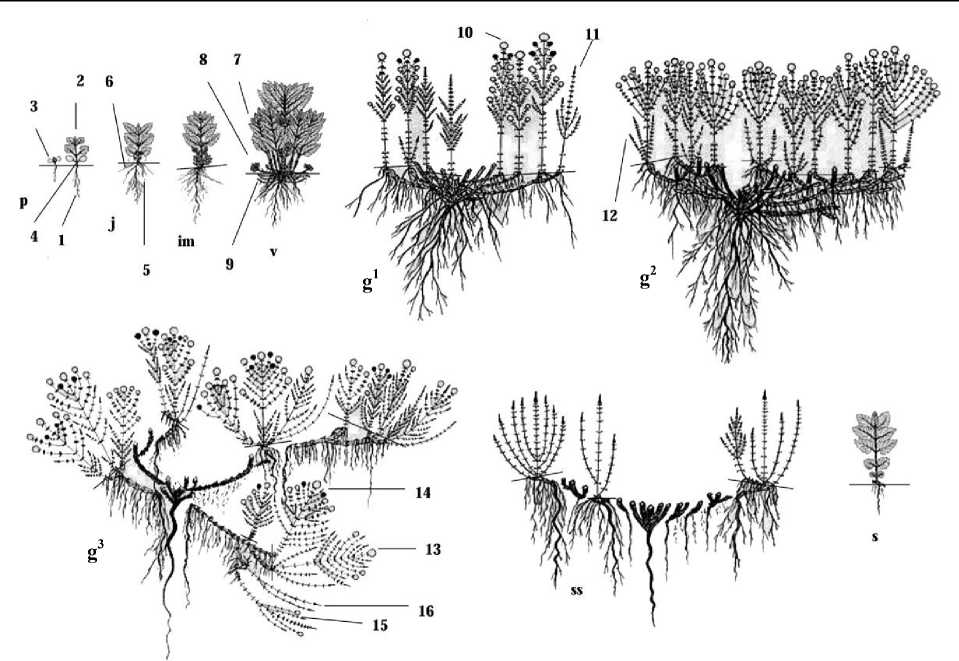
Рис. 1. Онтогенез Monarda didyma L. в условиях Омской области: количество корней, побегов на рисунке указано условно; ––– уровень почвы; « ––– » – неживые или разрушенные части особи; 1 – главный корень; 2 – первичный уединенный побег;
3 – семядольные листья; 4 – гипокотиль; 5 – боковые корни; 6 – придаточные корни;
7 – экспоненциально нарастающий вегетативный побег II порядка; 8 – анизотропный корневищно-удлиненный побег III порядка; 9 – плагиотропный удлиненно-корневищный побег
III порядка; 10 – дициклический ортотропный удлиненный генеративный побег; 11 – дициклический ортотропный удлиненный вегетативный побег; 12 – моноциклический неветвящийся ортотропный удлиненный вегетативный побег V порядка; 13 – полициклический ортотропный удлиненный генеративный побег III порядка; 14 – трициклический ортотропный удлиненный генеративный побег IV порядка; 15 – трициклический ортотропный удлиненный вегетативно-генеративный побег IV порядка; 16 – дициклический вегетативный побег V порядка
После отмирания семядольных листьев особи переходят в ювенильное состояние (j), которое длится от 20 дн. до 1,5 мес. Нарастая моноподиально, первичный одноосный четырехгранный опушенный побег несет 4–9 пар настоящих листьев. Пазушные почки семядольных и зеленых листьев 1–2-го метамера трогаются в рост, образуют силлептические побеги II порядка, на их оси располагаются 1–3 пары ассимилирующих короткочерешковых листьев. Выше по оси главного побега в пазухах вновь образованных листьев также закладываются почки. Зеленые листья третьего и последующих метамеров материнского побега имеют листовую пластинку яйцевидной формы длиной 1,0–2,9 см, шириной 1,2–2,3 см, черешок 0,6–1,4 см, заостренную верхушку, округлое основание и зубчатый край. Для всех листьев характерно рассеянное опушение железистыми волосками. На эпикотиле формируются 3–6 зачатков придаточных корней, что приводит к его расширению и изменению размера в среднем до 0,47 см. У ювенильных растений главный корень втягивает гипокотиль в почву, таким образом, семядольный узел оказывается на поверхности почвы, при этом гипокотиль, как и эпи- котиль, уменьшается в размере до 0,2–1,1 см. На нем насчитывается 5–9 придаточных корней. Главный корень хорошо выражен, длиной 6,4–12,1 см, светло-коричневого цвета, разрастается в глубину и ширину. На его оси располагается 15–26 боковых корней I–III порядков.
В онтогенетическое имматурное состояние (im) растения переходят на 60–75-й день, длится оно 1–1,5 мес. Первичный побег длиной 10,4–13,6 см состоит из 10–12 метамеров. Листья с 6 по 9-й метамер имеют схожую с взрослыми особями овальноланцетную листовую пластинку длиной 2,5–3,7 см, шириной 1,2–2,8 см, зубчатый край, заостренную верхушку, округлое основание, черешок длиной 0,5–1,0 см. В пазухе семядольного узла и листьев с 1 по 7-й метамер закладывается по одной сериальной почке. Ранее заложенные в пазухах 3–7-го метамера боковые почки трогаются в рост, продолжается экспоненциальное нарастание числа осей II порядка. На побегах II порядка семядольного узла и зеленых листьев 1–2-го метамерa материнского побега закладывается по одной пазушной почке. На главном побеге постепенно начинают отмирать листья снизу до 7-го метамера. На междоузлиях 1–2-го метамера образуются 3–6 придаточных корней. После отмирания 1–2 пары листьев главного побега происходит втягивание семядольного узла с его побегами в почву главным и придаточными корнями таким образом, что второе междоузлие оказывается на поверхности почвы и укореняется. В данном онтогенетическом состоянии наблюдается значительное разрастание корневой системы. На главном корне в среднем насчитывается до 23 боковых корней. В 2–3 раза увеличивается число придаточных корней. Корневая система смешанная, глубина залегания 13,0–15,2 см.
Виргинильное состояние (v) наступает на 100–120-й день и длится 45–54 дн. до конца октября. Виргинильные особи представлены первичным кустом, состоящим из 9–13 моноциклических ортотропных удлиненных вегетативных побегов I–II порядка, 4–8 анизотропных корневищно-удлиненных побегов III–IV порядка и 2–4 плагио-тропных длиннокорневищных побегов II–IV порядка. Диаметр куста 8,0–11,0 см. На оси первичного побега длиной 36,0–45,0 см располагаются зеленые листья такой же формы, как и в имматурном состоянии, но с большими размерами. Длина листовой пластинки 3,4–4,5 см, ширина 2,2–2,9 см, длина черешка 0,9–1,4 см. Материнский побег с побегами ветвления погружается в почву до 3-го метамера системой главного и придаточных корней. Таким образом, 4-е междоузлие первичного побега и 1–3-е междоузлие побегов II порядка оказываются на поверхности почвы и укореняются. Сериальные почки, расположенные в пазухах 1–3-го метамера подземной части первичного побега, трогаются в рост, образуя по одному плагиотропному длиннокорневищному побегу, несущему на своей оси 4–7 пар чешуевидных листьев с пазушными почками. В подземной части разветвленного первичного побега из пазушных почек побегов II порядка развиваются 4–8 анизотропных корневищно-удлиненных побегов длиной 3–8 см, несущие как чешуевидные, так и переходные мясистые темно-зеленые листья, и 4–6 плагиотропных длиннокорневищных побегов длиной 2–4 см с чешуевидными пленчатыми листьями. В пазухах листьев обоих типов побегов III порядка регулярно закладываются почки. В зависимости от погодных условий года 1–2 почки 4–5-го метамера трогаются в рост, приводя к ветвлению подземных частей анизотропных корневищно-удлиненных и плагиотропных длиннокорневищных побегов до IV порядка. Одновременно с ростом побегов IV порядка на узлах корневища формируются по 2– 3 придаточных корня. В октябре надземная часть главного и побегов II порядка засыхает и отмирает. Анизотропные корневищно-удлиненные побеги III порядка полегают, укореняются, засыпаются остатками старых листьев, уходят под снег с зелеными ли- стьями. Терминальная почка – открытого типа. Таким образом, в первый год формируется геофильная удлиненная часть дициклического побега, который на будущий год зацветет. Корневая система смешанного типа: сохраняется главный корень, ветвящийся до III порядка, и придаточные корни I–II порядка, расположенные в узлах разветвленного первичного побега, в приземных узлах анизотропных корневищно-удлиненных побегов и в 5–7-м узлах плагиотропных длиннокорневищных побегов. Как и в имма-турном состоянии, у виргинильных особей значительно увеличивается число придаточных корней, в среднем до 50 шт., большинство из них выполняет контрактильную функцию. Глубина залегания корневой системы – 13,5–18,0 см.
В первый год жизни особи M. didyma проходят 2 онтогенетических периода (латентный и прегенеративный) и 4 состояния (проростки, ювенильные, имматурные, вир-гинильные). Продолжительность жизни особей в первый год – 165–174 дн.
Молодые генеративные особи (g1) во второй год жизни формируют куртину, которая состоит из первичного куста и укоренившихся парциальных побегов III–IV порядка. Куртина представлена дициклическими ортотропными удлиненными 3–12 генеративными и 1–3 вегетативными побегами. Особи в высоту достигают 96,4 –140,0 см, в диаметре 14,0–18,2 см. Генеративный побег состоит из 11–14 метамеров, в пазухах 5–10-го метамера развертываются паракладии. На оси располагаются 5–9 пар длинночерешковых листьев овально-ланцетной формы. Нижние зеленые листья отмирают с конца мая, во время заложения генеративных органов на конусе нарастания. Листья срединной формации генеративного побега длиной 5,6–9,7 см, шириной 2,5–4,2 см; длина черешка 1,2–2,1 см. Синфлоресценция – закрытая кисть (рис. 2), состоит из 3–9 фрондозно-брактеозных головковидных тирсов. Тирс (рис. 2) представлен супротивно расположенными многоярусными дихазиями. Нередко дихазий третьего или более высокого порядка с резко неравномерными боковыми осями редуцируется до мо-нохазия. Листья верхней формации – переходные (3 пары), по центральной жилке и краям окрашены в тот же цвет, что и цветок. Брактеи, прилегающие к чашечкам, узколанцетные, пленчатые, по краю коротко мягко реснитчатые. Чашечка трубчатая, пятизубчатая, длиной 0,7–1,0 см, слегка опушена. Шиловидные зубцы чашечки окрашены в фиолетовый цвет. Венчик двугубый, верхняя губа прямая, нижняя изогнута, с тремя неровными лопастями, центральная имеет язычкообразный выступ, рассеченный надвое. Длина венчика 2,3–3,0 см. Тычинки две – выступающие. Цветки фиолетового или лилового цвета с сильным запахом мяты и лимона. Общее число цветков на одно соцветие в среднем 163,0. Цветение молодых генеративных особей растянутое, начинается с верхушечного головковидного тирса в III декаде июля – I декаде августа и продолжается до II декады сентября. Цветки распускаются акропетально. Часто соцветия, располагающиеся ниже главного, в пазухах 10–12-го метамера не переходят к цветению, засыхают и отмирают.
Вегетативные побеги развиваются из пазушных почек базальных участков побегов II и III порядков, реже из сериальных почек главного побега (если предшествующая осень была затяжной и теплой). На геофильных участках вегетативных побегов меньшее число метамеров (2–4). Вегетативные побеги в надземной части мезотонно ветвятся. На их оси располагаются 13–18 удлиненных метамеров. К моменту распускания первых цветков на вегетативном побеге образуется 8–11 пар зеленых листьев такой же формы, как у генеративных побегов.
У единичных генеративных побегов наблюдается базитонное ветвление вегетативными побегами обрастания, развертываются они из заложенных в мае пазушных почек на их базальных укорененных участках.
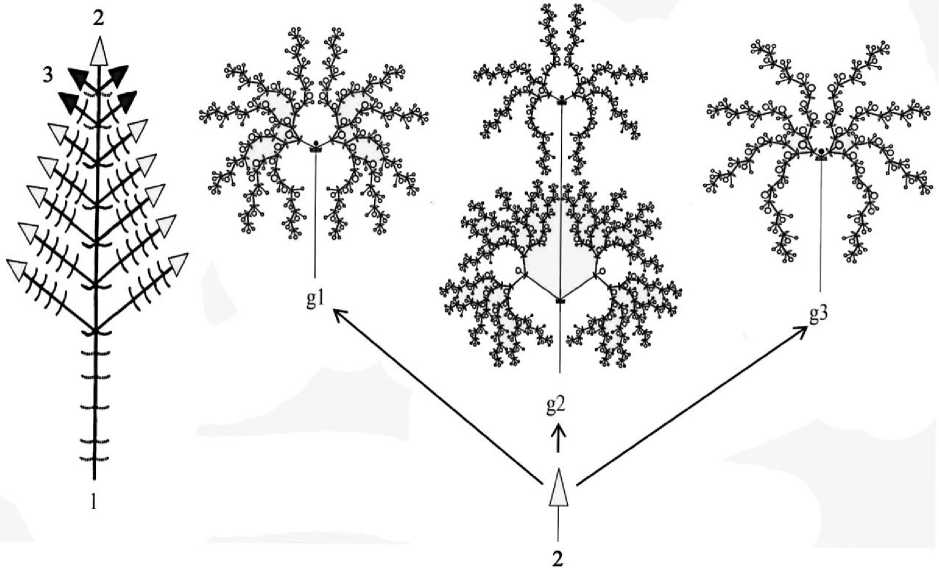
Рис. 2. Схема строения тирса в генеративном периоде по онтогенетическим состояниям:
1 – синфлоресценция – закрытая кисть из тирсов; 2 – фрондозно-брактеозный головковидный тирс;
3 – сформированный тирс, но не переходящий к цветению, g 1, 2, 3 – схема строения тирса в генеративном периоде по онтогенетическим состояниям: g1 – головковидный, g2 – колосовидный, g3 – головковидный; – зеленый лист;.1. – опавший лист; ср – цветок; – дихазий
| I о-ҒЧғто с редуцированной апикальной меристемой и 3 парами зеленых листьев нижней формации;
T – брактеи, ниже располагаются укороченные междоузлия
Одновременно с формированием головковидных тирсов парциальных побегов III порядка на геофильных участках корневищ в пазухах чешуевидных листьев закладывается по одной сериальной почке, в это же время трогаются в рост 2–4 перезимовавшие основные почки. Таким образом формируются геофильные участки побегов замещения IV порядка, которые в сентябре в подземной части ветвятся до V порядка, образуя небольшие геофильные участки новых побегов. Нереализованные почки остаются спящими или отмирают. В подземной части первичного куста плагиотропные длиннокорневищные побеги прошлого года, преимущественно сформированные из сериальных почек, не переходят к вегетации. Наблюдается загнивание конуса нарастания с последующим его отмиранием. Корневая система смешанная и состоит из главного корня и большого числа придаточных корней (41–65 шт.) I–III порядков ветвления, располагающихся плотно на узлах корневища по 4–6 шт., большая их часть сосредоточена в апогеотропной части генеративных побегов. Глубина залегания корневой системы 14,0–19,8 см. Длительность онтогенетического состояния – 1 год.
Средневозрастные особи (g2) представлены куртиной, в ее пределах обосабливаются 3–4 парциальных куста и 2–3 дициклических парциальных побега IV–V порядка. Парциальные кусты образованы 1–2 дициклическими ортотропными удлиненными генеративными побегами возобновления IV порядка и 2–3 силлептическими неветвящи-мися ортотропными удлиненными вегетативными побегами обрастания V порядка. В этом состоянии центральная часть куста начинает разрушаться, но морфологическая целостность особи сохранятся. Средневозрастные особи высотой 94–117 см, в диаметре 20,0–29,0 см, формируют 3–4 центра разрастания. Генеративный побег представлен 10–14 удлиненными метамерами с 4–10 парами длинночерешковых зеленых листьев. Паракладии длиной 19–35 см, располагаются в пазухах 4–12-го метамера. Синфлорес-ценция как у молодых генеративных особей, но с большим числом головковидных тирсов (3–17). С возрастом отмечено усложнение формы тирса, у 60% особей тирс становится колосовидным (рис. 2.2). Увеличивается число цветков в соцветии в среднем до 272,0. Средневозрастные особи зацветают раньше молодых генеративных на 5–7 дн., обильное цветение продолжается до сентября. В зависимости от погодных условий года вегетативные побеги могут отсутствовать. На геофильных участках годичных побегов из перезимовавших пазушных почек 3–5-го метамера развиваются 2–6 анизотропных корневищно-удлиненных побегов и плагиотропных длиннокорневищных побегов V порядка, длиной 1,5–14 см. В июле 1–2 пазушные почки длиннокорневищных побегов III порядка трогаются в рост, что приводит к их ветвлению до IV порядка (рис. 1). Корневая система смешанного типа. На узлах молодого корневища плотно друг к другу располагаются многочисленные придаточные корни. У парциальных побегов и кустов различают 4 типа придаточных корней: опорные, ростовые питающие, удлиненные ростовые, контрактильные. Один из придаточных корней утолщается и углубляется, выполняя опорную функцию. Главный корень сохраняется. Глубина залегания корневой системы – 18–38 см. Придаточные укороченные сосущие корни начинают отмирать. Длительность средневозрастного онтогенетического состояния – 1–2 года.
Старые генеративные особи (g3) образуют клон. В результате перегнивания центральной части первичного куста, осей II порядка и их корневищ происходит полная партикуляция. Как правило, в старом генеративном состоянии во второй половине лета у особей на 3–4-й год жизни полностью отмирает главный корень и один из придаточных корней принимает на себя его функцию. Клон состоит из 5–7 неомоложенных пар-тикул. Старые генеративные особи по высоте ниже, чем в предыдущих генеративных состояниях, на 22–24 см. Парциальные кусты расставлены примерно на одинаковое расстояние друг от друга. В образовании парциальных кустов принимают участие ди-циклические и трициклические ортотропные удлиненные генеративные побеги. В состав генеративного побега входит 10–15 удлиненных метамеров с 3–10 парами зеленых листьев, схожих по форме с молодыми генеративными и средневозрастными особями, но отличающимися меньшими размерами. Длина черешка 0,5–1,2 см, длина листовой пластинки 4,0–8,4 см, ширина 1,8–3,4 см. В надземной части генеративный побег ветвится, из пазушных почек 1–6-го метамера развиваются побеги обрастания, а в междоузлиях 7–11-го метамера – паракладии. В синфлоресценции снижается число головковидных тирсов до 1–5, в них значительно сокращается число цветков, в среднем 83,0. У большинства особей преобладает головковидная форма тирса (рис. 2), у 10% встречается колосовидная. Базальные участки генеративных побегов находятся на поверхности почвы, небольшое число образовавшихся придаточных корней теряют контрактильную способность, утолщаются, выполняя опорную функцию. Пазушные почки в базальной части годичных побегов развертываются в вегетативные побеги обогащения, достигают в высоту 3,5–13 см, нередко засыхают и отмирают, тем самым снижается способность к образованию побегов разрастания. Возобновление в следующем году происходит из спящих почек на сохранившихся живых участках корневища. Корневая система состоит 41–65 придаточных корней I–III порядка. Условный возраст парциальных кустов – 1–2 года.
Субсенильные особи (ss) представлены системой из 2–3 парциальных кустов и вегетативных парциальных побегов (1–2), расставленных далеко друг от друга. Особи в высоту достигают 45–70 см. В составе парциальных кустов 1–3 ортотропных вегетативных побега, которые в надземной части ветвятся, формируя 2–4 побега обогащения. Вегетативные побеги развиваются из спящих почек на сохранившихся живых участках корневищ. На оси побега возобновления располагается 9–18 метамеров и 7–11 пар короткочерешковых зеленых листьев. У листьев появляются черты ювенильного и имма-турного состояний, листовая пластинка овально-яйцевидной формы с зубчатым краем, верхушка заостренная, основание округлое. Длина листовой пластинки – 2,8–6,8 см, ширина – 2,1–3,0 см, длина черешка – 0,5–1,1 см. На апогеотропных участках вегетативных побегов образуются придаточные корни, отмирающие осенью вместе с надземной частью. Корневая система состоит из придаточных корней, образованных в старом генеративном состоянии, один из них утолщается и заякоривается. Условный возраст партикул – 1–2 года.
У сенильных особей (s) возобновление происходит из спящих почек живых участков многолетнего корневища, формируется парциальный побег, за счет которого они существуют в течение года. В данном онтогенетическом состоянии наблюдается вторичное проявление черт ювенильных особей, проявляющееся в размерах, длительности цикла развития и типе вегетативного побега, а также форме его листьев. Как правило, на ортотропном удлиненном побеге длиной 13–18 см располагается 6–7 пар зеленых листьев разной формы на коротких черешках 0,2–0,4 см. Листья первого метамера имеют почковидный тип листовой пластинки с тупой верхушкой и сердцевидным основанием; у листьев второго и третьего метамера обратно – сердцевидный тип листьев с заостренной верхушкой и сердцевидным основанием, вышерасположенные листья имеют округло-яйцевидную форму, заостренную верхушку и сердцевидное основание. Листья срединной формации синильных особей, в отличие от субсенильных, помимо смены формы листовой пластинки, имеют меньшие размеры – длина 1,7–2,0 см, ширина 1,2–4,4 см. Корневая система состоит из 2–6 придаточных корней, сосредоточенных в узлах функционирующего корневища. Условный возраст парциальных побегов – 1 год.
Закл ючение
Взрослая особь M. didyma в условиях Омской области формирует длиннокорне-вищно-стержнекорневую жизненную форму, ее основная структурная единица – разветвленный дициклический удлиненный ортотропный побег с полным и неполным циклом развития. В течение жизни растения проходят 4 онтогенетических периода и 10 состояний. В ходе индивидуального развития в данных почвенно-климатических условиях растения M. didyma проходят полный онтогенез. Общая продолжительность жизни особей в Омской области – 7–9 лет.
A.N. Kriklivaja, V.A. Cherjomushkina, N.V. Shorin
Ontogeny essential oil plant Monarda didyma L. in the conditions of Omsk region
Список литературы Онтогенез эфиромасличного растения Monarda didyma L. в условиях Омской области
- Изучение монарды как источника БАВ/А.А. Серкова //2-й Междунар. симп. «Новые и нетрадиционные растения и перспективы их практического использования»: материалы докладов. -Пущино, 1997. -Т. 2. -С. 88-90.
- Исследование химического состава эфирного масла Monarda fistulosa L. и Monarda didyma L./Р.В. Опирин //Химия растительного сырья. -2000. -№ 3. -С. 19-24.
- Composition of Leaf and Flower Essential Oil from Monarda didyma L. Cultivated France/A.P. Carnat, J.L. Lamaison, A. Remery//Flavour and Fragrance Journal. -1991. -Vol. 6. -P. 79-80.
- Мартынов А.М. Некоторые итоги интродукции монарды в условиях Пятигорска/А.А. Мартынов, Д.А. Муравьева//Науч. докл. высшей школы. Биолог. науки. -1973. -№ 2. -С. 82-84.
- Дрягина И.В. Монарда -растение индейцев Северной Америки/И.В. Дрягина, Л.Ю. Кан//Картофель и овощи. -1996. -№ 5. -С. 17-18.
- Effect of floral orifice width and shape on Hummingbird-flower interactions/C.E. Smith et al.//Jecologia. -1996. -Vol. 106. -P. 482-492.
- Identification of potential sources of thymoquinone and related compounds in Asteraceae, Cupressaceae, Lamiaceae, and Ranunculaceae families/J. Toborsky et al.//General European Journal of Chemistry. -2012. -Vol. 10. -P. 1899-1906.
- Работнов Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах/Т.А. Работнов//Тр. Бот. ин-та АН СССР. Сер. 3. Геоботаника. -М.: Из-во АН СССР, 1950. -Вып. 6. -С. 179-196.
- Серебряков И.Г. Экологическая морфология растений/И.Г. Серебряков. -М.: Высш. шк., 1962. -378 с.
- Серебряков И.Г. Морфология вегетативных органов высших растений/И.Г. Серебряков. -М.: Сов. наука, 1952. -391 с.
- Гатцук Л.Е. Жизненные формы в роде Hedyzarum L. и их эволюционные взаимотношения/Л.Е. Гатцук//Бюл. МОИП, отд. биол. -1967. -Т. 72. -Вып. 2. -С. 53-64.
- Нухимовский Е.Л. Структурная организация и морфогенез некоторых семенных растений: автореф. дис. … канд. биол. наук/Е.Л. Нухимовский. -Москва, 1970. -15 с.
- Нухимовский Е.Л. Осевая и побеговая системы семенных растений/Е.Л. Нухимовский//Изв. ТСХА. -1971. -Вып. 1. -С. 54-66.
- Кузнецова Т.В. Соцветия. Морфологическая классификация/Т.В. Кузнецова, Н.И. Пряхина, Г.П. Яковлев. -СПб., 1992. -126 с.
- Troll W. Die Infloreszenzen. Typologie and Stellung im Aufban des Vegetations koppers B. -Vena, 1964. -T. III. -Bd 1. -P. 73-75.
- Анищенко И.Е. Нетрадиционные пряно-ароматические растения семейства Lamiacea в Башкортостане/И.Е. Анищенко//Вестн. ОГУ. -2009. -№ 6. -С. 35-38.
