Онтоморфогенез клевера тихоокеаского на территории Лазовского государственного заповедника им. Л. Г. Капланова

Автор: Калинкина В.А.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: флора
Статья в выпуске: 1-3 т.12, 2010 года.
Бесплатный доступ
Изучено становление жизненной формы клевера тихоокеанского (Trifolium pacificum Bobr.), произрастающего на территории Лазовского государственного заповедника им. Л.Г. Капланова. Установлено, что жизненная форма в процессе развития изменяется: в надземной сфере прямостоячий побег, характерный для особей первого года жизни сменяется на стелющийся, в подземной сфере - стержнекорневая жизненная форма сменяется на длиннокорневищно-вторично-стержнекорневую. Такое строение особей сохраняется на большем отрезке жизненного цикла клевера тихоокеанского.
Жизненная форма, онтоморфогенез
Короткий адрес: https://sciup.org/148199017
IDR: 148199017 | УДК: 633.325
Ontomorphogenesis of Trifolium pacificum on territories of Lazovskiy State Reserve named after L. G. Kaplanov
Becoming a life form of Trifolium pacificum Bobr., growing on the territory of Lazovskiy state reserve named after L.G. Kaplanov is studied. It is established, that the life form during development changes: in elevated sphere upright runaway, characteristic for species of the first life year is replaced on creeping, in underground sphere - tap-root life form is replaced on long-root- secondary-tap-root. Such structure of species is preserved on a greater piece of Trifolium pacificum life cycle.
Текст научной статьи Онтоморфогенез клевера тихоокеаского на территории Лазовского государственного заповедника им. Л. Г. Капланова
Широко известно, что виды семейства бобовые ( Fabaceae ) рода Trifolium L. являются хозяйственно-ценными растениями, при этом биология некоторых из них до сих пор не изучена. Одним из таких видов является клевер тихоокеанский ( Trifolium pacificum Bobr.). Вид имеет дальневосточный ареал обитания, на территории Дальнего Востока России произрастает в трех флористических районах, имеющих выход к морю: Уссурийском, Южно-Сахалинском и ЮжноКурильском [2, 3, 6-8]. Характерными местами произрастания вида являются склоны морских террас, берег моря (зона супралиторали), каменистые осыпи, изредка - лесные прибрежные склоны.
Последнее время территория Приморского края находится в процессе массовой застройки. Отсутствие обширных ровных площадей в районах с преимущественно горным рельефом приводит интенсивному поиску новых площадок под строительство. Для многих людей одними из желаемых мест проживания являются чистые, еще мало освоенные приморские территории. Массовое стремление человека освоить остатки нетронутых прибрежных районов приводит к сокращению естественного места обитания растений супралиторали. Единственной защитой для растений являются особо охраняемые территории: заказники, заповедники и т.д. Лазовский район расположен в юго-восточной части Приморского края на восточных склонах Си-хоте-Алиня. География района такова, что часть его территории имеет выход в Японское море. Именно здесь находиться один из 6 приморских
заповедников – Лазовский государственный заповедник им. Л.Г. Капланова.
Для изучений хозяйственно-ценных растений в целях сохранения их природных популяций, а также введения их в культуру необходимо знать все аспекты биологии вида. В данной работе охарактеризован онтоморфогенез клевера тихоокеанского, на территории еще не затронутой человеческой деятельностью.
Изложенные данные о возрастном изменении T. pacificum сделаны главным образом на анализа растений взятых в 2007 г. на территории бухты Проселочная, являющейся частью Лазовского заповедника. Специфический комплекс факторов среды отражается на индивидуальном развитии растений: сильные ветра, влияние приливов и отливов накладывают отпечаток на развитие надземной сферы, необходимость закрепиться в субстрате и особая песчаная почва с мелкими камнями оказывает влияние на формирование подземной сферы. Кроме отбора образцов в течение 2007-2008 гг. проводились наблюдения за развитиями особей в природных популяциях. Для получения дополнительных сведений по морфологии вида сведений нами были использованы гербарные материалы, хранящиеся в крупных гербариях России: Ботанического института РАН им. В.Л. Комарова (LE), Главного ботанического сада РАН (VYF), Биологопочвенного института (VLA) и Лазовского государственного природного заповедника им. Л.Г. Капланова. Исследовано 250 экземпляров T. pacificum . В работе применены биоморфологи-ческий, онтогенетический и экологогеографический методы.
Исследования и описание жизненной формы (ЖФ) выполнено по методике И.Г. Серебрякова [13, 14], Т.И. Серебряковой [15] и их учеников. Этапы развития особей охарактеризованы с использованием концепции дискретного описания онтогенеза, предложенной Т.А. Работ-новым [10, 11] и детально разработанной А.А. Урановым [16].
Латентный период. Семена T. pacificum бобовидные, гладкие, зеленые или коричневозеленые, для семян этого вида клевера характерно автобарохорны способ распространения [4]. Семенам свойственно состояние органического покоя. Прорастание семян надземное, гипокоти-лярное.
Виргинильный период. Проросток (p) . Прорастание T. pacificum наблюдается в июне– июле. В связи с тем, что прорастание надземное, первыми на дневную поверхность появляются овальные, мясистые, зеленые семядоли. Выше семядолей развивается несколько настоящих листьев. Первый лист – сложный однолисточко-вый, округлой или округло-продолговатой формы, с выемкой на верхушке [12]. Последующие 4-5 листьев тройчатосложные, их листочки плотные, обратнояйцевидные с небольшой выемкой на верхушке, зубчатые по краю, более широкие в верхней трети. В первый год клевер тихоокеанский образует однолетний удлиненный прямостоячий побег, начало которому дает зародыш семени. Подземная сфера в первый год представлена системой главного корня, развитого из зародышевого корешка.
Ювенильное возрастное состояние (j) . Отмирание семядольных листьев, начинающее в июле-августе, характеризует начало ювенильного возрастного состояния. В это время у особей формируется еще 5-8 тройчатосложных листьев. Общая длина побега составляет 5,3 см. Положение побега в пространстве сохраняется. В конце вегетационного периода в пазухах семядольных листьев и 1-3-го настоящих листьев закладываются одиночные почки возобновления, редко в пазухе 1-го листа формируется группа сериальных почек. Осеннее отмирание побега приводит к образованию первой главы каудекса, которая является основой многолетнего побегового тела растения. Подземная сфера представлена системой главного и боковых корней 1-2 го порядка. За счет контрактильной деятельности главного корня происходит втягивание первой главы кау-деска с почками возобновления в почву, а также развитие на гипокотиле поперечных борозд.
На второй год растение переходит в им-матурное возрастное состояние (im), которое характеризуется: сменой типа нарастания и положения побега в пространстве и началом развития каудекса. В надземной сфере за один год формируется 1, редко 2 удлиненных стелющихся побега с тройчатосложными листьями. Начало побегам дают почки возобновления, расположенные на главе каудекса первого года жизни. Развитие стелющихся побегов является одной из приспособительных особенностей растений супралиторали [1]. В пазухах 1–3 нижних листьев формируются одиночные почки возобновления. Подземная сфера практически не изменяется, наблюдается сохранение главного корня и некоторое его утолщение в верхней трети. За счет вкручивания корня в почву на его поверхности формируются продольные складки.
Переходя на 3-4 год в вегетативное возрастное состояние (V) у особей T. pacificum происходят существенные морфологические изменения, как в надземной, так и в подземной сферах. Развивающиеся в этом возрастом состоянии побеги характеризуются неполным циклом развития. Они все также простые или слабо ветвящиеся в зоне обогащения, стелющиеся. С этого возрастного состояния основная масса листьев (7-9) на побеге пальчатосложные, лишь 1-3 нижних листа тройчатосложные. В подземной сфере наряду с сохранением системы главного корня образуются корневища. Формирование последних свойственно большинству растений побережья [1]. У клевера тихоокеанского в возрасте 2-3 (4) лет из почки, расположенной на главе материнского каудекса, развивается небольшое гипогеогенные симподиально нарастающее корневище. Развитие растений в дальнейшем приводит к тому, что годичные побеги формируются из почек, расположенных на корневище и главах материнского каудекса. Образование в течение нескольких лет побеговой системы на конце корневища приводит к образованию дочернего каудекса или парциального куста. За счет формирования последнего происходит увеличение общей массы особи. Необходимость закрепиться в субстрате приводит к развитию мощного главного корня, длина его достигает 20 (30) см, а диаметр основания 2 (2,2) см.
В связи с ранним формированием корневища и слабой выраженностью его годичных приростов определить длительность возрастных состояний является проблематичным, поэтому время, в течение которого особи прибывают в том или ином возрастном состоянии мы не указываем.
Группа генеративных растений T. pacificum не однородна, поэтому в генеративном периоде мы выделяем 3 возрастных состояния: G1, G2, G3.
Начало цветения происходит в молодом генеративном возрастом состоянии (G1), при этом структура особи весной нередко не отличаются от описанной выше. Формирующиеся генеративные побеги простые, стелющиеся, единичные (1-2), расположены в основании наиболее молодых глав материнского каудекса, состоящие из 15-17 метамеров. Листья побега пальчатосложные. Соцветия пазушные, головчатые; цветки малиновые. Материнский каудекс, сформированный в начале жизни особи, в этом возрастном состоянии многоглавый, рыхлый. В подземной сфере сохраняется система главного корня, в основании, развивающихся в базальной части побега почек возобновления, формируются хорошо ветвящиеся придаточные корни. Среди боковых корней отчетливо выделяются 1-2 утолщенных, остальные нитевидные, сильно ветвящиеся. По мере нарастания вегетативной мощности растения увеличивается длина и число корневищ, это способствует расширению площади питания растения и закреплению его в субстрате. Общее число корневищ может достигать 8-10, их длина - 6-7 см. Для усиления питания особи на корневище формируются тонкие сосущие придаточные корни.
В средневозрастном генеративном возрастном состоянии (G2) надземная и подземная сферы T. pacificum наиболее развиты. Согласно И.Г. Серебрякову [13] жизненную формы (ЖФ) особи определяется именно в этом возрастном состоянии. ЖФ клевера тихоокеанского, произрастающего на заповедной территории, в указанных в начале статьи экологических условиях, можно описать как длиннокорневищно-вторично-стержнекорневой травянистый поли-карпик с удлиненным стелющимся моноцикли-ческим монокарпическим побегом. За счет ежегодно трогающихся в рост почек возобновления в надземной сфере формируются генеративные и вегетативные побеги. Генеративные годичные побеги состоят из 12-20 метамеров, длиной до 41 см. Вегетативные побеги короче, число метамеров у них значительно меньше и составляет 58 (10). Побеги ветвятся в зоне обогащения [5]. Морфология листьев соответствует описанной в состоянии G1. В состоянии G2 размеры особи максимальны. Соцветия верхушечные и пазушные, головчатые; цветки в соцветии малиновые. Диаметр соцветия и длина его оси также достигают максимума.
Особи клевера тихоокеанского в этом возрастном состоянии представляет собой куртину, достигающую 20 см в диаметре, и может состоять из одного материнского и 9-12 парциальных кустов. Кроме этого подземная сфера представлена системой главного корня, корневищами и веретеновидно утолщенными придаточными корнями. Последние формируются как в основании почек расположенных на корневище, так и в основании почек возобновления, заложенных на главах парциальных кустов. Эти корни своим обликом напоминают главный корень. Длина их достигает 26-30 см, диаметр в верхней трети до 5 мм. В конце средневозрастного генеративного состояния начинается разрушение материнского каудекса в его средней части, в более старом участке, и главного корня.
Старых генеративных особей T. pacificum мы не обнаружили. Вероятно, в это время продолжается разрушение особи и распада ее на парциали. К сенильному периоду относятся особи, которые утратили способность к образованию генеративных побегов. В этом периоде мы выделяем два возрастных состояния: субсенильное и сенильное.
Особи субсенильного возрастного состояния (ss) это парциали, которые представляют собой корневищные вегетативные особи. В надземной сфере формируются простые, единичные побеги с неполным циклом развития. Число метамеров на побеге 2-5, общая длина побега достигает 8 см. В основании побега иногда закладываются 1 (2) слабо развитых почки возобновления. Развивающиеся листья на побеге тройчатосложные, их размеры соответствуют таким же в имматурном возрастном состоянии. Подземная сфера представлена только остатками корневища. Оно полуполое, продолжающее разрушаться в местах ветвления. Сенильные растения (s) T. pacificum найдены не были.
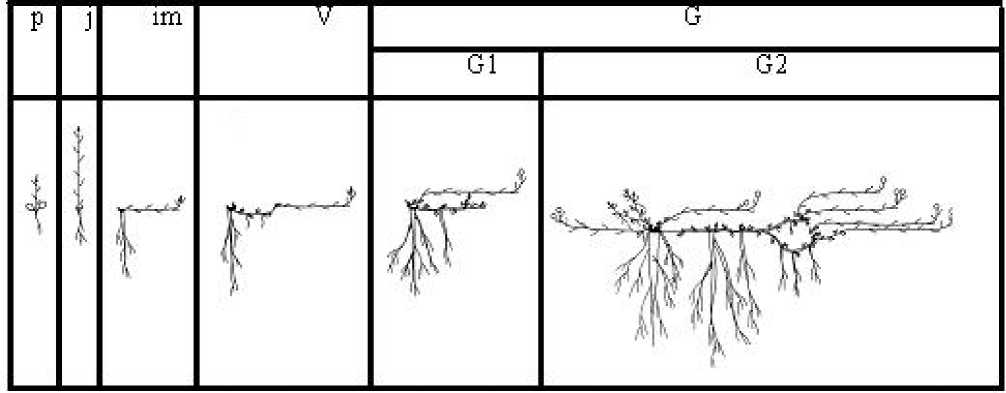
Рис. 1. Схема онтоморфогенеза T. pacificum на территории Лазовского заповедника
Выводы: нами был изучен онтоморфогенез клевера тихоокеанского на территории Лазовского государственного заповедника. Особенностью развития этого вида в заповеднике в зоне супралиторали является раннее формирование корневища (уже при переходе особи из имматурного в вегетативное возрастное состояние) и веретеновидно утолщенных придаточных корней, которые своим обликом напоминают главный корень. Продолжительность большого жизненного цикла длинноконевищно-вторично-стержнекорневой ЖФ T. pacificum точно мы определить не можем в связи с ранним формированием корневища и плохо выраженными годичными приростами.
Список литературы Онтоморфогенез клевера тихоокеаского на территории Лазовского государственного заповедника им. Л. Г. Капланова
- Безделева, Т.А. Биоморфологические особенности прибрежно-морских растений/Т.А. Безделева, Е.А. Пименова//Материалы конф., посвященной 50-летию Ботанического сада-института ДВО РАН. -Владивосток: Дальнаука, 1998. -С. 91-93.
- Бобров, Е.Г. О Люпиновидном клевере Trifolium lupinaster L.//Президенту АН СССР акад. В. Л. Комарову. -Л.: АН СССР, 1939. -С. 130-142.
- Ворошилов, В.Н. Флора советского Дальнего Востока. -М.: Наука, 1966. -С. 271.
- Дудик, Н.М. Морфология плодов бобоцветных в связи с эволюцией. -Киев: Наукова думка, 1979. -212 с.
- Михайлова, Т.Д. Биоморфологические особенности Astragalus glycyphylly L.//Бюлл. МОИП. Отд. биол. -1970. -Т. LXXV, вып. 5. -С. 74-81.
- Определитель растений Приморья и Приамурья. -М.-Л.: Наука, 1966. -С. 238.
- Определитель высших растений Сахалина и Курильских островов. -М.: Наука, 1974. -С. 215.
- Павлова, Н.С. Сем. Бобовые -Fabaceae Lindl s. l.//Сосудистые растения советского Дальнего Востока. -М.: Наука, 1989. -Т. 4. -С. 191-339.
- Работнов, Т.А. Длительность виргинильного периода жизни травянистых многолетников естественных ценозах//Бюлл. МОИП. Отд. биол. -1946. -Т. 51, вып. 2. -С. 917.
- Работнов, Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах//Тр. БИН АН СССР. Сер. 3. Геоботаника. -М.-Л., 1950. -Вып. 6. -С. 7-204.
- Работнов, Т.А. Методы определения возраста и длительности жизни у травянистых растений//Полевая геоботаника. -М.-Л.: АН СССР, 1960. -Т. 2. -С. 141-149.
- Росков, Ю.Р. Ревизия рода Trifolium L. s. l. во флоре СССР: Дисс… канд. биол. наук. -Л., 1990. -261 с.
- Серебряков, И.Г. Экологическая морфология растений. -М.: Высшая школа, 1962. -378 с.
- Серебряков, И.Г. Жизненные формы высших растений и их изучение//Полевая геоботаника. -Л.: Наука, 1964. -Т. 3. -С. 146-205.
- Серебрякова, Т.И. Учение о жизненных формах растений на современном этапе. -М.: ВИНИТИ, 1972. -Т. 1. -С. 84-169.
- Уранов, А.А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов//Биол. науки. -1975. -№2. -С. 7-34.
