Описание форм птичьих яиц с помощью геометрических эталонов

Автор: Митяй Иван Сергеевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Зоология
Статья в выпуске: 1, 2014 года.
Бесплатный доступ
Предложена оригинальная методика описания птичьих яиц на основании геометрии асимметрического овала (овоида), имеющего параметры: D=1,0; L=2-(√(2/2); r i=0,5; r l=2,0; r c=1-(√(2/2). Использование свойств этой фигуры позволило создать систему из 80 базовых овоидов-эталонов, имеющих соответствующие названия, цифровую и буквенную кодировки. Составление одинаковых базовых овоидов инфундибулярной зоной (тупой полюс) выявило 80 эталонов симметрических псевдоовоидов, из которых у птиц встречается 44. Такая же процедура с разными овоидами дала 375 эталонов асимметрических псевдоовоидов, объединенных нами в шесть групп. Применение упомянутой системы эталонов позволило определять реальные формы птичьих яиц, анализировать связь их морфометрических параметров с инкубационными свойствами, а также осуществлять сравнения и обобщения данных других авторов. Каждый из эталонов сопровождается количественными характеристиками в виде интерполярного индекса и индексов форм: инфундибулярной, клоакальной, латеральной зон, асимметрии, удлиненности и комплементарности.
Овоид, псевдоовоид, индексы формы яйца, классификация форм птичьих яиц
Короткий адрес: https://sciup.org/146116486
IDR: 146116486 | УДК: 59.087:
Description of avian eggs’ shapes using geometrical standards
The original technique of description of avian eggs on the basis of the geometry of asymmetrical oval (ovoid) - a figure with parameters: D=1,0; L=2-(√(2/2); ri=0,5; rl =2,0;rc= 1-(√(2/2) - is described. Specific properties of this figure allow creating a system of 80 basic ovoid standards, each given an appropriate name, digital and letter coding, and distinct quantitative characteristics. Combining infundibular zones (blunt poles) of basic ovoids in pairs gives 80 standards of symmetric pseudo-ovoids, 44 of which are found in birds. The same procedure applied to different ovoids produces 375 standards of asymmetrical pseudo-ovoids. They can be divided into six groups. Use of such system of standards enables us to identify real shapes of avian eggs, to analyze relation of morphometric parameters to incubatory properties of eggs, and also to carry out comparisons and generalizations of other authors’ data. Each standard is quantitatively characterized by means of indexes of infundibular, cloacal and lateral zones, by the index of asymmetry, elongation index, complementarity index as well as interpolar index.
Текст научной статьи Описание форм птичьих яиц с помощью геометрических эталонов
Введение. Птичье яйцо как биологическое тело с геометрической точки зрения устроено довольно просто. Однако специфика форм яиц у разных видов птиц порождает определенные трудности в процессе их описания. В полевых условиях непосредственно с яйца можно снять легко только два промера: диаметр и длину. Их, безусловно, недостаточно для полноценного описания формы. Дополнительные параметры можно получить или из плоскостной проекции (светокопии, фотографии) (Romanoff, Romanoff, 1949; Костин, 1977; Мянд, 1988; Fuhrer-Nagy, 2002), или же осуществить замеры при помощи специально изготовленных устройств (Preston, 1953). С появлением цифровой фотографии и возможностей их компьютерной обработки проблема снятия дополнительных параметров стала легко решаемой. В литературе появилось немало работ, посвященных этим методикам описания яиц (Makatsch, 1976; Anderson, 1978; Todd, Smart, 1984; Barta, Szekely, 1997; Baker, 2002; Митяй, 2003, 2008; Monus, Barta, 2005; Narushin, 2005; Bridge et al., 2007). Упомянутые методики все же не
Вестник ТвГУ. Серия "Биология и экология". 2014. № 1 разрешили проблем, связанных с названием, классификацией форм яиц, а также возможностью сравнения ооморфометрических данных, публикуемых различными авторами.
В литературе нет единого подхода в отношении названий форм яиц. Они или малоинформативны, типа «яйцо с ярко выраженным тупым и острым концом», или тавтологичны - «овоидный тип формы яйца». Не лишены недостатков и названия по подобию с геометрическими фигурами (сферические, эллипсоидные, овальные) или другими телами (каплевидные, грушевидные, юловидные). Зачастую это подобие очень условно, поскольку в реальных яйцах на разных полюсах проявляются разные фигуры. Не существует также единства в выборе количества базовых фигур. У разных авторов их: три (Makatsch, 1976; Климов, 1993), четыре (Preston, 1953; Narushin, 2005), пять (Gotman, Jablonski, 1972), восемь (Walters, 1994), десять (Schonwetter, 1985; Barta, Szekely, 1997). К тому же, предлагаемые фигуры рассматриваются вне единства и количественных характеристик.
В связи с этим, обобщение и сравнение данных разных авторов в значительной степени затруднено. На наш взгляд, проблема может быть решена при наличии комплексной системы описания форм яиц, включающей в себя название, геометрическую модель (фигуру) и ее количественные параметры. Другими словами, необходима общепринятая методика в виде эталонов форм, используя которые исследователи работали бы в едином русле. Обоснованию и изложению подобной методики посвящена данная статья.
Методика. Материалом для исследования послужили 16494 яиц 800 видов птиц Северо-Западной Палеарктики, относящихся к 20 отрядам.
В качестве базовой модели автором использована фигура, которая в начертательной геометрии и инженерной графике называется овоидом или асимметрическим овалом (рис. 1).
Согласно одному из определений, овоид является плоской, замкнутой, выпуклой, гладкой кривой, состоящей из взаимно сопрягающихся дуг окружностей разных радиусов. Характерными признаками этой кривой является наличие одной оси симметрии и не менее четырех вершин. В реальных яйцах этим вершинам соответствуют зоны: инфундибулярная (место размещения воздушной камеры), клоакальная (противоположная, более заостренная - место локализации аллантоиса) и две латеральные, являющиеся боками интерполярной зоны, в которой размещается зародыш. На плоскостной проекции каждая из этих зон очерчена одноименными дугами, сопряжение которых и образует овоид.
Для описания и классификации форм яиц автор использовал модель составного овоида, согласно которой все разнообразие форм получается путем составления (комбинации, сопряжения, плавного перехода) дуг адекватных кривизне зон овоида. Для каждой из форм строилась геометрическая фигура, наглядно отражающая отношения клоакальной, инфундибулярной дуг, длины и диаметра овоида. Количественное описание овоидов осуществляли с помощью семи индексов формы: традиционный индекс удлиненности - Iei= L/D и шесть индексов, предложенных нами: (112=ц/В; Iiz=ri/D; Icz=rc/D; Iipz=L-(й+Гс) lel^Hz+Icz), las Iiz~Icz O"i-l*c)/D, Icom (гс+1ф2)(1|р2+Г})/1{р2К, ГДС. ф2, Icz, 1ц, las, Icom, Lpz - индексы инфундибулярной, клоакальной, латеральной зон, асимметрии, интерполярный и индекс комплементарности; р, гС) р -радиусы дуг; L -длина; D - диаметр. Последний индекс отражает степень сбалансированности (гармоничности) инфундибулярного (р) и клоакального (гс) радиусов с длиной овоида. В конформной геометрии этот показатель называется двойным отношением или вурфом. Для описания форм яиц этот индекс нами предлагается впервые. Значения, приближающиеся к единице, характеризуют яйца с минимальным радиусом клоакальной зоны. Величины, приближающиеся к 1,333, свидетельствуют, что длина яйца состоит из трех равных отрезков, два из которых являются радиусами полярных зон. Показатели, уходящие за пределы 1,333, характеризуют яйца с большими радиусами клоакальных дуг.
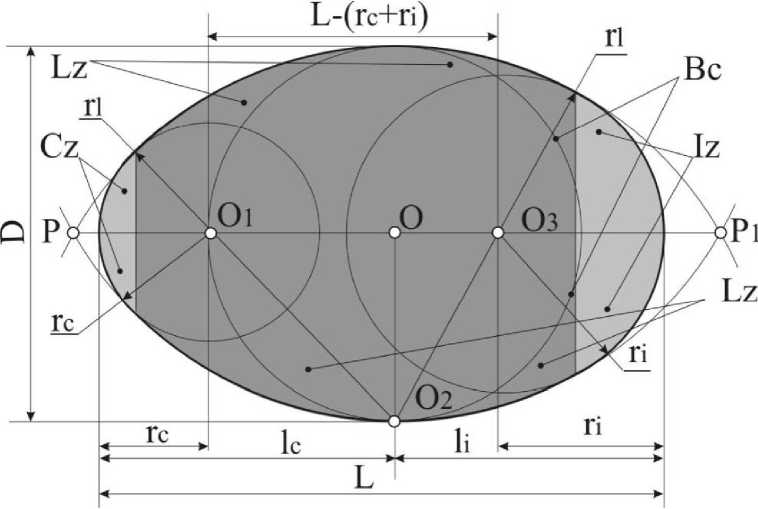
Рис. 1 . Обобщенная схема овоида и варианты снятия параметров: 0-0з - центры сопрягаемых дуг; Р, Pi - точки пересечения латеральных дуг; Вс - базовая окружность; Iz, Lz, Cz - инфундибулярная, латеральная и клоакальная зоны овоида и их радиусы: г,, ц, rc; D - диаметр; L - длина;
Н, 1с - инфундибулярная и клоакальная части длины
Исходные параметры, длина (L) и диаметр (D), реальных яиц измеряли штангенциркулем с точностью до 0,1 мм. Замеры необходимых радиусов дуг окружностей выполняли по цифровым
Вестник ТвГУ. Серия "Биология и экология". 2014. № 1
фотографиям с помощью компьютерных программ, разработанных Б. Троценко и С. Шелестюком. Им автор выражает искреннюю благодарность.
Результаты и обсуждение. Яйца большинства птиц, на первый взгляд, кажутся весьма однообразными. Различия становятся явными при сравнении параметров их форм между собой. Простое сравнение плоскостных проекций яиц показывает, что у одних полярные зоны одинаковы, а у других разные. На основании этого все разнообразие форм яиц можно разделить на две группы: симметрические и асимметрические (рис. 2).
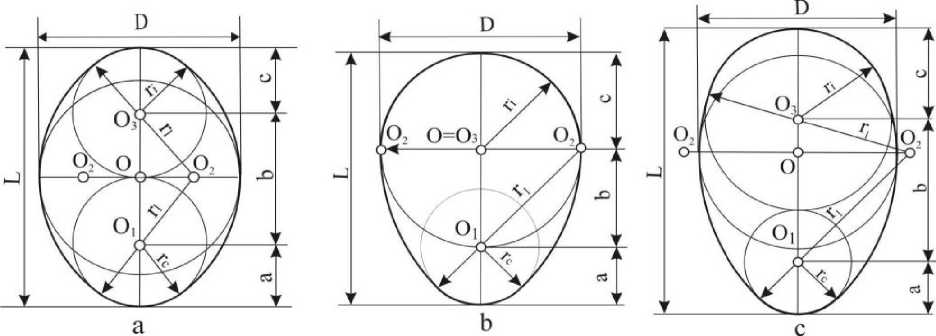
Рис. 2. Формы яиц по конфигурации полярных зон: а) симметрические; Ь, с) асимметрические
Для классификации форм яиц в качестве исходной (базовой) геометрической фигуры подходит овоид, в котором одна часть является половиной окружности, а другая - половиной овала (рис. 2, Ь). Исходя из этого, все фигуры, инфундибулярный радиус которых лежит в пределах половины базового диаметра (ц=0,5-0,499 D), получают название овоидов. Они составляют 21,3% от указанного выше количества яиц (п=16490). Учитывая то, что инфундибулярный радиус константный, дальнейшее разграничение форм осуществляется по отличиям латеральных и клоакальных радиусов. Все разнообразие форм возникает в пространстве, ограниченном двумя латеральными дугами, которые пересекаются в точках Р и Pi (рис. 3).
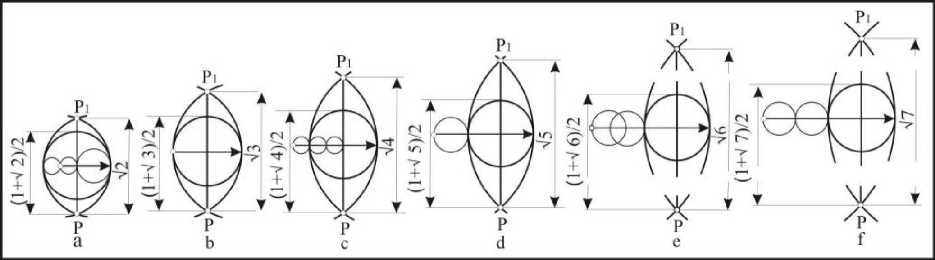
Рис. 3 . Интервалы и взаимосвязь латеральних дуг с диаметром и длиной овоидов
Взаимосвязь радиусов латеральных дуг и расстояний между точками их пересечения выражается следующим уравнением: PPi= V(4Dri - D2). В соответствии с ним, радиусам латеральных дуг 0,75D; 1,OD; 1,25D; 1,5D; 1,75D; 2,0D отвечают расстояния, равные корням квадратным из чисел 2, 3, 4, 5, б и 7. В каждом из упомянутых случаев радиусы сопрягающих дуг разные, что отражается на конфигурации клоакальной зоны. Сопряжение латеральных дуг осуществляется клоакальной окружностью таким образом, что его противоположная от места сопряжения часть может занимать различное положение на продольной оси профиля в зависимости от величины диаметра (рис. 4).
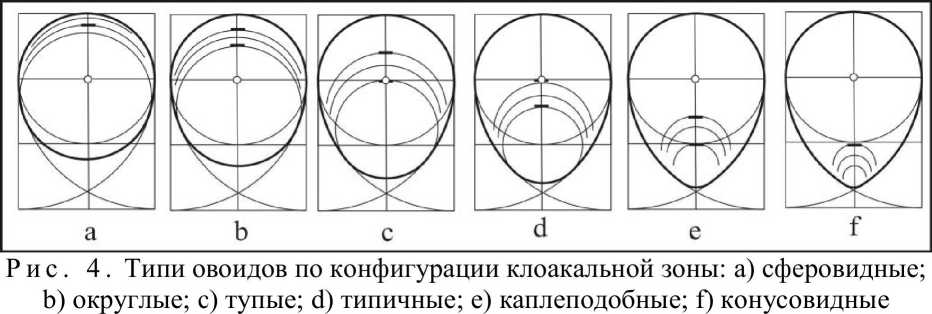
Если продольную ось условно разбить на интервалы, то мы получим возможность количественно выражать радиусы клоакальных дуг через половины отрезков длины овоида: L-0,125; L-0,25; L-0,5; L-0,75; L-D; (L-D)/2; (L-D)/4. Исходя из этого, было выделено шесть типов овоидов, названия которым были даны в соответствии с положением клоакальной окружности: сферовидные, округлые, тупые, типичные, каплевидные и конусовидные. Каждая из этих форм внутри себя имеет различные радиусы клоакальной дуги. Поэтому вышеупомянутые овоиды получили дополнительные названия: крупнорадиусные, среднерадиусные и мелкорадиусные. В зависимости от того, в пределах каких латеральных дуг происходит сопряжение, длина овоидов бывает различной. Это дало еще пять дополнительных названий: короткие, укороченные, нормальные, удлиненные и длинные. Приняв во внимание упомянутые геометрические особенности овоида, мы разработали систему из 80 базовых овоидов, объединенных в 6 типов.
Первый тип включает формы, приближающиеся к сфере. Они получили название сфероподобные. Их клоакальные окружности имеют диаметры в пределах D>dc>L-0,125D (рис. 5).
Индекс удлиненности таких яиц приближается к единице: 1,0<1е1<1,09. У птиц таких форм очень мало, около 0,2% (п=3498). Они встречаются отрядах Galliformes, Piciformes и Passeriformes.
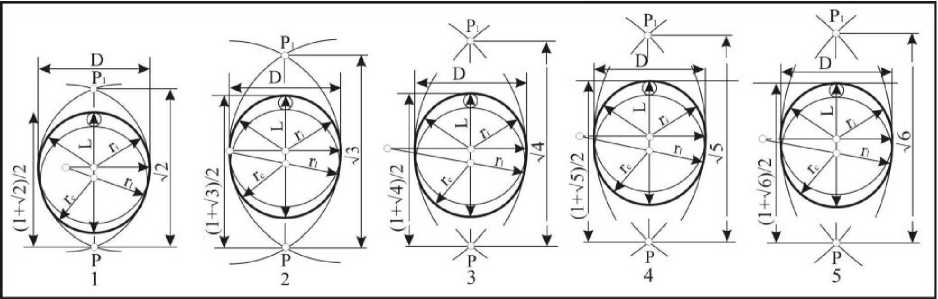
Рис. 5 . Сфероподобные овоиды
Второй тип объединяет формы с диаметрами в пределах L-0,125D>de>L-0,375D (рис. 6). Они получили условное название округлые: крупнорадиусные (6-10), среднерадиусные (11-15) и мелкорадиусные (16-20) с дополнительными характеристиками: короткие (6, 11, 16), укороченные (7, 12, 17), нормальные (8, 13, 18), удлиненные (9, 14, 19) длинные (10, 15, 20). Индекс удлиненности: 1,091<1е1< 1,287. Встречаются указанные формы в 4,0% случаев представителей отрядов, приведенных для первого типа (рис. 6).
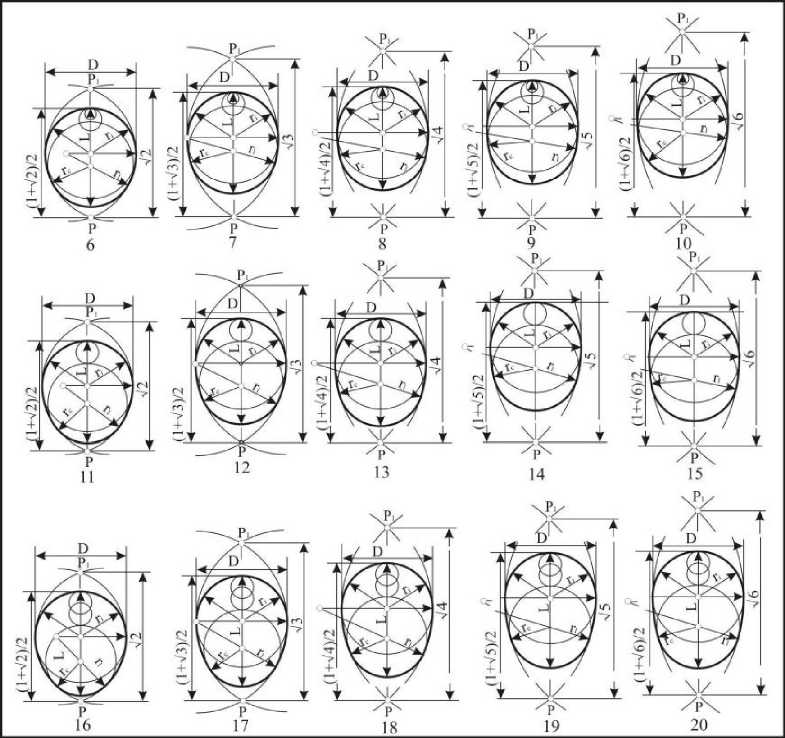
Рис. 6. Округлые овоиды
Третий тип, объединяющий формы с диаметрами в пределах L-0,375D>dc>L-0,625D (рис. 7), представлен формами, названными нами тупыми овоидами. Каждая из этих форм, как и в предыдущем случае, делится на крупно-, средне- и мелкорадиусные (21-25, 26-31, 32-36), и, соответственно, короткие (21, 26, 32), укороченные (22, 27, 33), нормальные (23, 28, 34), удлиненные (24, 30, 35) и длинные (25, 31, 36). Индекс удлиненности яиц третьего типа находится в пределах 1,146<1е1<1,4. Упомянутые формы встречаются в 40,9% случаев у представителей из отрядов Galliformes, Charadriiformes, Falconifonnes и Piciformes Passeriformes. Незначительное их число отмечено у Gruiformes и Coraciiformes.
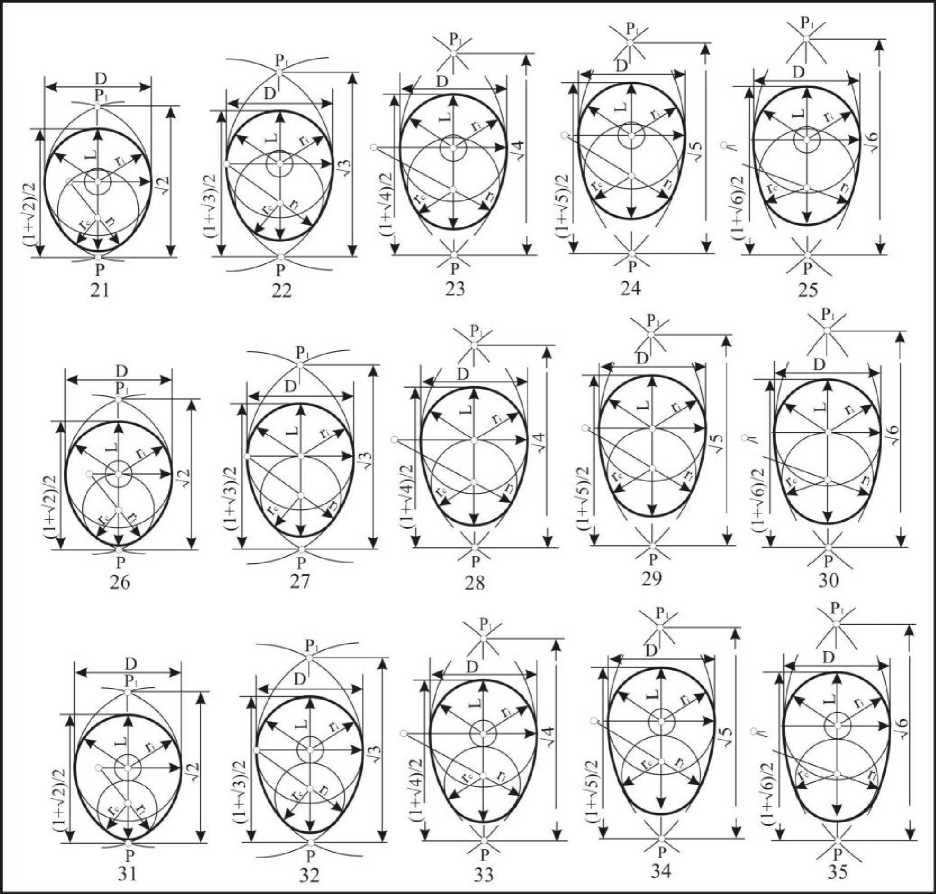
Рис. 7 . Тупые овоиды
Четвертый тип включает в себя типичные овоиды - формы с клоакальными окружностями в пределах L-0,625D>de>L-0,75D (рис. 8). Они подразделяются на: крупнорадиусные (36-40), среднерадиусные (41-45) и мелкорадиусные (46-50). По длине подразделение
Вестник ТвГУ. Серия "Биология и экология". 2014. № 1 осуществляется на: короткие (36, 41, 46), укороченные (37, 42, 47), нормальные (38, 43, 48), удлиненные (39, 44, 49) и длинные (40, 45, 50). Индекс удлиненности тупых овоидов находится в пределах 1,146<1е1<1,4. Упомянутые формы встречаются в 41,2% случаев у представителей отрядов Anseriformes, Charadriiformes, Falconiformes, Galliformes, Gruiformes, Picifomies и Passeriformes.
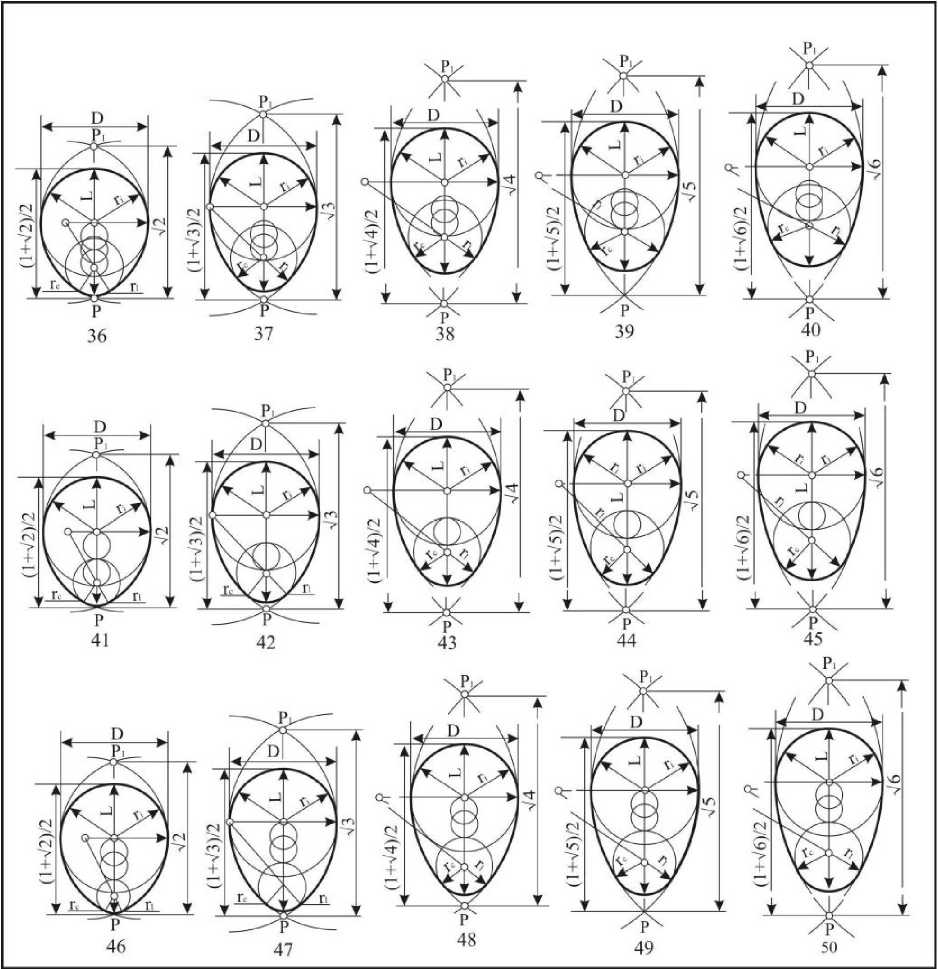
Рис. 8 . Типичные овоиды
Пятый тип: клоакальные радиусы находятся в пределах L-0,75D>dc>L-l,125D (рис. 9). Эти формы получили название каплевидных овоидов. Они подразделяются на: крупнорадиусные (5155), среднерадиусные (56-60) и мелкорадиусные (61-65). По длине подразделение осуществляется на: короткие (51, 56, 61), укороченные (52, 57, 62), нормальные (53, 58, 63), удлиненные (54, 59, 64) и длинные (55, 60, 65). Индекс удлиненности этих яиц находится в
Вестник ТвГУ. Серия "Биология и экология". 2014. № 1
пред елах,323<1е1< 1,643. Каплевидные овоиды встречаются в 48,2% случаев у представителей двух отрядов, Charadriiformes и Passerifonnes.
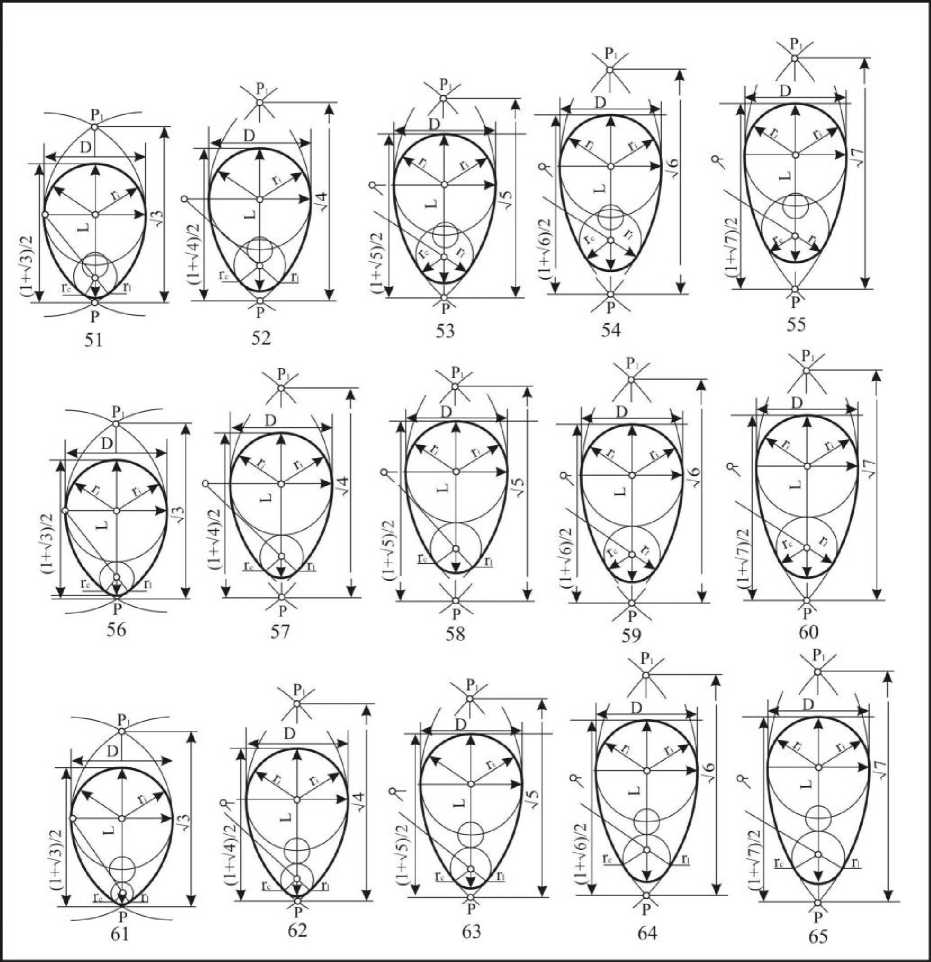
Рис. 9 . Каплевидные овоиды
Шестой тип : клоакальные окружности находятся L-0,125D>dc>L-2,0D (рис. 10). Они получили название конусовидных овоидов: крупнорадиусных (66-70), среднерадиусных (71-75) и мелкорадиусных (76-80). По длине овоиды шестрого типа подразделяются на: короткие (66, 71, 76), укороченные (67, 72, 77) нормальные (68, 73, 78), удлиненные (69, 74, 79) и длинные (70, 75, 80). Индекс удлиненности этих яиц находится в пределах 1,449<1е1<1,745. Конусовидные овоиды встречаются в 9,3% случаев у представителей отряда Charadriiformes.
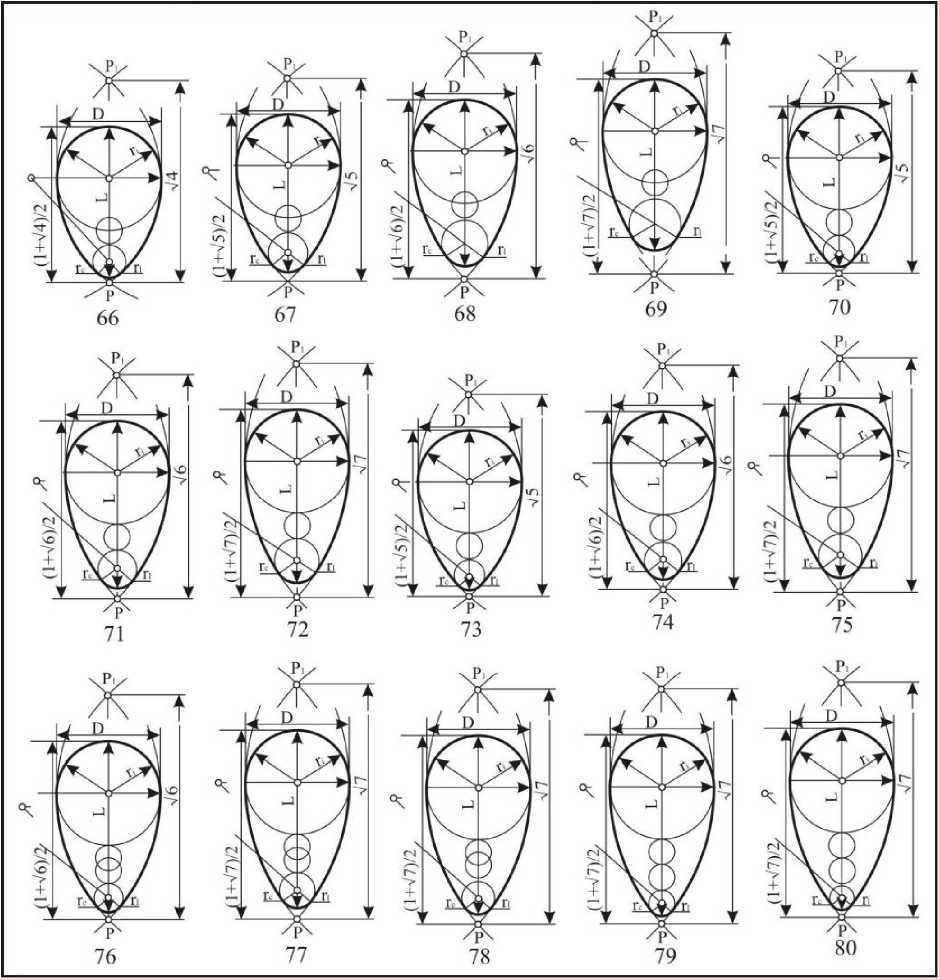
Рис. 10. Конусовидные овоиды
Вышеприведенные геометрические овоиды-эталоны имеют фиксированные (индивидуальные) характеристики, выраженные в виде индесов клоакальной и латеральной зон, а также индексов удлинености, комплементарности, асиметрии и интерполярности. Получившие порядковые номера, названия и характеристики, указанные эталоны предлагаются нами в качестве базовых при описании формы яйца (табл. 1).
Таблица 1
Названия и параметры базовых овоидов-эталонов
|
№ |
Название эталона |
Icz |
Ilz |
lei |
Icom |
las |
lipi |
|
Сферовидные овоиды: |
|||||||
|
1 |
короткие |
0,480 |
0,75 |
1,083 |
3,152 |
0,020 |
0,103 |
|
2 |
укороченные |
0,487 |
1,00 |
1,098 |
2,987 |
0,013 |
0,112 |
|
3 |
нормальные |
0,490 |
1,25 |
1,111 |
2,822 |
0,010 |
0,121 |
|
4 |
удлиненные |
0,493 |
1,50 |
1,115 |
2,803 |
0,007 |
0,123 |
|
5 |
длинные |
0,494 |
1,75 |
1,116 |
2,814 |
0,006 |
0,122 |
|
Округлые овоиды |
|||||||
|
6 |
Крупнорадиусные: короткие |
0,478 |
0,75 |
1,091 |
2,806 |
0,022 |
0,120 |
|
7 |
укороченные |
0,488 |
1,00 |
1,108 |
2,826 |
0,012 |
0,121 |
|
8 |
нормальные |
0,490 |
1,25 |
1,112 |
2,797 |
0,010 |
0,123 |
|
9 |
удлиненные |
0,491 |
1,50 |
1,140 |
2,439 |
0,009 |
0,150 |
|
10 |
длинные |
0,493 |
1,75 |
1,143 |
2,432 |
0,007 |
0,151 |
|
11 |
Среднерадиусные: короткие |
0,437 |
0,75 |
1,125 |
2,029 |
0,063 |
0,189 |
|
12 |
укороченные |
0,459 |
1,00 |
1,169 |
1,935 |
0,041 |
0,210 |
|
13 |
нормальные |
0,468 |
1,25 |
1,190 |
1,883 |
0,032 |
0,223 |
|
14 |
удлиненные |
0,473 |
1,50 |
1,200 |
1,868 |
0,027 |
0,227 |
|
15 |
длинные |
0,477 |
1,75 |
1,207 |
1,856 |
0,023 |
0,231 |
|
16 |
Мелкорадиусные: короткие |
0,387 |
0,75 |
1,151 |
1,637 |
0,113 |
0,264 |
|
17 |
укороченные |
0,423 |
1,00 |
1,218 |
1,589 |
0,077 |
0,295 |
|
18 |
нормальные |
0,441 |
1,25 |
1,256 |
1,556 |
0,059 |
0,316 |
|
19 |
удлиненные |
0,449 |
1,50 |
1,271 |
1,549 |
0,051 |
0,322 |
|
20 |
длинные |
0,456 |
1,75 |
1,287 |
1,535 |
0,044 |
0,331 |
|
Тупые овоиды |
|||||||
|
21 |
Крупнорадиусные: короткие |
0,382 |
0,75 |
1,146 |
1,629 |
0,118 |
0,265 |
|
22 |
укороченные |
0,416 |
1,00 |
1,218 |
1,565 |
0,084 |
0,302 |
|
23 |
нормальные |
0,433 |
1,25 |
1,266 |
1,512 |
0,067 |
0,334 |
|
24 |
удлиненные |
0,444 |
1,50 |
1,284 |
1,507 |
0,056 |
0,341 |
|
25 |
длинные |
0,452 |
1,75 |
1,301 |
1,498 |
0,048 |
0,349 |
|
26 |
Среднерадиусные: короткие |
0,335 |
0,75 |
1,167 |
1,432 |
0,165 |
0,332 |
|
27 |
укороченные |
0,375 |
1,00 |
1,250 |
1,400 |
0,125 |
0,375 |
|
28 |
нормальные |
0,403 |
1,25 |
1,306 |
1,383 |
0,097 |
0,403 |
|
29 |
удлиненные |
0,417 |
1,50 |
1,333 |
1,375 |
0,083 |
0,417 |
|
30 |
длинные |
0,430 |
1,75 |
1,359 |
1,368 |
0,070 |
0,430 |
|
№ |
Название эталона |
Icz |
Ilz |
lei |
Icom |
las |
lipi |
|
31 |
Мелкорадиусные: короткие |
0,285 |
0,75 |
1,177 |
1,309 |
0,215 |
0,392 |
Продолжение табл. 1
|
№ |
Название эталона |
Icz |
Ilz |
lei |
Icom |
las |
lipi |
|
32 |
укороченные |
0,342 |
1,00 |
1,275 |
1,310 |
0,158 |
0,433 |
|
33 |
нормальные |
0,371 |
1,25 |
1,334 |
1,300 |
0,129 |
0,463 |
|
34 |
удлиненные |
0,389 |
1,50 |
1,379 |
1,288 |
0,111 |
0,490 |
|
35 |
длинные |
0,408 |
1,75 |
1,400 |
1,296 |
0,092 |
0,492 |
|
Типичные овоиды |
|||||||
|
36 |
Крупнорадиусные: короткие |
0,269 |
0,75 |
1,169 |
1,287 |
0,231 |
0,401 |
|
37 |
укороченные |
0,309 |
1,00 |
1,289 |
1,250 |
0,191 |
0,480 |
|
38 |
нормальные |
0,342 |
1,25 |
1,342 |
1,255 |
0,158 |
0,500 |
|
39 |
удлиненные |
0,372 |
1,50 |
1,389 |
1,258 |
0,128 |
0,518 |
|
40 |
длинные |
0,383 |
1,75 |
1,427 |
1,247 |
0,117 |
0,544 |
|
41 |
Среднерадиусные: короткие |
0,217 |
0,75 |
1,183 |
1,196 |
0,283 |
0,467 |
|
42 |
укороченные |
0,276 |
1,00 |
1,299 |
1,203 |
0,224 |
0,524 |
|
43 |
нормальные |
0,312 |
1,25 |
1,374 |
1,202 |
0,188 |
0,562 |
|
44 |
удлиненные |
0,340 |
1,50 |
1,430 |
1,201 |
0,160 |
0,590 |
|
45 |
длинные |
0,356 |
1,75 |
1,146 |
1,534 |
0,144 |
0,606 |
|
46 |
Мелкорадиусные: короткие |
0,156 |
0,75 |
1,191 |
1,122 |
0,344 |
0,535 |
|
47 |
укороченные |
0,223 |
1,00 |
1,316 |
1,142 |
0,277 |
0,594 |
|
48 |
нормальные |
0,264 |
1,25 |
1,399 |
1,149 |
0,236 |
0,635 |
|
49 |
удлиненные |
0,307 |
1,50 |
1,451 |
1,164 |
0,193 |
0,644 |
|
50 |
длинные |
0,338 |
1,75 |
1,482 |
1,177 |
0,162 |
0,645 |
|
Каплевидные овоиды |
|||||||
|
51 |
Крупнорадиусные: короткие |
0,212 |
1,00 |
1,323 |
1,131 |
0,288 |
0,612 |
|
52 |
укороченные |
0,254 |
1,25 |
1,408 |
1,138 |
0,246 |
0,655 |
|
53 |
нормальные |
0,282 |
1,50 |
1,474 |
1,138 |
0,218 |
0,693 |
|
54 |
удлиненные |
0,305 |
1,75 |
1,532 |
1,137 |
0,195 |
0,728 |
|
55 |
длинные |
0,332 |
2,00 |
1,555 |
1,147 |
0,168 |
0,724 |
|
56 |
Среднерадиусные: короткие |
0,169 |
1,00 |
1,337 |
1,094 |
0,331 |
0,669 |
|
57 |
укороченные |
0,213 |
1,25 |
1,426 |
1,105 |
0,287 |
0,713 |
|
58 |
нормальные |
0,250 |
1,50 |
1,500 |
1,111 |
0,250 |
0,750 |
|
59 |
удлиненные |
0,277 |
1,75 |
1,553 |
1,115 |
0,223 |
0,777 |
|
60 |
длинные |
0,300 |
2,00 |
1,600 |
1,117 |
0,200 |
0,800 |
|
61 |
Мелкорадиусные: короткие |
0,122 |
1,00 |
1,345 |
1,063 |
0,378 |
0,723 |
|
62 |
укороченные |
0,175 |
1,25 |
1,445 |
1,079 |
0,325 |
0,770 |
|
63 |
нормальные |
0,212 |
1,50 |
1,516 |
1,087 |
0,288 |
0,804 |
|
64 |
удлиненные |
0,249 |
1,75 |
1,578 |
1,095 |
0,251 |
0,829 |
|
65 |
длинные |
0,259 |
2,00 |
1,643 |
1,089 |
0,241 |
0,884 |
|
Конусовидные овоиды |
|||||||
|
№ |
Название эталона |
Icz |
llz |
lei |
Icom |
las |
lipi |
|
66 |
Крупнорадиусные: короткие |
0,159 |
1,25 |
1,449 |
1,069 |
0,341 |
0,790 |
|
67 |
укороченные |
0,189 |
1,50 |
1,522 |
1,074 |
0,311 |
0,834 |
|
68 |
нормальные |
0,222 |
1,75 |
1,603 |
1,078 |
0,278 |
0,882 |
|
69 |
удлиненные |
0,249 |
2,00 |
1,643 |
1,085 |
0,251 |
0,894 |
|
70 |
длинные |
0,150 |
1,50 |
1,541 |
1,055 |
0,350 |
0,891 |
|
71 |
Среднерадиусные: короткие |
0,185 |
1,75 |
1,618 |
1,061 |
0,315 |
0,933 |
|
72 |
укороченные |
0,213 |
2,00 |
1,667 |
1,067 |
0,287 |
0,955 |
|
73 |
нормальные |
0,147 |
1,50 |
1,541 |
1,053 |
0,353 |
0,895 |
|
74 |
удлиненные |
0,180 |
1,75 |
1,621 |
1,059 |
0,320 |
0,942 |
|
75 |
длинные |
0,209 |
2,00 |
1,667 |
1,065 |
0,291 |
0,959 |
|
76 |
Мелкорадиусные: короткие |
0,139 |
1,75 |
1,649 |
1,042 |
0,361 |
1,011 |
|
77 |
укороченные |
0,171 |
2,00 |
1,705 |
1,048 |
0,329 |
1,035 |
|
78 |
нормальные |
0,157 |
1,75 |
1,720 |
1,043 |
0,343 |
1,063 |
|
79 |
удлиненные |
0,125 |
2,00 |
1,743 |
1,032 |
0,375 |
1,119 |
|
80 |
длинные |
0,116 |
2,00 |
1,745 |
1,029 |
0,384 |
1,129 |
Эти эталоны, показавшие свою эффективность к исследованным яйцам, можно использовать для построения эталонов остальных форм. Последние были получены путем совмещения (составления) базовых овоидов инфундибулярными зонами (тупыми полюсами) (рис. 11).
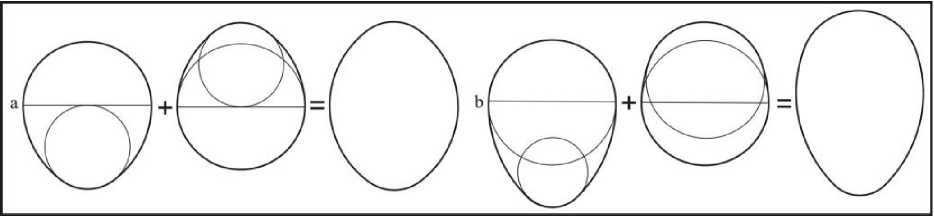
Рис. 11. Схемы составления симметрических (а) и асимметрических (Ь) псевдоовоидов
Получаемые при этом фигуры были названы нами «псевдоовоидами»; их инфундибулярный радиус меньше половины диаметра в отличие от такового у базовых овоидов. При совмещении между собой разных базовых овоидов получаются эталоны асимметрических псевдоовоидов. Комбинации одинаковых овоидов дают совокупность симметрических псевдоовоидов (рис. 12).
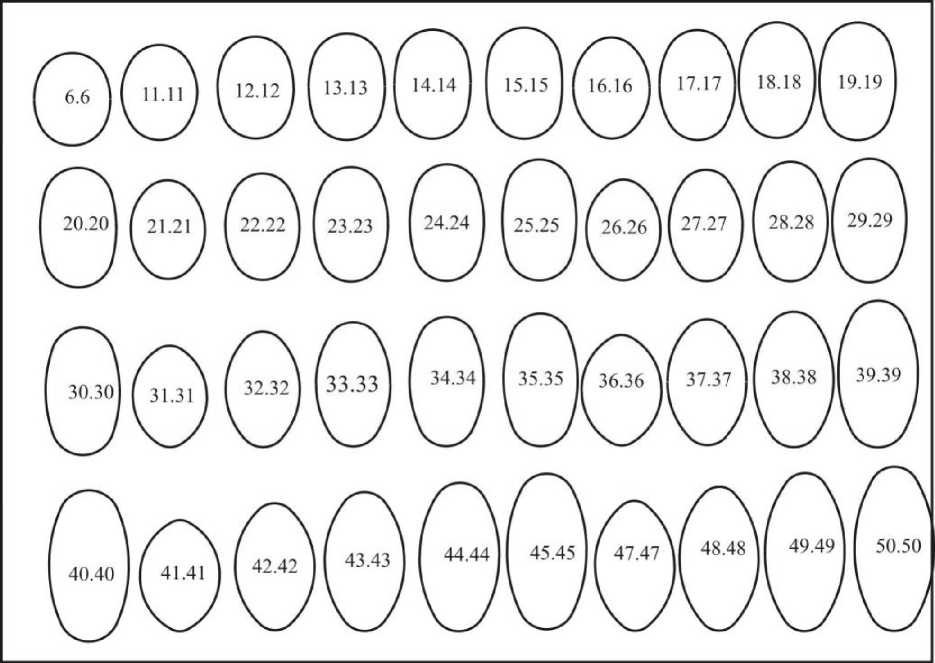
Рис. 1 2. Геометрические эталоны симметрических псевдоовоидов
При этом необходимо также учитывать то, что в природе яйца с абсолютно равными радиусами полярных зон практически отсутствуют. Поэтому к этой категории яиц мы предлагаем относить формы с индексом асимметрии не более 0,05. У птиц симметрические яйца встречаются гораздо реже (в пределах 5%, п=16490), чем у других животных, например, у рептилий. Это связано с множеством причин, среди которых наиболее существенными, на наш взгляд, являются три: а) некомпактность кладки; б) увеличенное раскатывание яиц; в) невозможность фиксированного направления зародышевого диска в сторону обогрева. Симметрические псевдоовоиды в незначительных количествах встречаются у представителей разных отрядов; они отсутствуют у Gaviiformes и Charadriiformes.
Совмещение 80 одинаковых эталонов базовых овоидов дает 80 теоретически возможных симметрических псевдоовоидов. У птиц таких форм меньше. У них отсутствуют крайние, т.е. приближающиеся к сфере и очень длинные и заостренные формы, хотя у других животных такие яйца являются нормой. Анализ нашей базы (п=16490) показал, что реальным формам яиц соответствуют 44 предлагаемых нами эталона.
Названия в данном случае даются, как и для базовых овоидов, с добавлением слов симметрический псевдоовоид. Цифровой код представляет собой двойной порядковый номер базового овоида, например, «тупой крупнорадиусный укороченный симметрический псевдоовоид (22.22)». Максимальное количество яиц относится к
Вестник ТвГУ. Серия "Биология и экология". 2014. № 1 асимметрическим псевдоовоидам (73,4%)- Их геометрические эталоны получаются путем совмещения разных базовых овоидов. Названия при таком подходе получаются очень длинными, т.к. включают в себя названия двух разных базовых овоидов. Поэтому для классификации яиц этого типа мы выбрали более простой способ.
Как отмечалось выше, у асимметрических псевдоовоидов радиусы дуг инфундибулярной зоны меньше 0,5D, но всегда больше, чем радиусы клоакальных дуг. Поэтому, сопрягая инфундибулярные дуги разного радиуса с 80 базовыми овоидами, мы получим совокупность эталонов асимметрических псевдоовоидов. Анализ нашей оологической базы данных показал, что радиусы инфундибулярных дуг рассматриваемого типа яиц колеблются в пределах от 0,285D до 0,491D. Всю эту совокупность мы условно разделили на шесть групп (рис. 13).
4500 -


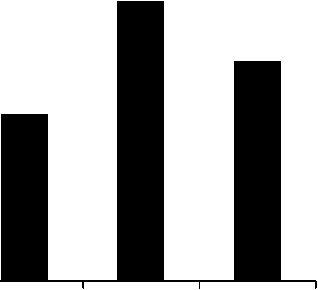
3000 .....
2500 -
2000 -
1000 -
500 -
0,400 0,418 0,437 0,455 0,473 0,491
Рис. 13. Распределение асимметрических псевдоовоидов по радиусам инфундибулярных дуг
Учитывая то, что формы, входящие в состав каждой из групп, отличаются только радиусами инфундибулярной дуги, их названия складываются из названия базового овоида и номера группы. Шифр состоит из комбинация цифр, отражающих номер базового овоида и номер группы, например: тупой крупнорадиусный укороченный псевдоовоид второй группы (22.2). Количество эталонов в каждой группе разное и уменьшается по мере совпадения радиусов инфундибулярной и клоакальной дуг.
Первая группа включает 75 эталонов, у которых инфундибулярный радиус находится пределах 0,491D>ri>0,474D. По форме эта группа наиболее близка к базовым овоидам (рис. 14). Среди асимметричных псевдоовоидов таких яиц 46,24% (п=12104). Чаще всего они встречаются в отрядах Charadriifomies и Passerifonnes. Их можно встретить также в отрядах Falconiformes, Gallifonnes, Gruifonnes и Piciformes. Индекс удлиненности таких яиц находится в пределах: 1,125<1е1<1,798.

Рис. 14. Асимметрические псевдоовоиды первой группы
Вторая группа (рис. 15) включает 68 эталонов с инфундибулярными радиусами 0,474D>ri>0,456D. Индекс удлиненности этих яиц находится в пределах 1,129<1е1< 1,803.
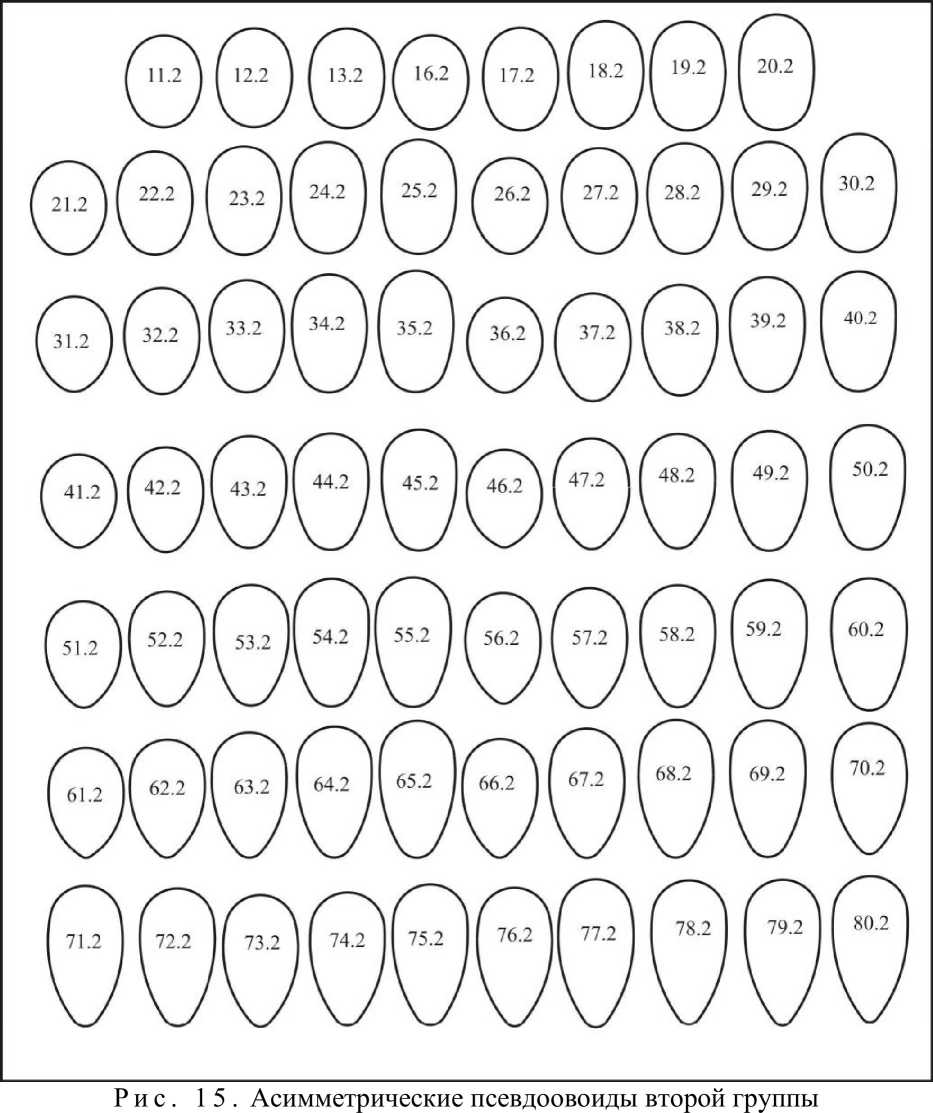
Среди асимметрических псевдоовоидов таких яиц 28,43%. Их больше всего в указанных выше отрядах. Появляются они у Anseriformes, Ciconiiformes, Strigiformes, Apodiformes, Cuculiformes, Coraciiformes и Upupiformes.
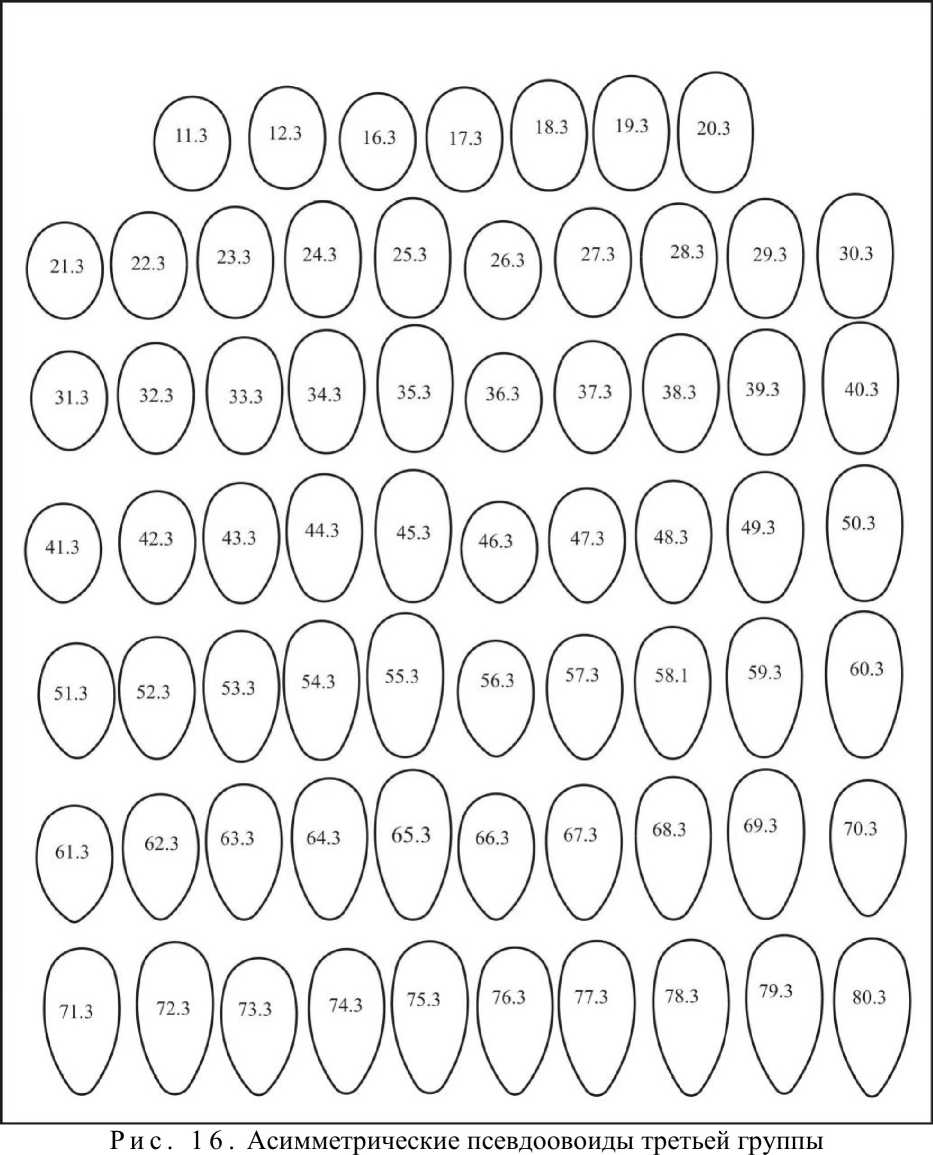
В третью группу (рис. 16) входят 67 эталонов, инфундибулярные радиусы которых находятся в пределах 0,456D>ri>0,437D. Индекс удлиненности этих яиц - 1,134<1е1<1,786.
Четвертая группа (рис. 17) представлена 60 эталонами, у которых инфундибулярный радиус находится в пределах 0,437D>ri>0,419D. Индекс удлиненности 1,150<1е1<1,806.
Указанные формы встречаются в 12,34% случаев. Максимальное их количество отмечается в отрядах Anseriformes, Falconiformes, Galliformes, Gruiformes, Strigiformes, Upupiformes, Piciformes и Passeriformes. Они встречаются также у Gaviiformes, Podicipediformes, Ciconiiformes, Coraciiformes, Columbiformes, Pelecaniformes, Procellariiformes, Caprimulgiformes, Cuculiformes, Apodiformes; их число уменьшается у Charadriiformes.

Рис. 17. Асимметрические псевдоовоиды четвертой группы
Таких форм насчитывается 7,54%. По отрядам наблюдается примерно такое же распределение, что и третьей группе.
В пятой группе насчитывается 55 эталонов с инфундибулярным радиусом в пределах 0,419D>ri>0,401D (рис. 18). Индекс удлиненности этих яиц находится в пределах 1,153<1е1<1,672. Таких форм насчитывается 3,44%. Максимальное их количество наблюдается в отрядах Anseriformes, Ciconiiformes Pelecaniformes, Podicipediformes и Gruiformes. В других отрядах (Struthionifomies, Falconiformes, Galliformes, Columbifonnes Strigiformes, Upupiformes, Picifonnes, Passeriformes) их количество сходно небольшое (рис. 18).
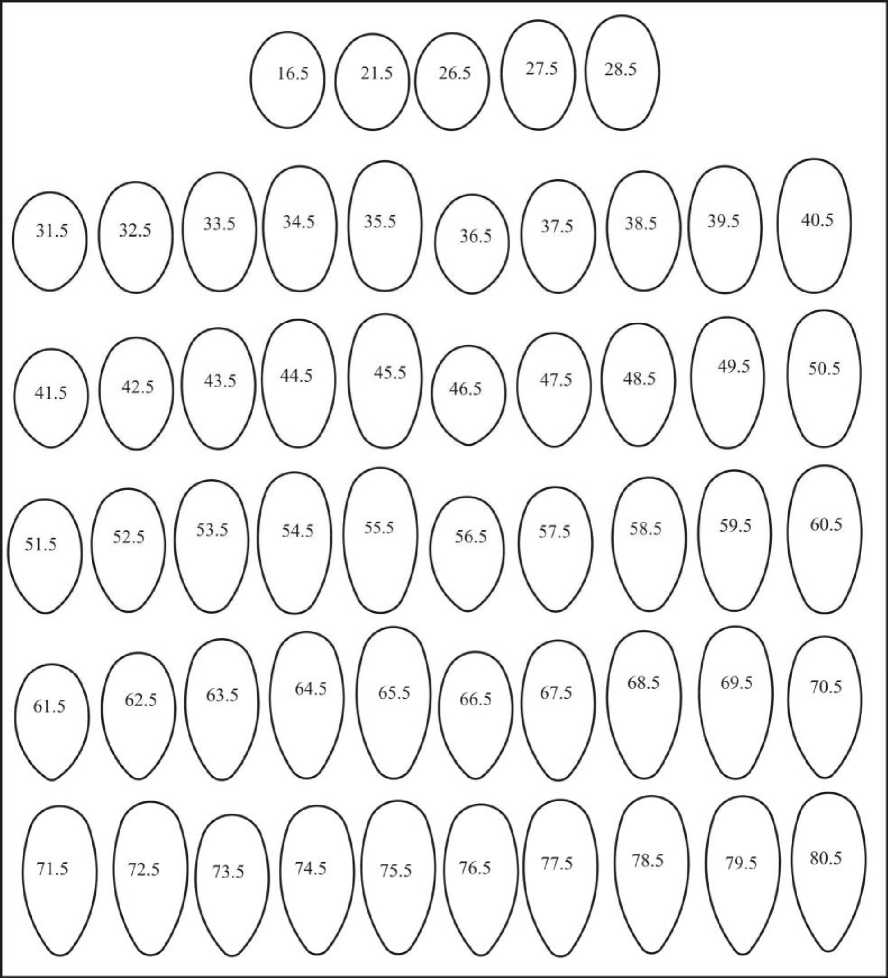
Рис. 18. Асимметрические псевдоовоиды пятой группы
Шестая группа (рис. 19) представлена 50 эталонами с инфундибулярным радиусом 0,409D>ri>0,285D. Индекс удлиненности 1,161<1е1< 2,0. Доля этих форм составляет 2,0%.
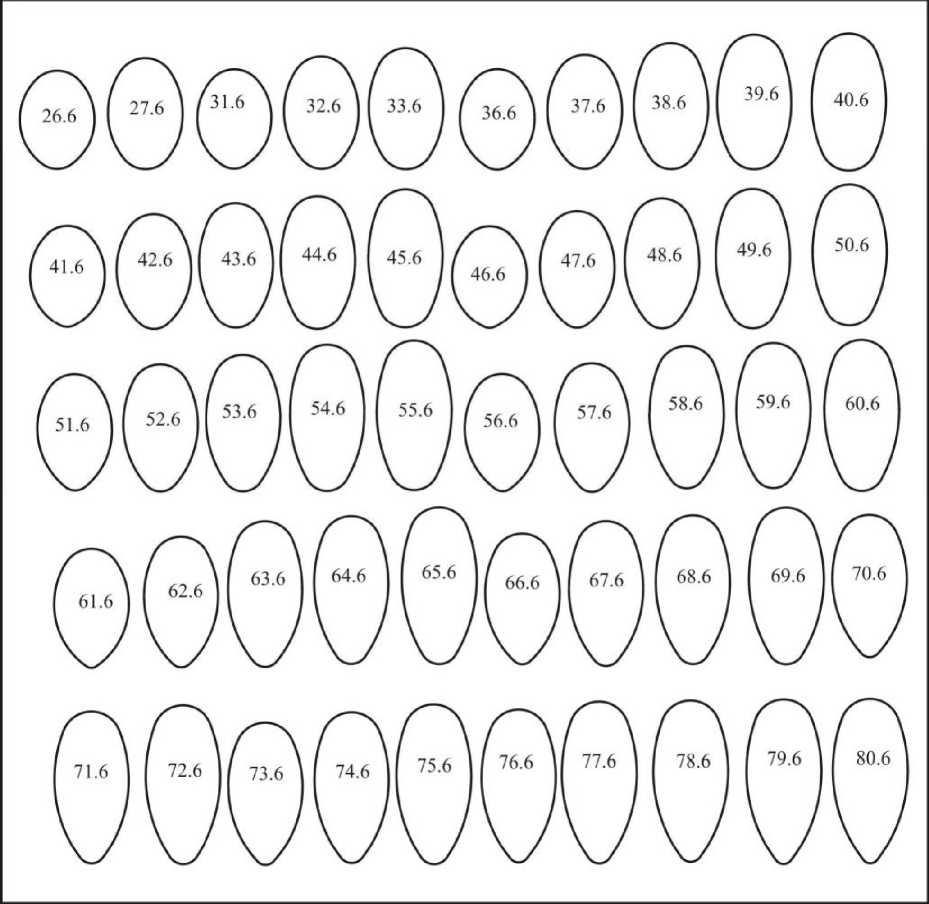
Рис. 19. Асимметрические псевдоовоиды шестой группы
Группа представлена, главным образом, в отрядах Anseriformes, Ciconnformes, Podicipediformes и Pelecaniformes. Такие отряды, как Gaviifonnes, Falconiformes, Caprimulgiformes, Gruiformes и Passeriformes представлены одинаково незначительно; остальные (Caprimulgiformes, Charadriiformes, Galliformes, Upupiformes, Strigifonnes, Piciformes) -единичными экземплярами.
Приведенное выше количество эталонов (п=375) выведено только по средним значениям инфундибулярного радиуса шести групп асимметрических псевдоовоидов. Если по той же схеме вовлечь минимумы и максимумы, то мы получим еще 750 эталонов.
Заключение. Предлагаемая система эталонов форм птичьих яиц дает несколько важных перспектив в оологических исследованиях. Присвоение фиксированного названия и количественного выражения формам яиц путем сопоставления их фотографий с геометрическими эталонами позволяет осуществлять привязку важной биологической информации к любому птичьему яйцу. Простота методики дает
Вестник ТвГУ. Серия "Биология и экология". 2014. № 1 возможность широкого ее применения на различных уровнях: визуальном и компьютерном. В последнем случае возможно использование как существующие, так и специально написанные программы. Унификация и скоординированность оологических работ орнитологов открывает большие перспективы для широкомасштабных обобщений и создания глобальных баз данных.
Митяй И.С. Описание форм птичьих яиц с помощью геометрических эталонов / И.С.
Митяй // Вести. ТвГУ. Сер. Биология и экология. 2014. № 1. С. 125-147.
Список литературы Описание форм птичьих яиц с помощью геометрических эталонов
- Климов С.М. 1993. Форма яйца птиц и метод ее расчета//Современные проблемы оологии: материалы I Междунар. конф. стран СНГ. Липецк. С. 63-65.
- Костин Ю.В. 1977. О методике ооморфологических исследований и унификации описаний оологических материалов//Методики исследования продуктивности и структуры видов птиц в пределах их ареалов: сб. науч. ст. Ч. 1. Вильнюс. С. 14-22.
- Митяй И.С. 2003. Новая методика комплексной оценки формы яйца//Бранта. Вып. 6. С. 179-192.
- Митяй И.С. 2008. Использование современных технологий в исследованиях птичьих яиц//Вiсник ЗНУ: зб. наук. ст. Бiол. науки. Запорiжжя: ЗНУ. Вип. 1. С. 191-200.
- Мянд Р. 1988. Внутрипопуляционная изменчивость птичьих яиц. Таллин: Валгу. 195 с.
- Anderson M. 1978. Optimal egg shape in waders//Ornis Fennica. V. 55. P. 105-109.
- Baker D.E. 2002. A geometric method for determining shape of bird eggs//Auk. V. 119. P. 1179-1186.
- Barta Z., Székely T. 1997. The optimal shape of avian eggs//Functional Ecology. V. 11. P. 656-662.
- Bridge E.S., Boughton R.K., Aldredge R.A., Harrison T. E., Bowman R., Schoech S.J. 2007. Measuring egg size using digital photography: testing Hoyt`s method using Florida Scrub-Jay eggs//Journal of Field Ornithology. V. 78. P. 109-116.
- Führer-Nagy G. 2002. Mathematical modeling of bird egg curves//Termeszetvedelmi Hivatal. Aquila. V. 107-108. P. 67-73.
- Gotman J., Jablonski В. 1972. Gniazda naszych ptakow. Warszawa: Panstwowe zaklady wydawnictw szkolnych. 282 s.
- Makatsch W. 1976. Die Eier der Vögel Europas. Band 1, 2. Neumann Verlag. S. 468-460.
- Monus F., Barta Z. 2005. Repeatability analysis of egg shape in a wild tree sparrow (Passer montanus) population: a sensitive method for egg shape description//Acta Zool. Hung. V. 51. P. 151-162.
- Narushin V.G. 2005. Egg geometry calculation using the measurements of length and breadth//Poultry Science. V. 84. P. 482-484.
- Preston F.W. 1953. The shapes of birds egg//Auk. V. 70. P. 160-182.
- Romanoff A.L., Romanoff A.J. 1949. The avian egg. New York: Wiley. 918 p.
- Schonwetter M. 1985. Handbuch der Oologie//Leiferung 8. Berlin. 154 p.
- Todd P.H., Smart I.H.M. 1984. The shape of birds' eggs//J. Th. Biol. V. 106. P. 239-243.
- Walters M. 1994. Birds' eggs. London: Dorling Kindersley. 256 p
