Определение наследуемости в популяциях растений при сочетании перекрестного опыления и самоопыления

Автор: Астахов А.И.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Методика
Статья в выпуске: 1 т.40, 2005 года.
Бесплатный доступ
На основе принципа интегрированного вклада в признак гетеро- и гомозиготных локусов предложена формула определения коэффициента наследуемости в смежных поколениях реальных селекционных популяций растений при разном сочетании перекрестного опыления и инбридинга.
Короткий адрес: https://sciup.org/142133048
IDR: 142133048 | УДК: 631.523.5
Determination of hereditability in plant populations at the combination of cross-pollination and self-pollination
On the basis of principle of integrated contribution to the determinant of hetero- and homozygous loci the author proposes the formula for determination of coefficient of hereditability in adjacent generations of real selectional plant populations at the different combinations of cross-pollination and inbreeding.
Текст научной статьи Определение наследуемости в популяциях растений при сочетании перекрестного опыления и самоопыления
Как известно, фенотипическое разнообразие количественного признака в популяции складывается под влиянием двух факторов — наследственных и ненаследственных (1, 2). Если принять сумму всех влияний на признак за единицу, то можно записать:
Н + Е = 1, [1]
где Н — коэффициент наследственной детерминации признака, Е — степень разнообразия признака в популяции, которая определяется влиянием среды. Поскольку невозможно непосредственно оценить генотип особи, Wright предложил другое решение этой проблемы с помощью метода коэффициентов путей (3, 4). При этом он исходил из следующих положений: связи между родительскими формами и потомками приближенно прямолинейны; все влияния на формирование признака суммируются в результате простого сложения, или аддитивны; все факторы действуют независимо друг от друга; степень детерминации признака каждым отдельным фактором равна квадрату коэффициента прямолинейной корреляции между признаком и этим фактором; популяция размножается посредством свободного скрещивания в условиях генетического равновесия по закону Харди-Вайнберга. При соблюдении этих условий коэффициент корреляции между генотипами потомков и родительских форм равен 0,5.
Предлагая схему коэффициентов путей, Wright использовал понятия генотип и фенотип. Позже Fischer предложил различать понятия фенотип, генотип и генетическая формула (в последнем случае учитывается число доминантных генов) (5). На основе этих понятий Никоро разработала более детальную схему коэффициентов путей для обоснования теории наследуемости (рис.) (6).
Мы не задаемся целью анализировать все взаимодействия между генотипами, генетическими формулами и фенотипами родительских форм и потомков, а ограничиваемся более узкой задачей — определить аддитивную наследуемость на основе коэффициентов путей Wright. Рассмотрим путь от фенотипа материнской формы к фенотипу потомка. При условии независимости действия и прямолинейности связей коэффициент корреляции будет равен произведению коэффициентов путей: r = h2⋅c2⋅a2⋅b2⋅c3⋅h3. Если принять, что в смежных поколениях h2 = h3, с2 = c3, то r = c2h2ab, откуда c h = rфенотип мать→потомок ,
ab где с2h2 — доля влияния генетической формулы на разнообразие фенотипов.
Это и есть коэффициент наследуемости в узком смысле, ко-
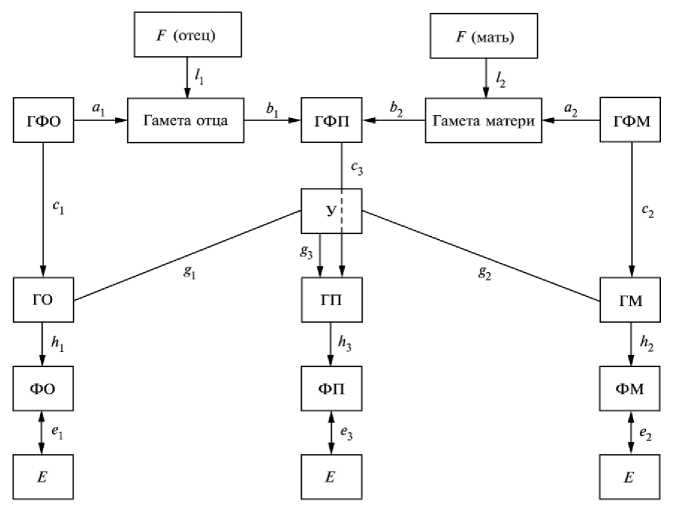
Корреляционные связи между генотипом и фенотипом родительских форм и потомков в смежных поколениях (цит. по 6 ист.): ГФО, ГФП, ГФМ, ГО, ГП, ГМ, ФО, ФП, ФМ, F , Е и У — соответственно генетическая формула отца, генетическая формула потомка, генетическая формула матери, генотип отца, генотип потомка, генотип матери, фенотип отца, фенотип потомка, фенотип матери, коэффициент инбридинга, степень разнообразия признака в популяции и уникальная формула генотипа потомка на основе рекомбинации материнских и отцовских генов; а — коэффициент пути между генетической формулой родителя и гаметой; b — коэффициент пути между гаметой родителя и генетической формулой потомков; с — путь влияния генетической формулы на разнообразие генотипов, то есть роль аддитивности в создании разнообразия; h — путь наследуемости в широком смысле слова; g — путь влияния аллельного и неаллельного взаимодействия; l — путь закономерностей расщепления; е — путь влияния условий среды.
который отражает долю аддитивного разнообразия в общем фенотипическом разнообразии.
Таким образом, коэффициент наследуемости в узком смысле определяется фенотипическим сходством родительской формы и потомков и зависит от коэффициентов путей а и b ; а · b = r (коэффициент генетической корреляции, который определяет связь между генетическими формулами родительской формы и потомков). В условиях равновесия популяции при свободном скрещивании а ⋅ b = 0,5. Подставляя это значение в выражение [2], получим
c h = 2 r фенотип мать→потомок .
Эту формулу используют для определения коэффициента наследуемости в узком смысле; она дает объективную информацию при указанных ограничениях. Однако в реальных условиях работы селекционеров по этой формуле можно весьма приближенно (или совсем неверно) оценить наследуемость. Во-первых, селекционеры всегда работают не с панмиктической, а с ограниченной популяцией, в которой они ведут искусственный отбор и ограничивают свободу скрещиваний. В результате отбора изменяются концентрация и частота отдельных генов, влияющих на конкретный признак. Во-вторых, в результате ограниченного числа особей в скрещиваниях или тесного инбридинга в популяции происходит снижение доли гетерозиготности и увеличение доли гомозиготности, что также оказывает влияние на проявление признака. Поэтому при расчете коэффициента наследуемости по формуле [3] часто возникает бессмыслица: c 2 h 2 > 1 или с 2 h 2 < 0 .
Рассмотрим случай прямо противоположный свободному скрещиванию
— самоопыление особей из перекрестноопыляемой популяции. Представим себе какой-то количественный признак, фенотипическое выражение которого зависит от многих генов, и на него работает n аллелей, одна часть из которых может находиться в гетерозиготном, а другая — в гомозиготном состоянии. Гетерозиготные локусы при самоопылении растений будут выщеплять гомозиготы, не нарушая равновесия:
Аа х Аа
г
АА + 2 Аа + аа
2 х 1 х 1
)2; 1 х 1 = 1.
1х1 = (--------- r
При этом условии а ■ b = 0,5; с1 h2 = --- = 2 r .
0,5
Гомозиготные локусы при самоопылении не расщепляются:
АА х АА г 4АА .
В этом случае коэффициент корреляции между генетической формулой материнской формы и гаметой, а также между гаметой и генетической формулой по-
22 r r томка равен единице. Тогда с h = ----=---= r . Действительно, при полной го- a х b 1 х 1
мозиготности потомство подобно материнской форме, что обычно наблюдается при размножении инбредных линий.
Для определения сходства родительских форм и потомства используют коэффициент внутриклассовой корреляции rw. Рассчитывая этот коэффициент, мы фиксируем интегральный вклад гетеро- и гомозиготных локусов в конкретный признак. Вероятно, для того чтобы определить долю наследования с учетом аддитивного влияния этих локусов на проявление признака, необходимо оценить вклад локуса по средневзвешенному признаку на основе вышеизложенных закономерностей:
22 rw х доля гомозиготных локусов + 2 rw х доля гетерозиготных локусов
.
сh = доля гомозиготных локусов + доля гетерозиготных локусов
При сочетании перекрестного опыления и самоопыления доля растений, размножающихся в каждом поколении посредством самоопыления и свободного опыления, определяется следующим соотношением (7):
w1 _ 2 F
1 - w 1 - F ’ где w1 и (1 – w1) — доля растений в популяции, размножающихся в каждом поколении посредством соответственно самоопыления и свободного опыления; F — коэффициент инбридинга, или вероятная доля гомозиготности; 1 – F — относительный уровень гетерозиготности. Это же соотношение справедливо для гомо- и гетерозиготных локусов.
Подставив значения доли гомо- и гетерозиготности в вышеприведенное выражение, получим
2,2 r w ■ 2 F + 2 r w ■ (1 - F)
сh =
2 F + (1- F )
После математического преобразования формула [4] принимает вид
2r w с2h2 = , [5]
В реальной ситуации коэффициент c 2 h 2 будет иметь разные значения в зависимости от давления естественного и искусственного отборов и объема популяции, из которой селекционер отбирает особей для скрещивания или самоопыления.
В случае полной гетерозиготности родительских форм с 2 h 2 принимает максимальное значение: с2h2 - 2 r w родительская форма^потомок ( F = 0). Эта формула применяется для определения доли аддитивного разнообразия в равновесной свободно скрещивающейся популяции. По мере уменьшения гетерозиготности материнской формы (или обеих родительских форм) по рассматриваемому признаку коэффициент с 2 h 2 будет уменьшаться. Вероятно, относительное уменьшение c 2 h 2 в смежных поколениях при одинаковой схеме селекции будет тем меньше, чем больше генов работает на конкретный признак.
При полной гомозиготности локусов, детерминирующих признак ( F = 1), формула принимает вид с 2 h 2 - r w родительская форма^потомок . В этом случае коэффициент наследуемости c 2 h 2 будет иметь минимальное значение.
Таким образом, определив вероятную долю нарастания гомозиготности и коэффициент внутриклассовой корреляции между родительской формой и потомком, можно рассчитать коэффициент наследуемости в узком смысле в смежных поколениях любых селекционных популяций растений по формуле [5]. При этом отпадают ограничения панмиксии и генетического равновесия в оцениваемых популяциях.
Л И Т Е Р А Т У Р А
-
1. W r i g h t S. The relative importance of heredity and environment in determining the piebald pattern of guinea pig. Proc. Nat. Acad. Sci., 1920, 6: 320-332.
-
2. W r i g h t S. Systems of mating. Genetics, 1921, 6: 111-178.
-
3. W r i g h t S. The theory of path coefficients. Genetics, 1923, 8.
-
4. W r i g h t S. The method of path coefficients. Ann. Math. Stat., 1934, 5, 3: 161-215.
-
5. F i s c h e r R.A. The genetical theory of natural selection. N.Y., 1930.
-
6. Н и к о р о З.С., С т а к а н Г.А., Х а р и т о н о в а З.Н. и др. Теоретические основы селекции животных. М., 1968: 176-199.
-
7. Л и Ч. Введение в популяционную генетику. М., 1978.
Поступила в редакцию 20 мая 2004 года
Всероссийский НИИ люпина, 241524, Брянск, п/о Мичуринское; e-mail: lupin@online.bryansk.ru
DETERMINATION OF HEREDITABILITY IN PLANT POPULATIONS AT THE COMBINATION OF CROSS-POLLINATION AND SELF-POLLINATION
S u m m a r y
On the basis of principle of integrated contribution to the determinant of hetero- and homozygous loci the author proposes the formula for determination of coefficient of hereditability in adjacent generations of real selectional plant populations at the different combinations of cross-pollination and inbreeding.
Новые книги
К ш н и к а т к и н а А.Н., Г у щ и н а В.А., В а р л а м о в В.А. и др. Технология выращивания и использования нетрадиционных кормовых и лекарственных растений /Под ред. А.Н. Кшникаткиной. М.: ВНИИССОК, 2003, 373 с.
В монографии на основе многолетних экспериментальных исследований авторов и обобщения данных литературы и производственного опыта рассматривается состояние и перспективы возделывания нетрадиционных кормовых и лекарственных растений. Дана агроэкологическая оценка возделывания многолетних трав, козлятника восточного, топинамбура, тописолнечника, левзеи сафлоровидной, свербиги восточной, кормового щавеля, рапса. Приведены оптимальные технологические приемы выращивания расторопши пятнистой, эхинацеи пурпурной. Проанализированы закономерности роста и развития растений. Обсуждается значение интродукции в расширении видового состава кормовых культур и лекарственных растений.
Л а п а В.В., Б о с а к В.Н. Оптимальные дозы азотных удобрений под сельскохозяйственные культуры (рекомендации) . Минск, 2002, 23 с.
В рекомендациях рассматривается ресурсосберегающая система удобрения, используемая при интенсивных технологиях возделывания основных сельскохозяйственных культур. Приведены нормативные материалы по применению минеральных и органических удобрений на дерново-подзолистых и торфяно-болотных почвах с учетом содержания подвижных форм фосфора и калия, биологических особенностей и планируемой урожайности растений. Описаны системы удобрения озимых и яровых зерновых, зернобобовых, пропашных культур, однолетних и многолетних трав, сенокосов и пастбищ.
П а л ф и т о в В.Ф. Диагностика самоплод-ности и силы роста яблони. Мичуринск: изд-во МГАУ, 2003, 198 с.
В монографии приведены разработанные автором методы ранней диагностики само-плодности деревьев и силы роста подвоев яблони. Описаны способы количественного определения содержания различных веществ в пыльце, пестиках и других частях и органах растений. Отмечено, что при склонности к самоплодности наблюдается повышенное содержание водорастворимых соединений в пыльце и пониженное количество флоризина (полифенольный глюкозид) в пестиках. Пред- ставлены данные по диагностике силы роста подвоев яблони на основе количества флоризина в зоне корневой шейки. Рассматривается возможность применения флоризина в практике садоводства, в частности для укоренения зеленых черенков у сортов и подвоев яблони с различной силой роста.
П о м а з к и н а Л.В., К о т о в а Л.Г., Л у б н и н а Е.В. и др. Устойчивость агроэкосистем к загрязнению фторидами. Иркутск: изд-во Института географии СО РАН, 2004, 225 с.
В монографии описан разработанный и апробированный авторами подход к исследованию агроэкосистемы, позволяющий интегрально оценивать режим функционирования последней и степень антропогенной нагрузки. Рассматриваются негативные последствия техногенного загрязнения фторидами всех компонентов агроэкосистемы (почва— микроорганизмы—растения—атмосфера). Режим функционирования агроэкосистем предлагается оценивать на основе интегральных показателей цикла азота, которые количественно определяют при помощи изотопа N15. Показано, что в условиях загрязнения усиливаются минерализация и газообразные потери азота и углерода, а также снижается (ре)им-мобилизация, что приводит к дефициту этих веществ в почве. Отмечено, что устойчивость агроэкосистемы в целом выше, чем ее отдельных компонентов, однако при загрязнении снижается. Рассматриваются особенности микробиологической трансформации органического вещества в агроэкосистемах на загрязненных фторидами почвах. Дана оценка влияния степени загрязнения почв фторидами на обеспеченность минеральными веществами и продуктивность полевых культур. Результаты исследований могут быть положены в основу новых экологических нормативов допустимого антропогенного воздействия на окружающую среду.
