Определение озерного байкальского эндемического амфиполя Eulimnogammarus verrucosus (Gerstf., 1858). Термический оптимизм ограничивается изменениями его метаболических маркеров
. Термический оптимизм ограничивается изменениями его метаболических маркеров")
Автор: Аксенов-грибанов Д.В., Лубяга Я.А., Шахтанова Н.С., Гурков А.Н., Бедулина Д.С., Шатилина Ж.А. М., Кондратьева Е.М., Верещагина К.П., Тимофеев М.А.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 4 т.8, 2012 года.
Бесплатный доступ
В настоящем исследовании было проведено определение клеточных метаболических маркеров под воздействием эндемического амфипода Eulimnogammarus озера Байкал (Amphipoda, Crustacea) в широком диапазоне температур окружающей среды. Метаболические данные сравнивались с данными о терминопрефердуме вида, полученными в наших предыдущих исследованиях. Было обнаружено, что воздействие амфипод при повышенных температурах приводит к увеличению содержания HSP70 и лактата, активации антиоксидантных ферментов (каталазы и пероксидазы), а также снижению активности лактатдегидрогеназы и глутатиона S-трансферазы. Воздействие E. verrucosus при низких температурах приводило к уменьшению содержания HSP70, увеличению уровня лактата, активации пероксидазы и лактатдегидрогеназы и снижению активности S-трансферазы глутатиона. Было отмечено, что наиболее выраженные изменения маркеров метаболизма у видов, наблюдаемых при превышении температурного порога, превышали 11 ° С или затем температура опускалась ниже 4 ° С. Температурный диапазон 4-11 ° C представлял собой область стабильности клеточных метаболических маркеров в E. verrucosus и коррелировал с предпочтительными температурными пределами, ранее полученными в поведенческих экспериментах. Таким образом, впервые с амфиполями озера Байкал мы показали, что зона стабильности клеточного метаболизма в E. verrucosus тесно связана с их поведенческими зонами тепловых предпочтений и, вероятно, отражает температурные оптические пределы видов.
Короткий адрес: https://sciup.org/14323963
IDR: 14323963
Determination of lake Baikal endemic amphipod Eulimnogammarus verrucosus (Gerstf., 1858) thermal optima limits by changes in its metaboliс markers
In present study the determination of cellular metabolic markers under exposure of Lake Baikal endemic amphipod Eulimnogammarus verrucosus (Amphipoda, Crustacea) in a wide range of ambient temperatures was carried out. The metabolic data was compared with species termopreferendum data obtained in our previous studies. It was found that exposure of amphipods under increased temperatures resulted in increase of HSP70 content and lactate, activating of antioxidant enzymes (catalase and peroxidase), as well as reduction of lactate dehydrogenase and glutathione S-transferase activities. Exposure of E. verrucosus at low temperatures resulted in decrease of HSP70 content, increase of lactate level, peroxidase and lactate dehydrogenase activation, and reduction of glutathione S-transferase activity. It was noted that the most expressed changes of metabolism markers in the species observed when temperature threshold was exceeded 11°C or then temperature dropped below 4°C. The temperature range 4-11°C represented the area of stability of cellular metabolic markers in E. verrucosus and correlated with a preferred temperature limits, previously obtained in behavioral experiments. Thus, for a first time with Lake Baikal amphipods we showed that the zone of stability of cellular metabolism in E. verrucosus closely related to their behavioral thermal preference zones and is likely to reflect thermal optima limits of the species.
Текст научной статьи Определение озерного байкальского эндемического амфиполя Eulimnogammarus verrucosus (Gerstf., 1858). Термический оптимизм ограничивается изменениями его метаболических маркеров
В ходе исследования у байкальских эндемичных амфипод Eulimnogammarus verrucosus (Amphipoda, Crustacea) проводили оценку изменений показателей клеточного метаболизма при экспонировании рачков в широком диапазоне температур окружающей среды. Полученные данные были сопоставлены с известными показателями термопреферендума вида. Выявлено, что при градиентном повышении температуры среды у E. verrucosus происходит увеличение содержания БТШ70 и лактата, активация антиоксидантных ферментов (каталазы и пероксидазы), а также снижение активности лактатдегидрогеназы и глутатион S-трансферазы. Экспозиция E. verrucosus в условиях низких температур вела к снижению содержания БТШ70, увеличению содержания лактата, активации пероксидазы и лактатдегидрогеназы, а также снижению активности глутатион S-трансферазы. Отмечено, что наиболее выраженные изменения показателей клеточного метаболизма у вида происходят при превышении температурного порога в 11°С и при снижении температуры ниже 4°С. Установленный температурный диапазон 4-11°С отражает зону стабильности показателей клеточного метаболизма у E. verrucosus и коррелирует с диапазоном предпочитаемых видом температур, выявленным ранее в поведенческих экспериментах.
Таким образом, в работе впервые для байкальских амфипод установлено, что зона стабильности показателей клеточного метаболизма у E. verrucosus тесно связана с его термопреферентной зоной и, вероятно, отражает границы температурного оптимума данного вида.
Температура является важнейшим экологическим фактором среды, от которого напрямую зависят обмен веществ и развитие водных организмов (Pörtner, Knust 2007). Именно от температуры в большей степени зависят конформация и активность макромолекул, интенсивность метаболизма и работа электрон-транспортной цепи в клетке (Хочачка, Сомеро, 1988). Смена температуры среды приводит к изменениям функциональной активности ряда биологических катализаторов, которые составляют основу системы регуляции метаболизма (Pörtner, Farell, 2008).
Большинство пойкилотермных водных организмов приспособлено к обитанию в определенном температурном диапазоне, в пределах которого их метаболические процессы функционируют с наибольшей эффективностью. Отклонение параметров среды от оптимальных, ведет к изменению характера функционирования метаболических процессов и, в случае критических величин, к развитию стрессовой реакции. Большинство гидробионтов, способных к активному перемещению в среде, развили выраженные термопреферентные поведенческие реакции, позволяющие индивидууму выбирать ту или иную температурную зону.
Экспериментальное определение зоны термопрефрендума является одним из подходов, широко применяемых для оценки границ температурного оптимума вида. Другой подход представляет собой оценку терморезистентных способностей видов. Распространенная практика определения терморезистентных показателей в условиях экспозиционных экспериментов достаточно информативна с точки зрения межвидового сопоставления, однако она слабо отражает экологические характеристики видов. Случаи быстрых критических изменений температуры среды возможны (например, при ветровых подвижках воды), но они редко встречаются в естественных водоемах. Большинство изменений показателей среды обитания происходит постепенно и активные организмы способны избегать условия, выходящие за пределы предпочитаемых параметров. В связи с этим довольно сложно определить, как именно изменения параметров среды отражаются на метаболических процессах индивидуумов и в какой мере показатели терморезистентности или термопреференции отдельных видов связаны с характеристиками клеточного метаболизма. Иными словами, как именно температурный оптимум метаболических процессов в организме связан с экологическими характеристиками вида и зоной его обитания?
Для оценки особенностей взаимосвязи параметров клеточного метаболизма с температурным оптимумом вида наиболее интересными являются организмы, которые эволюционировали в стабильных условиях среды и не сталкивались с ее критическими изменениями. Одним из таких мест, характеризующихся высокой стабильностью условий среды обитания, является озеро Байкал (возраст более 25 млн. лет). Фауна озера представляет порядка 2600 видов, 80% из которых являются эндемиками (Тимошкин, 2004; Беркин, 2009). В ходе данного исследования у байкальских эндемичных амфипод Eulimnogammarus verrucosus (Amphipoda, Crustacea) проводили оценку изменений показателей клеточного метаболизма при экспонировании рачков в широком диапазоне температур окружающей среды.
МАТЕРИАЛЫ И МЕТОДЫ
Объектом настоящего исследования были выбраны байкальские эндемичные амфиподы Eulimnogammarus verrucosus (Gerstf., 1858).
Данный вид является одним из самых широко распространенных видов в Байкале. E. verrucosus - типичный обитатель литорали, встречается повсеместно под камнями, начиная с глубины в несколько сантиметров вплоть до 10 - 15 метров (Базикалова, 1945). Фитофаг (Kamaltynov, 1999). Имеет зимний период размножения. Окончание репродуктивного периода в июле (Базикалова, 1945; Гаврилов, 1949; Бекман, Деньгина, 1969). По сообщениям ряда авторов, взрослые представители вида в летние месяцы отходят на б о льшие глубины (Вейнберг, 1995; Вейнберг, Камалтынов, 1995, 1998).
Из ранее опубликованных нами материалов по термопреферендуму амфипод, известно, что E. verrucosus холодолюбивый вид. В экспериментальной установке в условиях температурного градиента взрослые особи вида концентрировались в диапазоне температур от 3-4 до 9-10°С с пиком предпочитаемых температур в зоне 5-6°С (Timofeyev, Shatilina, 2007). При этом в температурном диапазоне от 3-4 до 9-10°С в установках концентрировалось более 80% рачков.
Для текущего исследования взрослые особи E. verrucosus были выловлены в литоральной зоне (0 - 1,5 м) в районе пос. Листвянка. Сбор бокоплавов производили с использованием гидробиологического сачка. До экспериментов рачков преакклимировали в аэрируемых термостатах при температуре 6(±0,5) °С в течение 3 - 4 суток. При этом температура преакклимации была приближена к среднегодовому показателю температуры зоны обитания рачков. В преакклимационный период животных кормили препаратом Tetramin (Германия). Эксперименты проводили со здоровыми и активными особями.
В ходе исследования проводили 2 типа экспериментов: первый – экспозиция E. verrucosus в условиях градиентного (постепенного) повышения температуры среды от 6°С (принимаемой за контрольный показатель) до температуры, при которой отмечали гибель 50% особей; второй – экспозиция в условиях градиентного понижения температуры среды от контрольного уровня до минимальных положительных температур (0,5°С). Скорость изменения температуры составила 1°С•ч-1. Эксперименты проводили в цифровом ультратермостате WiseCircu Wcr6 (Германия).
Фиксацию материалов проводили в жидком азоте: при экспозиции гидробионтов в условиях повышения температуры - каждые 2°С (2 часа); при экспозиции в условиях понижения температуры – каждый 1°С (1 час). Контрольные образцы фиксировали непосредственно в начале эксперимента при температуре 6°С.
Для оценки влияния изменения температуры среды на показатели клеточного метаболизма (в том числе стресс-маркеров) определяли изменение содержания белков теплового шока БТШ70 и лактата, активности ферментов антиоксидантной системы (пероксидазы, каталазы, глутатион S – трансферазы) и фермента анаэробного гликолиза -лактатдегидрогеназы.
На начальном этапе работы проводили эксперименты по оценке выживаемости амфипод при экспозиции в условиях повышения и понижения температуры среды. Далее была определена степень воздействия изменения температуры среды на содержание БТШ70. Изменение содержание БТШ70 определяли с использованием стандартного метода денатурирующего электрофореза с ДДС-Na в 10 % полиакриламидном геле c последующим Вестерн-блоттингом (Willmer at al, 2000) и инкубацией мембран в антителах к БТШ70 (monoclonal anti-heat shock protein 70, produced bovine brain, Sigma Chemical Co) и антителах к актину (polyclonal Anti-Actin antibody produced in rabbit, Sigma Chemical Co). Актин применяли в качестве референтного белка. Денситометрический анализ содержания белка на мембранах проводили с помощью программы ImageJ v.1,41.
На следующем этапе работы был определен характер изменения активности ферментов антиоксидантной системы (АОС) у амфипод, экспонированных в условиях различных температур. Оценку активности ферментов АОС (пероксидазы, каталазы и глутатион S-трансферазы) проводили согласно модифицированным спектрофотометрическим методикам Drotar (1985), Aebi (1984) и Habig (1974) соответственно (Тимофеев, 2010). На заключительном этапе работы была проведена оценка изменения содержания лактата и активности лактатдегидрогеназы. Данные измерения проводили энзиматическими, спектрофотометрическими методами с применением стандарт-наборов «Лактат-витал» и «ЛДГ-витал» (Vital– Diagnostics Spb) (Павличенко и др, 2009). Спектрофотометрический анализ образцов был проведен на спектрофотометре Cary 50 (Varian, США) при λ=340 нм для определения активности пероксидазы и лактатдегидрогеназы, при λ=240 нм для каталазы и при λ=436 нм для глутатион S – трансферазы. Определение содержания лактата проводили при λ=505 нм
Все эксперименты проведены в 9 биологических повторностях. Биохимический анализ каждой пробы был проведен в 3-х аналитических измерениях. Оценку достоверности проводили, используя двувыборочный u-критерий Манна-Уитни. Статистический анализ проводили с использованием программы Statistica 8.0. На диаграммах указаны доверительные интервалы. * - обозначены случаи достоверного отличия значений от контрольного при доверительной вероятности – 0,95.
РЕЗУЛЬТАТЫ
На рисунке 2 представлены материалы по оценке выживаемости амфипод E. verrucosus при экспозиции в условиях изменения температуры среды. Как следует из представленных материалов, при повышении температуры среды со скоростью 1°С•ч-1, температура гибели 50 % особей (LT50) составила 29,7°С , а LT100 - 30,5°С (рис. 2). Смертность амфипод при понижении температуры среды не отмечали.
На рисунке 3 представлены материалы по оценке изменения содержания БТШ70. В диапазоне температур с 4°С до 9°С, уровень БТШ70 достоверно не отклонялся от контрольного содержания БТШ70. Отмечали двукратное (относительно контрольного уровня) увеличение содержания БТШ70 уже при достижении температуры 11°С. На более поздних этапах экспозиции (при повышении температуры среды до 29°С) у амфипод отмечали снижение содержания БТШ70 на 55% относительно контрольных значений. При других повышенных температурах, содержание БТШ70 не отличалось от контрольных значений.
Экспозиция E. verrucosus в условиях низких температур вела к снижению содержания БТШ70 относительно контрольных значений на 37 - 58% при температуре экспозиции ниже 3°С. При данных условиях, пониженное содержание БТШ70 сохранялось до окончания экспозиции.
Материалы по оценке изменения активности ферментов АОС у E. verrucosus даны на рисунке 4. Как видно из представленных данных (рис. 4А) в диапазоне температуры среды с 4°С до 11°С, активность пероксидазы была стабильной и не отклонялась от контрольного значения. При повышении температуры среды у E. verrucosus до 13°С отмечено увеличение активности пероксидазы на 38% и далее к 17°С - на 42%. При других температурах активность пероксидазы флуктуировала около контрольных показателей. При снижении температуры среды до 3оС, у амфипод происходила активация пероксидазы на 28%. Понижение температуры среды до 0,5°С приводило к снижению активности фермента на 33%.
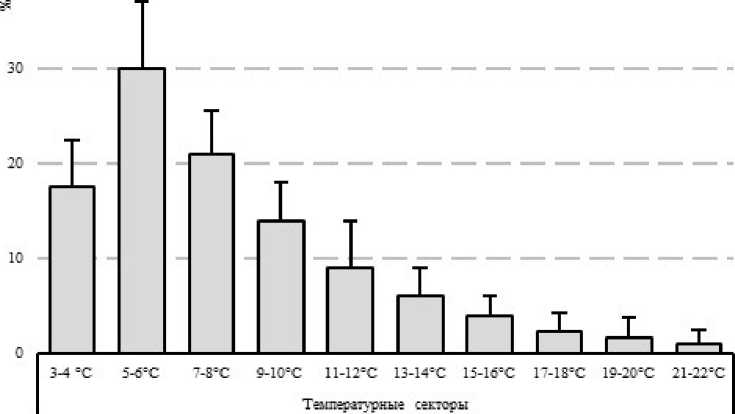
Рисунок 1. Распределение E. verrucosus в температурном градиенте, % от общего числа.
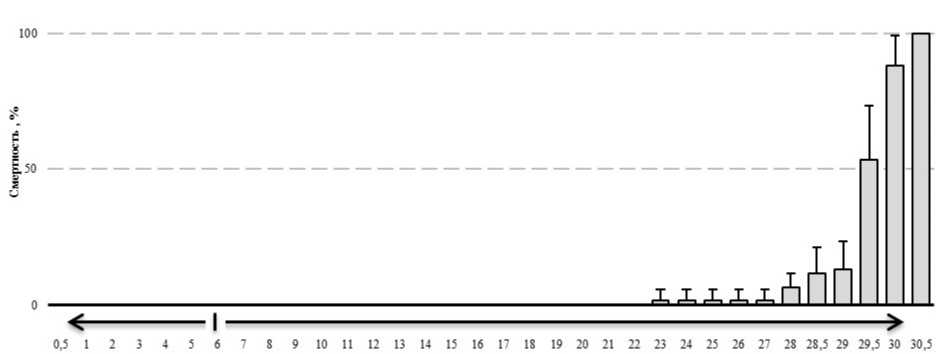
Рисунок 2. Смертность E. verrucosus в условиях повышения температуры среды (в процентах)
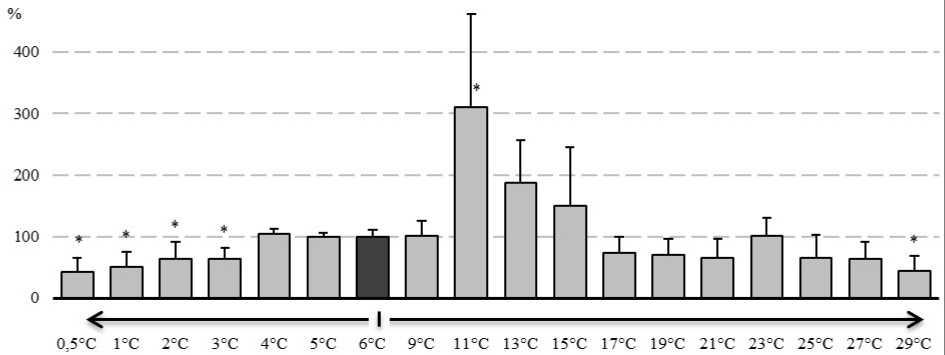
Рисунок 3. Изменение содержания БТШ70 у E. verrucosus при изменении температуры среды (в процентах, относительно содержания БТШ70 при 6°С)
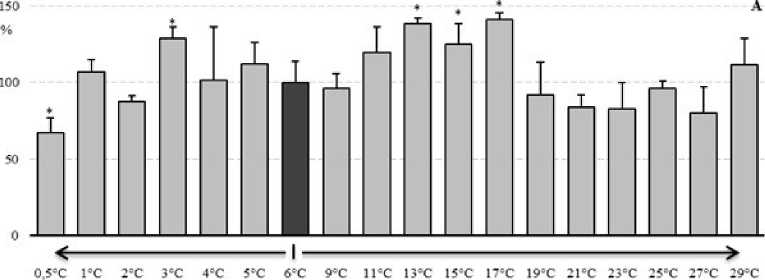
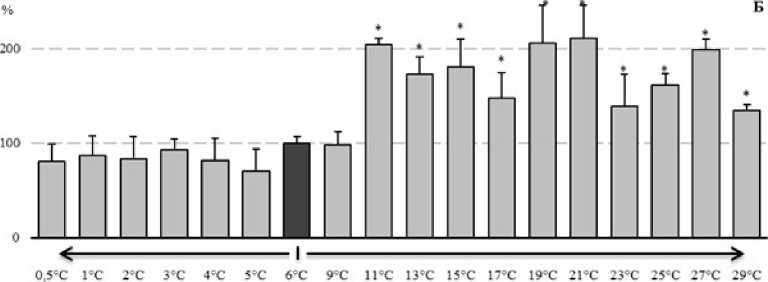
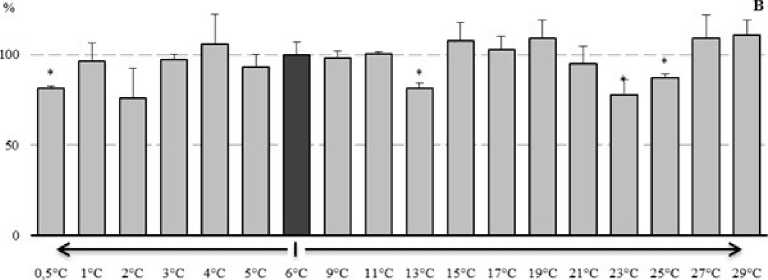
Рисунок 4. Изменение активности ферментов АОС у E. verrucosus при изменении температуры среды (в процентах, относительно активности ферментов при 6°С): А – пероксидаза, Б – каталаза, В – глутатион S-трансфераза
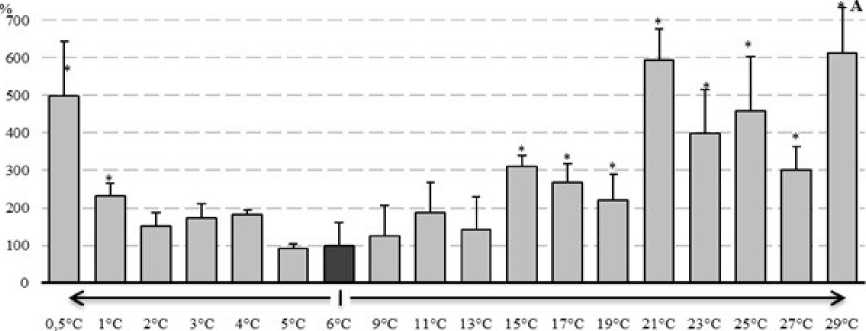
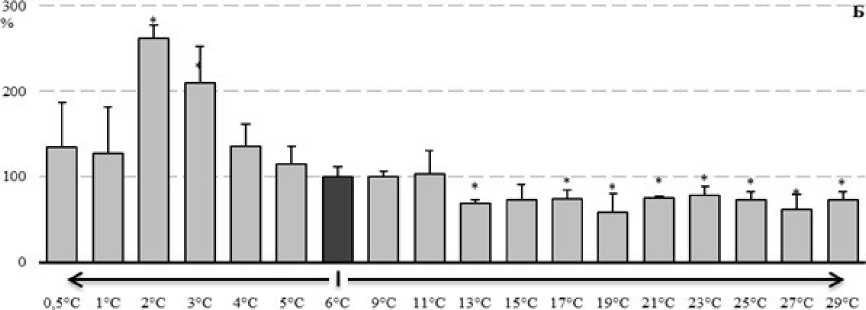
Рисунок 5. Изменение содержания лактата (А) и активности лактатдегидрогеназы (Б) у E verrucosus при изменении температуры среды (в процентах, относительно 6°С)
Как следует из представленных материалов (рис. 4Б), при температуре среды выше 11°С у исследуемых амфипод происходило двукратное увеличение активности каталазы относительно контрольного уровня. При этом повышенная активность каталазы сохранялась до окончания экспозиции. Понижение температуры не приводило к статистически значимым изменениям активности каталазы.
В ходе оценки активности глутатион S-трансферазы установлено (рис. 4В), что повышение температуры среды до 13°С ведет к статистически значимому снижению активности фермента. При дальнейшей экспозиции наблюдали флуктуацию показателей активности с достоверным снижением в периоды 23°С и 25°С. Понижение температуры среды вызывало достоверное снижение активности фермента на 25% при температуре 0,5°С.
Материалы оценки изменения содержания лактата и активности лактатдегидрогеназы даны на рисунке 5. Как следует из представленных материалов (рис. 5А), в диапазоне температуры от 2 до 13°С уровни содержания лактата варьировали в пределах контрольных значений. Повышение температуры среды до 15°С и выше вело к достоверному росту содержания лактата у исследованных амфипод. Показатели активности лактатдегидрогеназы (рис. 5Б) в диапазоне температур от 4 до 11°С не отличались от контрольного уровня. Повышение температуры до 13°С и выше вело к достоверному снижению активности фермента, а понижение до 4 и 3°С напротив к росту активности фермента.
ОБСУЖДЕНИЕ
В ходе проведенного исследования было установлено, что при повышении температуры среды у E. verrucosus происходит увеличение содержания БТШ70 и лактата, активация ферментов АОС (каталазы и пероксидазы), а также снижение активности лактатдегидрогеназы и глутатион S-трансферазы. Экспозиция E. verrucosus в условиях снижения температур вела к падению содержания БТШ70, увеличению содержания лактата, активации пероксидазы и лактатдегидрогеназы, снижению активности глутатион S-трансферазы, но не влияла на активность каталазы.
Учитывая тот факт, что использованные в работе биохимические параметры являются маркерами различных метаболических процессов, проходящих на клеточном уровне амфипод, следует рассмотреть возможные функциональные причины и последствия этих изменений.
БТШ70 являются одними из наиболее изученных маркеров стрессовых состояний. Будучи молекулярными шаперонами они восстанавливают нативную конформацию и функциональную активность белков (Farnsworth, Sign, 2000; Ito at al, 2003). Таким образом, синтез БТШ70 направлен на противодействие и (или) компенсацию негативных последствий процессов, связанных с изменением температуры среды. Накопление БТШ70 у E. verrucosus при отклонении температуры среды от оптимальной направлено на защиту от стрессового воздействия. Снижение содержания БТШ70, наблюдаемое при высоких температурах экспозиции и в условиях понижения температуры среды ниже 3°С, может указывать на угнетение метаболизма, деградацию БТШ70 и возрастающий энергодефицит клеток (Sokolova, 2004; Geret et al., 2002).
В условиях пессимальных температур происходит увеличение энергодефицита в клетках за счет нарушения работы электрон-транспортной цепи митохондрий, что приводит к переключению энергетического метаболизма с эффективного аэробного на менее эффективный анаэробный. Об активации анаэробиоза свидетельствуют представленные в нашей работе данные по изменению активности лактатдегидрогеназы и содержания лактата. Известно, что лактат является активным метаболитом, который способен перемещаться между клетками, тканями и органами, где он может быть окислен в качестве источника энергии или повторно преобразован в пируват или глюкозу. Также, лактат способен регулировать окислительно-восстановительные процессы в клетке и НАД+/НАДH баланс, тем самым регулируя метаболизм в разных тканях. Таким образом, совместно с лактатдегидрогеназой, лактат может выполнять роль псевдогормона, регулируя запасы глюкозы и гликогена в разных тканях и поддерживая его на нужном уровне, тем самым поддерживая энергетический баланс организма (Mark et al, 2002; Маянский, 1994).
Увеличение содержания лактата в тканях E. verrucosus было индуцировано как повышением, так и понижением температуры среды относительно оптимальной. На фоне изменения содержания лактата, активность лактатдегидрогеназы увеличивалась только при понижении температуры и снижалась - при её повышении. Из литературы известно, что снижение активности лактатдегидрогеназы отмечают в случаях высокого содержания лактата, недостатка кофермента НАДН, истощения запасов глюкозы, в результате чего происходит угнетение функционального и физиологического состояния организмов, что и отмечали в случае повышения температуры (Motohashi et al, 1999; Selvakumar, Geraldine, 2003; Kawall et al, 2002; Stillman, Somero, 2001). Установленное в нашей работе увеличение активности лактатдегидрогеназы при понижении температуры среды, может свидетельствовать о попытках амфипод нарастить долю анаэробного метаболизма, следствием которого и является последующее накопление лактата.
Одной из возможных причин смещения энергетического равновесия в сторону анаэробиоза может быть нарастание окислительных процессов в клетках и развитие оксидативного стресса, сопровождающегося накоплением АФК (Fridovich, 1998; Меньшикова, и др., 2006). На это в настоящем исследовании указывают изменения активности антиоксидантных ферментов. Рост активности пероксидазы и каталазы у E. verrucosus в условиях повышения температуры среды указывают на активацию АОС для элиминации активных форм кислорода и свободнорадикального окисления (Кулинский, 1999). Важно отметить, что у E. verrucosus при повышении температуры среды увеличение активности каталазы происходит раньше и при меньших температурах экспозиции, чем изменение активности пероксидазы. Вероятно, это свидетельствует о большей значимости каталазы в механизмах антиоксидантной защиты амфипод, по сравнению с пероксидазой. Это предположение уже обсуждалось нами и в ранее опубликованных работах (Тимофеев, 2010;
Timofeyev et al., 2006; Axenov-Gribanov, Lubyaga, 2012).
Среди механизмов выведения токсичных метаболитов из организма, важнейшее место занимает глутатион S-трансфераза. Одной из функций фермента является элиминация вторичных продуктов перекисного окисления липидов. Ранее нами уже были приведены данные об увеличении переоксилительных процессов у амфипод в условиях градиентного повышения температуры (Аксенов – Грибанов и др., 2012). Таким образом, снижение активности глутатион S – трансферазы у E. verrucosus в условиях повышения температуры, может быть связано как с инактивацией фермента вследствие высокой физиологической температуры, так и по причине высокой интенсивности процессов переокисления липидов и роста концентрации его продуктов.
Общее снижение активности глутатион S-трансферазы и пероксидазы у исследованных амфипод при понижении температуры, как и ранее показанное снижение содержания БТШ70, вероятно, свидетельствует о замедлении метаболизма или перераспределение энергии на более значимые процессы.
В заключение, возвращаясь к экологическим характеристикам вида, следует отметить связь полученных данных с ранее опубликованными материалами оценки температурного преферендума вида E. verrucosus (Timofeyev, Shatilina, 2007). Из приведенных в работе данных хорошо видно, что наиболее выраженные изменения показателей клеточного метаболизма у амфипод E. verrucosus наступают при превышении температурного порога в 11°С или при снижении температуры ниже 4оС. Установленный температурный диапазон 4-11°С отражает зону стабильности показателей клеточного метаболизма у данного вида и, что самое важное, этот диапазон совпадает с диапазоном наиболее предпочитаемых видом температур в условиях температурного градиента, выявленным ранее нами в поведенческих экспериментах.
Таким образом, в работе впервые для байкальских амфипод продемонстрировано, что зона стабильности показателей клеточного метаболизма у E. verrucosus тесно связана с его термопреферентной зоной и, вероятно, отражает границы температурного оптимума данного вида.
Авторский коллектив выражает огромную благодарность проф. И.Соколовой за ценные консультации, а также к.б.н. Павличенко В. В. и к.б.н. Протопоповой М. В. за помощь в проведении настоящего исследования.
Работа выполнена при частичной финансовой поддержке грантов РФФИ (12-04-31767 мол_а, 12-04-90039 Бел_а, 12-04-98062-р_сибирь_а, 11-04-00174-а, 11-04-91321-СИГ_а), грантов
Президента РФ (МК-5466.2012.4, МД-2063.2012.4, МК-4772.2011.4), ФЦП «Научные и научно-педагогические кадры инновационной России», а также при поддержке НОЦ Байкал ФГБОУ ВПО «ИГУ» и программы DAAD М. Ломоносов 2012-2013 (10.51.2011).
Список литературы Определение озерного байкальского эндемического амфиполя Eulimnogammarus verrucosus (Gerstf., 1858). Термический оптимизм ограничивается изменениями его метаболических маркеров
- Farnsworth P.N., Singh K. (2000) Self-complementary motifs (SCM) in alpha-crystallin small heat shock proteins. FEBS Latters, 482. pp.175-179.
- Fridovich I. (1998) Oxygen toxicity: a radical explanation. J. Exp Biol, 201, 8, pp.1203-1209.
- Geret F., Serafim A., Barreira L. et al. (2002) Effect of cadmium on antioxidant enzyme activities and lipid peroxidation in the gills of the clam Ruditapes decussates. Biomarkers, 7, 3, pp. 242-256
- Ito H., Kamei K., Iwamoto I. et al. (2003) Hsp27 suppresses the formation of inclusion bodies induced by expression of 120Ga aB-crystallin, a cause of desminrelated myopathy. CMLS, Cell. Mol. Life Sci, 60. p.1217-1223.
- Kamaltynov R. M. (1999) On the higher classification of lake baikal amphipods. Crustaceana, 72, 8, pp. 933 -944.
- Kawall H. G., Torres J. J., Sidell B. D. et al. (2002) Metabolic cold adaptation in Antarctic fishes: evidence from enzymatic acrivities od brain. Marine biology, 140, pp. 279-286
- Motohashi K., Watanabe Y., Yohda M. et al. (1999) Heat-inactivated proteins are rescued by the DnaKzJ-GrpE set and ClpB chaperones. Proc. Natl. Acad. Sci. USA, 96, pp. 7184-7189
- Pörtner H. O., Farrel A. P. (2008) Ecology. Physiology and climate change. Science, 322, pp. 690-692.
- Pörtner H. O., Knust R. (2007) Climate change affects marine fishes through the oxygen limitation of thermal tolerance. Science, 315, pp. 95-97.
- Selvakumar S., Geraldine P. (2003) Thermal modulation of pyruvate metabolism in the freshwater prawn Macrobrachium malcolmsonii: the role of lactate dehydrogenase. Fish Physiology and Biochemistry, 29, 149-157.
- Sokolova I. M. (2004) Cadmium effects on mitochondrial function are enhanced by elevated temperatures in a marine poikilotherm Crassostrea virginica Gmelin (Bivalvia: Ostreidae). J. Exp. Biol, 207, pp. 2639-2648.
- Stillman J. H., Somero G. N. (2001) A comparative analisys od the evolutionary patterning and mechanistic bases of lactate dehydrogenase thermal stability in porcelain crabs, genus petrolisthes. J. Exp. Biol, 204, pp. 767-776.
- Timofeyev M. (2006) Antioxidant enzyme activity in endemic Baikalean versus Palaearctic amphipods: Tagma-and size-related changes. Comparative biochemistry and physiology B: Biochemistry and Molecular Biology, 143, 3, pp 302-308.
- Timofeyev M. A., Shatilina Z. M. (2007) Different preference reactions of three Lake Baikal endemic amphipods to temperature and oxygen are correlated with symbiotic life. Crustaceana, 80, 2, pp.129-138.
- Timofeyev M.A., Steinberg C (2006) Comparative study of the antioxidant response to natural organic matter (NOM) exposure in three Baikalean amphipod species from contrasting habitats. Comparative biochemistry and physiology B: Biochemistry and Molecular Biology, 145, 197-203.
- Willmer P., Stone G., Johnson I. (2000) Environmental physiology of animals. Oxford. Blackwell Science, 644p.
- Аксенов -Грибанов Д. В., Шатилина Ж. М., Лубяга Ю. А. и др. (2012) Температурно -зависимые изменения показателей перекисного окисления липидов у эндемичных байкальских и палеарктических амфипод и гастропод. Известия Иркутского государственного университета. Серия Биология. Экология, (в печати).
- Базикалова А. Я. (1945) Амфиподы оз. Байкал. Труды Байкальской Лимнологической станции АН СССР, 11, 440 с.
- Бекман М. Ю., Деньгина Р. С. (1969) Население бентали и кормовые ресурсы рыб Байкала. Биологическая продуктивность водоемов Сибири. М., с. 42-47.
- Беркин Н. С. И др. (2009) Байкаловедение. Иркутск: Изд-во Ирк. гос. ун-та, 291 с.
- Вейнберг И. В. (1995) Сообщества макрозообентоса каменистого пляжа озера Байкал: Дис. канд. биол. наук. Иркутск, 180 с.
- Вейнберг И. В., Камалтынов Р. М. (1998) Сообщества макрозообентоса каменистого пляжа озера Байкал. Зоологический журнал, 77, 3, с.259 -265.
- Гаврилов Г. Б. (1949) К вопросу о времени размножения амфипод и изопод оз. Байкал. Докл. АН СССР, LXIV, 5, с.739 -742.
- Кулинский В. И. (1999) Активные формы кислорода и оксидативная модификация макромолекул: польза, вред и защита, Соросовский образовательный журнал, 1, с. 2-7.
- Маянский Д. Н. Проблемы хронического воспаления в современной патофизиологии//Патол. физиология и эксперим. терапия -1994. -№2, с. 51-55.
- Меньщикова Е. Б., Ланкин В. З., Зенков Н. К. и др (2006) Окислительный стресс. Прооксиданты и антиоксиданты. М.: Слово, 553 с.
- Павличенко В. В., Шатилина Ж. М., Бедулина Д. С. И др (2009) Накопление лактата и белков теплового шока (БТШ) при остром температурном стрессе у байкальских термочувствительных Eulimnogammarus vittatus и Eulimnogammarus marituji (Crustacea, Amphipoda). Амурский зоологический журнал, 1, 3, с.190-196
- Тимофеев М. А. (2010) Экологические и физиологические аспекты адаптации к абиотическим факторам среды эндемичных байкальских и палеарктических амфипод: Дис. … Д-р. биол. Наук. ТГУ, Томск, 384с.
- Тимошкин О. А., Провиз В. И., Ситникова Т. Я., и др (2004) Аннотированный список фауны озера Байкал и его водосборного бассейна. Новосибирск: Наука, 1679 с.
- Хочачка П. Сомеро Дж. (1988) Биохимическая адаптация. М.: Мир, 568с.
