Определение пола моевки Rissa tridactyla по морфометрическим признакам

Автор: Бирина У.А., Бирина Г.А., Краснов Ю.В.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 118 т.9, 2000 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149096
IDR: 140149096
Sex determination in the black-legged kittiwake Rissa trudactyla through the measurements
Текст статьи Определение пола моевки Rissa tridactyla по морфометрическим признакам
-
У. А. Бирина^, Г.А.Бирина, Ю.В.Краснов2)
° Кафедра зоологии позвоночных, Биолого-почвенный факультет, Санкт-Петербургский университет, Университетская набережная, д. 7/9, Санкт-Петербург, 199034, Россия
-
2) Мурманский морской биологический институт Российской Академии наук, ул. Владимирская, д. 17, Мурманск, 183023, Россия
Поступила в редакцию 29 мая 1992
Определение пола моевки Rissa tridactyla на живых птицах по размерным характеристикам до настоящего времени было затруднено, поскольку литературных данных по этому вопросу имелось очень мало. Сведения по морфометрии, представленные в работах В.И.Придатко (1984) и К.АЮдина и Л.В.Фирсовой (1988), не могут использоваться для определения пола из-за своей нерепрезентативности.
Наблюдая за живыми птицами, пол моевок можно определить во время формирования пар при церемонии выпрашивания корма и при спаривании (Chardine, John 1986). Сообщений о том, можно ли определить пол моевок по поведению на других стадиях репродуктивного цикла, в литературе нет.
Всё это побудило авторов исследовать размеры моевок, населяющих о-в Харлов в Баренцевом море (Кандалакшский заповедник), с целью разработки метода, позволяющего определить пол пойманных птиц.
Материал был собран Ю. В. Красновым и обработан У .А. Бириной и Г. А. Бириной.
Материал и методика
Использованы измерения 219 самцов и 102 самок моевки, отстрелянных с целью изучения питания на о-ве Харлов в гнездовой сезон 1983, 1984, 1985 и 1987.
Измерялись следующие параметры: длина головы (L2(Mf длина клюва (Z^), длина крыла (LKp), длина хвоста ^Lxef длина среднего пальца с когтем (LcnK), длина цевки (Ьц), масса тела (М). Отмечали наличие или отсутствие когтя на четвёртом пальце. Длину крыла и хвоста измеряли линейкой с точностью до 1 мм, массу тела — с точностью до 10 г. Все остальные промеры выполняли штангенциркулем с точностью до 0.1 мм.
Для проверки способов определения пола на живых птицах в 1989 была выбрана группа гнёзд на одном из базаров. Все гнёзда были нанесены на план местности и пронумерованы, на гнёздах была отловлена 61 особь.
При статистической обработке материала использовали критерии Вилкок-сона, со , дискриминантным анализ.
Результаты и обсуждение
При проверке выборок, характеризующих размеры самцов и самок моевки на нормальность с помощью критерия о2 оказалось, что большинство их распределений нормальному закону распределения не подчиняется (табл. 1). Поэтому для сравнения параметров использовали свободный от вида распределения критерий Вилкоксона. Итоги приведены в таблице 2 (массу тела брали по всем годам сразу, т.к. характеризовали её в целом). Как показывают результаты сравнения выборок (табл. 2), для всех параметров, кроме наличия когтя на 4-м пальце, гипотеза об однородности выборок отвергается, т.е. различия между всеми остальными параметрами самцов и самок значимы. По всем параметрам построены парные гистограммы. Их примеры даны на рисунках 1 и 2.
По данным В.И.Придатко (1984), размеры самцов и самок моевки не перекрываются: длина крыла самцов больше 330 мм, длина крыла самок меньше 320 мм; длина цевки самцов больше 39 мм, длина цевки самок меньше 37 мм. По нашим данным, несмотря на значимость различий между средними значениями, перекрытие происходит по всем параметрам. Поэтому определить пол по любому одному промеру возможно лишь в части случаев (см. рис. 1 и 2).
Мы попытались комбинировать разные параметры попарно на одной координатной плоскости. Сочетания “масса тела (конкретный год) — длина крыла” и “длина клюва — длина крыла” для определения пола использовать оказалось невозможным из-за большого перекрытия значений, поэтому эти графики даже не приводятся. Наиболее удобны комбинации “масса тела (1985 г.) — длина головы” (рис. 3) и “длина головы — длина крыла” (рис. 4).
По этому способу у 47 (78.3%) пойманных и измеренных особей на контрольном участке пол был определён верно. У 6 особей (10%) пол определён при сравнении с полом партнёра. В итоге пол удалось определить у 53 особей из 42 пар (88.3%) и у 7 особей (11.7%) пол определить не удалось. Таким образом, действенность метода определения пола одновременно по двум представленным графикам составила 78% плюс добавка в результате сравнения с полом партнёра, если он точно известен.
Интересно, что в гнезде № 2 обе птицы претендовали по размерам на самца — это пример сбоя метода.
Для более точного решения задачи отнесения каждой особи к самцам или самкам мы использовали метод дискриминантного анализа, реализованный в пакете программ STATGRAPHICS на ЭВМ IBM PC XT.
Предварительно были рассчитаны коэффициенты корреляции между всеми 7 признаками (табл. 3). Они оказались небольшими: максимальный коэффициент корреляции (0.5262) был между длиной клюва и длиной головы. Это допускает проведение дискриминантного анализа. Критическое значение абсолютной величины коэффициента корреляции, выше которого он может считаться значимым для объёма выборки (N = п\ + и2 = 69 + 32 = 101) равно 0.2.
Сначала мы провели дискриминантный анализ всех 7 признаков, различия по которым между выборками самцов и самок по критерию Вил- коксона оказались значимыми (табл. 2). Затем из этих признаков были выбраны 4, различия по которым между самцами и самками были большими, чем по другим признакам. Потом мы выделили всего 2 параметра с наибольшими различиями (масса тела и длина головы), которые уже были использованы нами ранее при определении пола моевок по графикам.
При построении дискриминантных функций для всех этих трёх случаев (по 7, 4 и 2 признакам) мы взяли выборки из 69 самцов и 32 самок, для которых были известны значения
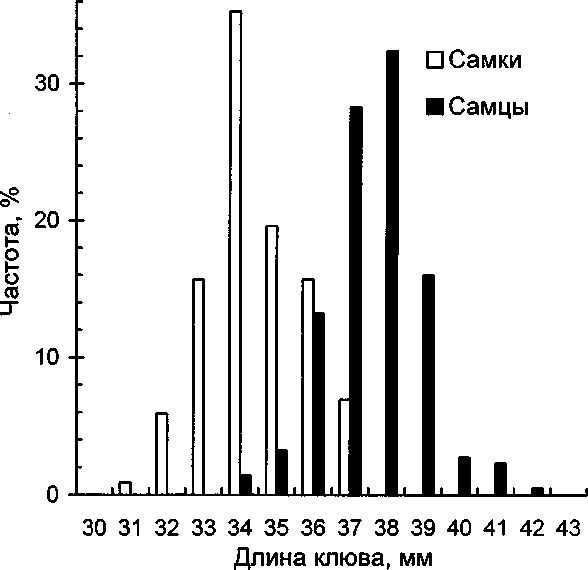
Рис. 1. Гистограммы распределений длины клюва у самцов и самок моевки.
Таблица 1. Статистические характеристики морфометрических признаков моевки Rissa tridactyla (число вверху относится к самцам, внизу- к самкам)
|
Параметр |
N |
S.D. |
Асимметрия |
Эксцесс |
Среднее |
min |
max |
Размах |
R* |
Нормальность |
|
Длина клюва, мм |
219 |
1.27 |
0.15 |
3.90 |
37.5 |
33.5 |
41.6 |
8.1 |
0.601 >Rxp |
- |
|
102 |
1.23 |
0.15 |
2.73 |
35.3 |
32.0 |
38.1 |
6.1 |
0.787> RKp |
- |
|
|
Длина головы, мм |
139 |
1.80 |
0.0053 |
2.84 |
94.1 |
89.9 |
99.5 |
9.6 |
0.343< RKp |
+ |
|
54 |
2.51 |
-1.0 |
5.47 |
88.9 |
80.2 |
93.6 |
13.4 |
0.718> Rkp |
- |
|
|
Длина цевки, мм |
219 |
1.29 |
-0.47 |
3.68 |
35.2 |
30.2 |
38.2 |
8.0 |
0.653> Rkp |
- |
|
102 |
1.09 |
-0.34 |
3.63 |
34.0 |
30.7 |
37.0 |
6.3 |
0.729> Rxp |
- |
|
|
Длина среднего |
149 |
2.06 |
-0.42 |
3.28 |
47.7 |
41.8 |
51.9 |
10.1 |
0.413< Rkp |
+ |
|
пальца с когтем, мм |
80 |
1.91 |
-0.25 |
2.70 |
45.8 |
41.0 |
49.7 |
8.7 |
0.265< Ркр |
+ |
|
Длина крыла, мм |
216 |
7.55 |
-0.27 |
2.81 |
319.4 |
299 |
339 |
40 |
1,035> Rkp |
- |
|
95 |
7.34 |
-.027 |
3.48 |
310.8 |
294 |
335 |
41 |
0.272< Rkp |
+ |
|
|
Длина хвоста, мм |
146 |
5.10 |
-0.18 |
2.41 |
135.3 |
125 |
147 |
22 |
1.095> Rkp |
- |
|
73 |
5.21 |
-0.41 |
2.77 |
129.9 |
118 |
141 |
23 |
0.767> Rkp |
- |
|
|
Масса тела, г |
219 |
41.11 |
-0.13 |
2.93 |
445.8 |
330 |
560 |
230 |
2.510> Ркр |
|
|
101 |
45.31 |
-0.27 |
3.89 |
416.6 |
300 |
540 |
240 |
1.694> Ркр |
- |
* RKp = 0.4614; а = 0.05.
Таблица 2. Результаты сравнения параметров самцов и самок моевки по критерию Вилкоксона
|
Параметр |
Эмпирическое значение U |
Границы доверительного интервала для теоретического значения U |
Значимость различий между самцами и самками |
|
Длина клюва,мм |
2398 |
9651.6 4- 12686.4 |
< 0.05 |
|
Длина головы, мм |
2645 |
3262.8 4- 4243.2 |
< 0.05 |
|
Длина цевки, мм |
5382 |
9651.6 4- 12686.4 |
< 0.05 |
|
Длина среднего |
|||
|
пальца с когтем, мм |
2799 |
5023.2 4- 6896.8 |
< 0.05 |
|
Длина крыла,мм |
4194.5 |
8828.4 4- 11691.6 |
< 0.05 |
|
Длина хвоста, мм |
2499.5 |
4462.6 4- 6195.4 |
< 0.05 |
|
Масса тела, г |
7767.5 |
9551.4 4- 12566.6 |
< 0.05 |
|
Наличие когтя |
|||
|
на 4-м пальце |
11245 |
9751.3 4- 12805.7 |
> 0.05 |
Таблица 3. Коэффициенты корреляции между признаками
-
1. Для случая 7 признаков:
-
2. Для случая 4 признаков:
-
3. Для случая 2 признаков:
= 0.17981^ + 0.76291^ - 0.0193ZKD +
+ 0.1369Л/ + 0.1472£„ + 0.0999Z,„ + 0.1496Z„
ZIW = 0.2193Z„ + 0.8155Z,_. + 0.0613ZKD + 0.150454/
Zim = 091¥)Lio. + 0.16454/. t J СМЛ
При получении значений Z^ > ZKp индивид относится к самцам, в противном случае — к самкам. Критическое значение Z в случае с 7 признаками равно 156.45; 4 признаками — 164.16; 2 признаками — 157.14.
Качество проведённой классификации оценивается обычно по двум характеристикам: расстоянию Махаланобиса 52, характеризующее обобщённое “расстояние” между группами: чем оно больше, тем лучше клас- сификация; или по величине X — статистики Уилкса, которая изменяется от 0 до 1 и оценивает величину остаточной дискриминации. Чем остаточная дискриминация меньше, тем лучше разделение. Эти характеристики использованы нами для проверки гипотезы об отсутствии разобщения выборок параметров самцов и самок с применением ^-распределения для расстояния Махаланобиса и распределения %2 для Х-ста-тистики Уилкса. Эмпирические оценки описанных выше характеристик для 7, 4 и 2 признаков, а также критические значение Fh х2 для соответствующих значений степеней свободы и уровня значимости 0.05 приведены в таблице 4.
Таблица 4. Характеристики качества классификации
|
Число признаков |
Xw |
х2 |
f |
х2 а - 0.05 |
52 |
fi |
^2 |
F3 |
Ft а = 0.05 |
|
7 |
0.3518 |
99.76 |
7 |
14.1 |
6.9875 |
7 |
93 |
20.62 |
2.1 |
|
4 |
0.3648 |
97.80 |
4 |
9.49 |
6.8765 |
4 |
96 |
36.82 |
2.5 |
|
2 |
0.3733 |
96.56 |
2 |
5.99 |
7.14 |
2 |
98 |
77.96 |
3.1 |
550 --
500 --
ГО
Ф
450 --
ГО о о го
400 --
350 --
80 85 90 95
Длина головы, мм
Рис. 3. Скаттер-диаграмма значений массы тела и длины головы для самцов и самок моевки
Полученные эмпирические значения F и %2 намного превышают критические табличные, что позволяет отвергнуть гипотезу об отсутствии разделения выборок и говорит о высоком его уровне. О высоком уровне разобщения свидетельствуют также данные отнесения особей моевки к самцам или самкам по результатам приведённых выше расчётов дискриминантных функций для каждой птицы известного пола. Так, по 7 признакам к самцам из 69 особей отнесено 65 (ошибка разделения 5.8%), а в группе самок из 32 особей правильно определены 30 (ошибка 6.25%). Для 4 признаков результаты следующие: из 69 самцов правильно классифицированы 66 (ошибка 4.35%), из 32 самок — 28 (ошибка 12.3%). В случае использования 2 признаков к самцам отнесены 66 особей из 69 (ошибка 4.35%), а к самкам — 29 из 32 (ошибка 9.4%).
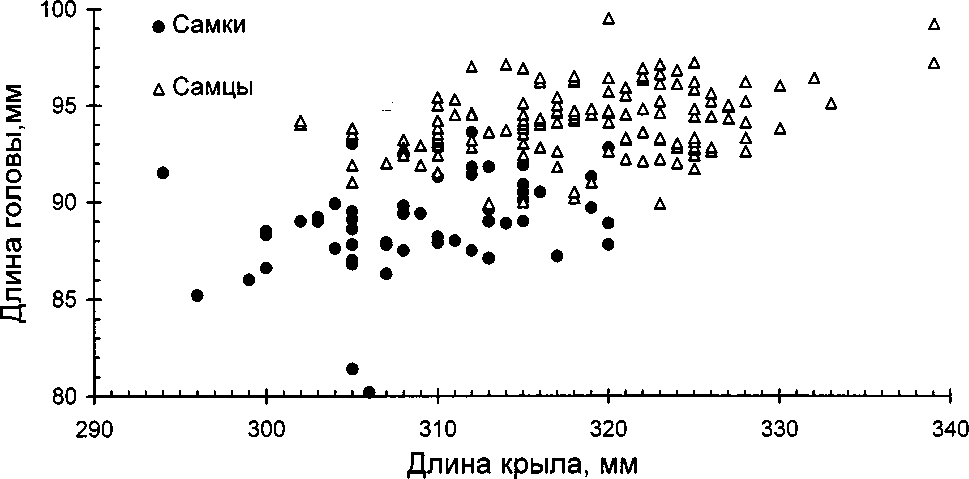
Рис.4. Скаттер-диаграмма значений длины головы и длины крыла для самцов и самок моевки
Для того, чтобы оценить степень различия дискриминантной способности всех трёх вариантов разделения, мы сравнили Х-статистики Уилкса и расстояния Махаланобиса 52, рассчитанные ранее по дискриминантным функциям. Проверка значимости различий этих характеристик проведена с помощью соответствующих критериев, использующих /'’-распределение (Энслейн и др. 1986). При сравнении статистик Уилкса наибольшее эмпирическое значение F3, равное 1.06, получено при сравнении дискриминации по 2 и 7 признакам при критическом табличном значении F = 9.55 (/1 = 2; f2 = 7, а = 0.05), что говорит об отсутствии достаточных оснований отвергнуть гипотезу об их равнозначности, т.е. свидетельствует о незначительности различия в качестве дискриминации моевок на самцов и самок при использовании 7 и 2 признаков. При сравнении Х-статистик для 4 и 7 признаков (см. табл. 4) также получено значение F3, равное 1.04, значительно ниже критического (критическое табличное значение F — 4.12, /1=4, / = 7, а = 0.05), что позволяет сделать такой же вывод. Аналогичная картина наблюдается и при сравнении эмпирических оценок расстояний Махаланобиса для всех трёх способов разделения. Например, максимальное различие в значениях 52 для 2 и 4 признаков — F3 = 1.11 при критическом табличном F= 3.09 (/i =2,/ = 96, а = 0.05).
Исходя из того, что характеристики качества разделения выборок (б2 и X) рассчитанных нами дискриминантных функций статистически равнозначны, для определения пола моевок можно использовать любую дискриминантную функцию из трёх, приведённых на стр. 17. Естественно, что легче пользоваться уравнением, включающим всего два параметра: длину головы и массу тела.
Мы определили пол моевок на контрольном участке по формуле и сравнили результаты с итогами определения пола по графикам 3 и 4. Из 59 птиц, у которых определили пол по формуле, у 5 (8.5%) результаты не совпали с полученными по графикам. Дважды получалось, что в гнезде оказывалось 2 самца (помимо гнезда № 2). Одна птица по всем параметрам графика была определена как самка, а по формуле — самец; две птицы, определённые по графикам как самки, по уравнению оказались самцами из-за того, что имели слишком большую для самок массу тела.
Из всего вышесказанного следует, что нельзя слепо доверяться полученной формуле, необходимо сопоставлять результаты расчётов с графиками и сравнивать пол птицы с полом партнёра, когда отловлена пара из одного гнезда. Кстати, два самца в гнезде № 2 в результате проведённых расчётов так и остались определёнными как самцы, что говорит о том, что встречаются настолько крупные самки, что их невозможно узнать по морфометрическим признакам.
Выводы
Пол моевок можно определять по любому из трёх уравнений, представленных на стр. 17 и имеющих равнозначную дискриминантную способность. Необходимо сопоставлять результаты расчётов с определением пола по графикам 3 и 4, а также сравнивать пол птицы с полом партнёра, если последний известен. Только при проведении всех сопоставлений ошибка определения будет минимальной.
Необходимо искать другие прижизненнные способы определения пола моевок, поскольку в некоторых редких случаях могут быть встречены настолько крупные самки, что по их промерам правильно определить их пол невозможно
