Опыт эколого-популяционного анализа микрогемипопуляций партенит трематод

Автор: Ишкулов Дмитрий Геннадиевич, Ишкулова Татьяна Геннадьевна
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Краткое сообщение
Статья в выпуске: 2 т.22, 2019 года.
Бесплатный доступ
В ходе проведения комплексных паразитологических исследований выполнен анализ популяционной структуры плоских паразитических червей вида Himasthla larina (Trematoda: Echinostomatidae). Показано, что формирование численной и размерной структуры популяции представителей данного вида напрямую связано с размерами и массой их первого промежуточного хозяина - моллюсков рода Littorina. Установлено, что при незначительных значениях показателя интенсивности инвазии увеличение размера паразитических организмов происходит одновременно с увеличением их численности независимо от массы и размера моллюска. С возрастанием данного показателя увеличение их размеров может идти только параллельно с увеличением массы хозяина, в дальнейшем мы наблюдаем обратную связь между размерами и количеством паразитов. На основании проведенных исследований сделан вывод, что формирование популяций партенит трематод подчиняется тем же законам, что и формирование популяций свободноживущих организмов.
Паразит, популяция
Короткий адрес: https://sciup.org/142220020
IDR: 142220020 | УДК: 576.895.122 | DOI: 10.21443/1560-9278-2019-22-2-288-291
The ecological analysis of the microhemipopulations of trematode parthenites: an experience
The paper analyzes the population structure of parasitic flatworms of the Himasthla larina species (Trematoda: Echinostomatidae). Abundance and size structure of their population directly relate to the size and weight of their first intermediate hosts, mollusks of the genus Littorina. When the invasion intensity is insignificant, growth of the size of parasitic organisms occurs simultaneously with the increase in their abundance regardless of the weight and size of the mollusk. If the weight and size of the host increase, the size of flatworms can grow only simultaneously with an increase of the host's weight. Further on inverse relationship between the size and abundance of parasites is observed. As a result of this study we may conclude that the formation of populations of trematode parthenites is subject to the same laws as the formation of populations of free-living organisms.
Текст научной статьи Опыт эколого-популяционного анализа микрогемипопуляций партенит трематод
*Мурманский морской биологический институт, г. Мурманск, Россия; ORCID: , e-mail:
В популяционной биологии экологический подход означает выяснение связей популяции в качестве целостной системы с внешними (по отношению к популяции) факторами – как биотическими, так и абиотическими (Яблоков, 1987). Однако при исследовании природных популяций мы зачастую сталкиваемся с тем, что даже численность популяции только в исключительно редких случаях удается установить напрямую – простым подсчетом. То же касается и изучения других параметров популяций: демографическая структура, скорость популяционного роста, плотность, биомасса и т. д.
Большинство подобных проблем снимается, если объектом исследований являются либо искусственная популяция (например микроорганизмов) in vitro, либо микропопуляция. По В. Н. Беклемишеву (1970), популяциям и субпопуляциям свойственен один общий признак – местом их обитания являются строго определенные биотопы. В отличие от них микропопуляции занимают микробиотопы. Согласно В. Н. Беклемишеву (1970) микробиотопы представляют собой "не части земной поверхности, а находящиеся на этой поверхности или вблизи нее. Это могут быть помет и трупы животных, норы, гнезда, а в случае рассмотрения паразито-хозяинных систем – организмы хозяина". Микропопуляции, как и популяции, способны к самовоспроизводству.
Большинство паразитических организмов, в жизненном цикле которых присутствуют два и более хозяев, образуют гемипопуляции. Гемипопуляции формируются из особей отдельных фаз жизненного цикла. В случае если гемипопуляции способна к самовоспроизводству, то для ее описания предлагается термин "микрогемипопуляция". Цель нашей работы – показать возможность определения зависимости формирования структуры микрогемипопуляций паразитических организмов от населяемого ими биотопа – хозяина.
Материалы и методы
Объектом исследований были выбраны представители вида Himasthla larina. Их партеногенетические личинки – редии – паразитируют на литоральных гастроподах рода Littorina (Ишкулов и др., 1998). Выбор именно этой паразито-хозяинной системы обусловлен следующими причинами:
-
1. Как показали данные предыдущих исследований, численность редий этого вида, паразитирующих в одном моллюске, может колебаться от нескольких десятков до нескольких сотен экземпляров.
-
2. Редии этой группы трематод являются гистиофагами, т. е. питаются непосредственно тканями моллюска, не оказывая на последнего патогенного влияния, приводящего к его смерти.
-
3. Зараженность моллюсков рода Littorina редиями химастлин не изменяет скорость роста организма-хозяина, в отличие от ряда других паразито-хозяинных систем (Дружков, 1990; Sousa, 1990).
Материал для исследований отбирали на побережье Кольского п-ова, в районе губы Ярнышная (Восточный Мурман, Баренцево море). Моллюсков L. saxatilis собирали со среднего горизонта литорали, с поверхности обнажающихся во время отлива камней. Паразитологическое обследование производилось по стандартным методикам (Галактионов и др., 1988). У моллюсков, зараженных редиями химастлин, определяли вес и среднюю длину паразитов, производили тотальный подсчет численности микрогемипопуляций.
Все собранные данные обработаны с помощью программы Golden Software Surfer 7.0. Итоги обработки представлены на рисунке.
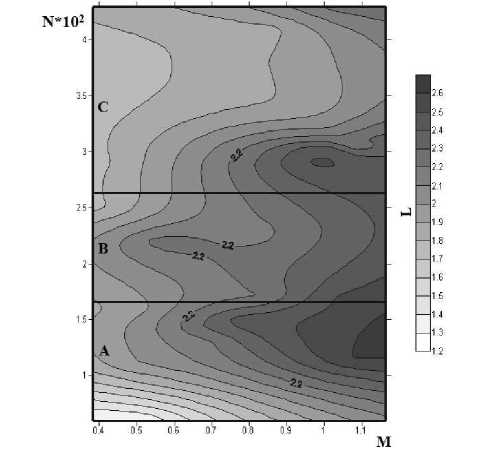
Рис. Зависимость между массой моллюска (М, г), количеством редий (^) и их средней длиной (L, мм) (пояснения в тексте)
Fig. The relationship between the mollusk weight (M, g), the number of redies (^), and their average length (L, mm) (explanations in the text)
Результаты и обсуждение
С точки зрения классической экологии для многих микропопуляций, и, прежде всего, для микропопуляций паразитических организмов, биотоп, в котором они обитают (микробиотоп), одновременно является и ресурсом.
Таким образом, для партеногенетических стадий трематод биотопом и одновременно ресурсом будет их первый промежуточный хозяин, в роли которого в подавляющем большинстве случаев выступает брюхоногий моллюск. Причем в тех случаях, когда паразитические организмы не оказывают на хозяина патогенного воздействия, приводящего к его гибели, речь идет о ресурсе возобновляемом. Следовательно, чтобы скорость использования ресурса не превысила скорость его возобновления, должны существовать механизмы, ее регулирующие. Таким образом, условия, при которых в моллюске может существовать микрогемипопуляция редий, описываются формулой (1):
Кпотр. < Квоз., (1)
где Кпотр. - скорость использования (потребления) ресурса; Квоз. - скорость его возобновления.
В свою очередь, скорость использования ресурса будет напрямую связана с количеством паразитов и их размерами (2):
Кпотр. = j(N; L), (2)
где N - численность, L - средний размер организмов в популяции. Понятно, что последние два показателя определяют общую биомассу популяции.
Следовательно, логично предположить, что при увеличении численности группировки паразита в организме хозяина должно происходить уменьшение их размеров, что в конечном итоге будет приводить к стабилизации общей биомассы микрогемипопуляции.
Если обратиться к рисунку, то на полученной нами модели можно выделить три области, определяющие условия формирования микрогемипопуляций. В микрогемипопуляциях, где число редий менее 130 экземпляров (область А), соматический рост происходит одновременно с увеличением численности и не зависит от размера моллюска. В случае когда в моллюске содержится от 150–250 редий (область В), увеличение их размера происходит только с увеличением массы хозяина. При дальнейшем количественном росте микрогемипопуляций химастлин (область С) наблюдается обратная связь между размером редий и их количеством.
Причинами такого разделения послужили следующие факторы. В популяциях первой области количество редий невелико, и насыщение микробиотопа не произошло. Следовательно, существует потенциал как для увеличения численности организмов, так и их размеров. Именно в этот момент происходит резкое нарастание биомассы паразитов в хозяине. Данный случай может быть описан формулой (3):
Кпотр. < Квоз. (3)
По мере увеличения числа редий за счет партеногенеза происходит насыщение биотопа; и их размер и, соответственно, биомасса микрогемипопуляции может возрастать только с увеличением массы моллюска, т. е. с увеличением емкости биотопа (область В).
При превышении пороговой численности в 250–300 редий в одном моллюске (область С) мы наблюдаем обратную зависимость между численностью и размерами паразитов. Именно благодаря этому происходит стабилизация биомассы паразитической микрогемипопуляции, которая приводит к снижению патогенного влияния на хозяина и позволяет не превысить скорость использования ресурса.
Заключение
Исследования показывают, что в паразито-хозяинных системах существуют механизмы, регулирующие общую биомассу паразитического компонента за счет обратной зависимости между численностью паразитов и их размерами, т. е. на популяционном уровне.
Таким образом, формирование микрогемипопуляций партенит трематод в целом подчиняется тем же законам, что и популяции свободноживущих организмов.
Работа выполнена в рамках фундаментальных научных исследований государственного задания ММБИ КНЦ РАН по следующим темам: "Паразиты в экосистемах северных морей: видовой состав, распределения, взаимоотношения с хозяевами" № в ГЗ 0228-2019-0002, № госрегистрации 01 2013 66844 "Особенности организации арктических планктонных сообществ в условиях современных климатических изменений" № в ГЗ 0228-2019-0003, № госрегистрации АААА-А-17-117052310083-5.
Список литературы Опыт эколого-популяционного анализа микрогемипопуляций партенит трематод
- Беклемишев В. Н. Биоценологические основы сравнительной паразитологии. М.: Наука, 1970. 502 с.
- Галактионов К. В., Марасаев С. Ф., Тимофеева С. В., Марасаева Е. Ф. Методы оценки паразитологической ситуации в прибрежье Баренцева моря: . Апатиты: Кол. фил. АН СССР, 1988. 46 с.
- Дружков Н. В. Статистический анализ размерной структуры популяции беломорской Hydrobia ulva (Pennant, 1777): распределение партенит трематод в популяции хозяина//Морфология и экология паразитов морских животных: . Апатиты, 1990. С. 52-61.
- Ишкулов Д. Г., Куклин В. В. К фауне химастлин Восточного Мурмана//Паразитология. 1998. Т. 32, вып. 1. С. 84-94.
- Яблоков А. В. Популяционная биология. М.: Высш. шк., 1987. 303 с.
- Sousa W. P. Spatial scale and the processes structuring a guild of larval trematode parasites//Parasite Communities: Patterns and Processes/eds.: G. Esch, A. Bush, J. Aho. London: Chapman & Hall, 1990. P. 41-67. DOI: https://doi.org/10.1007/978-94-009-0837-6.
