Организация ценопопуляций Salvia verticillata L. в Республике Марий Эл с позиций экогеографического подхода

Автор: Османова Г.О., Животовский Л.А.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 3 т.19, 2025 года.
Бесплатный доступ
Изучена онтогенетическая структура 10 ценопопуляций (ЦП) Salvia verticillata, произрастающих в Республике Марий Эл (РМЭ). Состояние ЦП S. verticillata неоднозначно. ЦП 1–5 нормальные, полночленные (кроме ЦП 2). Значительная доля участия в ЦП 1–5 особей прегенеративного и генеративного периодов свидетельствует о высокой семенной продуктивности S. verticillata и способности ЦП к самоподдержанию, и устойчивому развитию, но сильная антропогенная и зоогенная нагрузки (ЦП 6–10) могут стать губительными для численности этого вида. S. verticillata проявляет мезобионтность по всем климатическим и почвенным факторам (Itклим.= 0.55; ItLc = 0.56; Itпочв. = 0.53) и не выходит за рамки фундаментальной экологической ниши, но на территории республики S. verticillata не может реализовать все возможные потенции, находясь в узких диапазонах экологических параметров. Выделение экогеографических единиц (ЭГЕ) и экогеографических агрегаций (ЭГА) основано на анализе тематических географических карт (почв, рельефа) и данных фитоиндикационной оценки экотопов, полученных с помощью экологических шкал Д.Н. Цыганова. ЦП S. verticillata, расположенные территориально близко друг к другу и находящиеся в сходных почвенных и иных условиях, объединили в ЭГЕ, а их, согласно тем же принципам – в ЭГА. Установлена схожесть изученных ЦП S. verticillata: в Новоторъяльском районе РМЭ выделили 2 ЭГЕ, которые объединили в одну ЭГА, в Мари-Турекском – 3 ЭГЕ и 2 ЭГА. Данные проведенной работы позволят выявить границы будущих природоохранных территорий.
Salvia verticillata L., редкий вид, Республика Марий Эл, фитоиндикация, экологические шкалы, ценопопуляция, онтогенетическая структура, экогеографическая единица, экогеографическая агрегация, особо охраняемые природные территории
Короткий адрес: https://sciup.org/148331555
IDR: 148331555 | УДК: 574.34; 582.929.4 | DOI: 10.24412/2072-8816-2025-19-3-274-288
Organization of coenopopulations Salvia verticillata L. in the republic of Mari El from the position of ecogeographical approach
The ontogenetic structure of 10 coenopopulations (CP) of Salvia verticillata growing in the Mari El Republic (MER) was studied. The state of the S. verticillata CP is ambiguous. The CPs 1–5 are normal, full-fledged (except for CP 2). A significant proportion of participation in CP 1–5 of individuals of the pregenerative and generative periods indicates high seed productivity of S. verticillata and the ability of the CP to self-sustain and sustainable development, but strong anthropogenic and zoogenic loads (CP 6–10) can be detrimental to the population of this species. S. verticillata exhibits mesobiontism in all climatic and soil factors (Itclim. = 0.55; ItLc = 0.56; Itsoil = 0.53) and does not go beyond the fundamental ecological niche, but on the territory of the republic S. verticillata cannot realize all possible potentials, being in narrow ranges of ecological parameters. The allocation of ecogeographic units (EGU) and ecogeographic aggregations (EGA) is based on the analysis of thematic geographical maps (soils, relief) and data on phytoindication assessment of ecotopes obtained using D.N. Tsyganov's ecological scales. The CPs of S. verticillata, located territorially close to each other and located in similar soil and other conditions, were combined into EGU, and they, according to the same principles, into EGA. The similarity of the studied CPs of S. verticillata was established: in the Novotoryalsky district of the Republic of Mari El, 2 EGE were identified, which were combined into one EGA, in Mari-Tureksky – 3 EGE and 2 EGA. The data from the work will allow us to identify the boundaries of future nature conservation areas.
Текст научной статьи Организация ценопопуляций Salvia verticillata L. в Республике Марий Эл с позиций экогеографического подхода
Сегодня губительные для большинства видов изменения среды во многом обусловлены хозяйственной деятельностью человека (распашка земель, выпас скота, осушение болот, ведение строительных работ, интродукция видов и др.), а также браконьерством и бесконтрольным использованием ресурсных видов. Некоторые популяции, находящиеся на краю ареала и имеющие пониженные адаптационные возможности, гибнут, будучи не в состоянии приспособиться к меняющимся условиям среды и не выдержав конкуренции со стороны других видов. Поэтому важно найти подходы к районированию популяций (Zhivotovsky, Osmanova, 2020).
Границы популяций определяются двумя основными факторами: условиями среды и пределами генных потоков. Природная популяция (многие поколения, обитающие в одном месте), адаптируется к его условиям и благодаря внутренним генным потокам формирует свои 274
уникальные особенности, общие для особей этой популяции. Условия среды – главный, определяющий фактор в жизни популяции. Ее фенотипический облик и генетический профиль – эволюционные производные этих условий. Изменение среды обитания – важнейшая причина исчезновения популяций и видов.
Ранее (Zhivotovsky, 2016, 2017; Zhivotovsky, Osmanova, 2018, 2019; Osmanova et al., 2019) было предложено использовать в целях управления природными биологическими ресурсами (промысла, воспроизводства, охраны) экогеографические единицы (ЭГЕ), каждая из которых представляет собой природное тело, скомпонованное из эколого-географических характеристик ареала и эколого-биологических признаков исследуемых популяций.
Шалфей мутовчатый ( Salvia verticillata L.) – евро-западносибирско-древнесредиземноморский степной вид (Abramov, 2000). С 2013 г. S. verticillata включен в Красную книгу РМЭ (Red…, 2013) со статусом 3 (редкий вид), в Красные Книги Пензенской, Калужской и Кировской областей (Red…, 2013, 2014, 2015). В России S. verticillata распространен в большей части на Кавказе и Алтае, в нечерноземных областях как заносное, встречается на обнажениях известняков и мергелей, по обочинам железных дорог (Gubanov et al., 2004). В Республике Марий Эл S. verticillata произрастает в Волжском, Мари-Турекском, Моркинском и Новоторьяльском районах (Red…, 2023). Большинство местообитаний S. verticillata не входят в особо охраняемые природные территории (ООПТ), исключением являются ЦП Волжского района, находящиеся в национальном парке «Марий Чодра» (рис. 1).
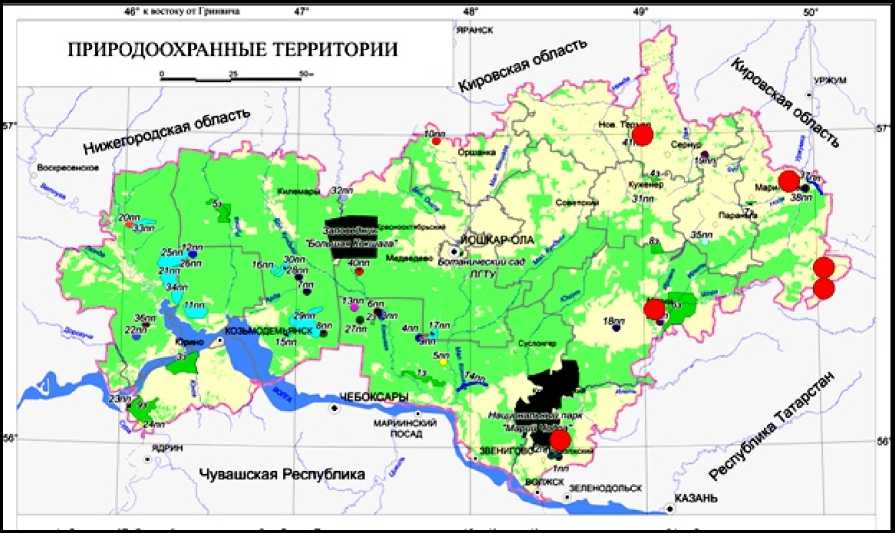
Рис. 1. Карта природоохранных территорий Республики Марий Эл (места произрастания S. verticillata отмечены красными кружками) (Ekologo-geograficheskii atlas)
Fig. 1. Map of protected areas of the Mari El Republic ( S. verticillata habitats are marked with red circles) (Ekologo-geograficheskii atlas)
Поэтому возникла проблема сохранения ЦП данного вида на территории Новоторъяльского, Моркинского и Мари-Турекского районов. Мониторинг объектов животного и растительного мира, занесенных в Красную книгу Республики Марий Эл (Red book, 2013, 2023) показал, что в местообитаниях S. verticillata в Новоторъяльском и Мари-Турекском районах происходит периодический выпас скота, что без сомнений, ставит под угрозу способность ЦП к самоподдержанию и восстановлению. Все это требует принятия мер по сохранению данного вида.
Цель работы – изучить онтогенетическую структуру ЦП S. verticillata с позиций экогеографического подхода и охарактеризовать его местообитания фитоиндикационным методом.
О БЪЕКТЫ И МЕТОДЫ
Шалфей мутовчатый – многолетнее травянистое растение семейства Lamiaceae (Gybanov et al., 2004). Жизненная форма S. verticillata разными авторами трактуется неоднозначно. И.Г. Серебряков (Serebryakov, 1964) определяет S. verticillata как длиннокорневищный травянистый поликарпик. Е.М. Олейникова (Oleynikova, 2012) и Н.П. Мехтиева с соавторами (Mehtieva et al., 2016) описали этот вид как стержнекорневое растение. Анализ литературных источников позволяет заключить, что данный вид проявляет морфологическую пластичность подземных органов в зависимости от типа почвы и её физико-химических свойств – гранулометрического состава, плотности, влажности и т.д. Следует отметить, что у генеративных растений стержнекорневой биоморфы, начиная со старого генеративного, а иногда и средневозрастного генеративного состояний, в корневой системе наблюдаются некротические процессы. Главный корень начинает шнуровидно закручиваться, а затем происходит его вертикальное расщепление. Такое поведение главного корня описано разными авторами (Nukhimovsky, 2002; Kozyreva, Osmanova, 2010; Osmanova, 2009; 2014) у растений разных биоморф и связано с недостаточным увлажнением почвы и высоким содержанием солей кальция. Поэтому, при выкопке такие растения распадаются в руках. Таким образом, происходит партикуляция особей, которая тоже является одним из способов вегетативного размножения. Видимого омоложения партикул не наблюдается. На дерновоподзолистых почвах S. verticillata может быть и длиннокорневищным травянистым поликарпиком, гемикриптофитом (Bakin et al., 2000).
По биологической классификации Х. Раункиера (Raunkiaer, 1934) S. verticillata – гемикриптофит, по отношению к водному фактору – мезоксерофит (Goryshina, 1979), по отношению к почвенным условиям – петрофит, гемерофоб, автохор (Tarasova, 2007).
Шалфей мутовчатый – полиморфный вид, часто можно видеть различия в характере зубчатости края листа, степени опушения листьев – от голых с рассеянными волосками до густо, войлочно-опушенных до бархатистых с обеих сторон (Flora, 1954). Основной способ самоподдержания S. verticillata – семенной, а если учесть, что он может быть представлен длиннокорневищной биоморфой, то и вегетативный, с помощью корневищ.
В работе использовали фитоиндикационный, популяционно-онтогенетический и картографический методы. Работа состояла из нескольких этапов. На 1 этапе исследования провели геоботанические описания экотопов с использованием фитоиндикационного метода по экологическим шкалам Д.Н. Цыганова (Tsyganov, 1983). Флористические списки обработали по программе ECOSCALE WIN методом средневзвешенной середины интервала (Zubkova et al., 2008). С использованием подходов Л.А. Жуковой (Zhukova, 2004) рассчитали потенциальную (PEV) и реализованную (REV) экологические валентности. Эффективность освоения экологического пространства ЦП оценили с помощью коэффициента экологической эффективности (Kec.eff.). Для характеристики отношения конкретного вида к совокупному воздействию нескольких факторов рассчитали индекс толерантности (I t ) (Ecological scale …, 2010).
Второй этап заключался в изучении онтогенетической структуры ЦП S. verticillata. Нами изучено 10 ЦП S. verticillata , четыре из которых располагались на территории Новоторъяльского района, остальные шесть – в Мари-Турекском. Для характеристики онтогенетической структуры ЦП S. verticillata рассчитаны индексы возрастности (∆) (Uranov, 1975) и эффективности (ω) (Zhivotovsky, 2001). Типы спектров определены по Л.Б. Заугольновой (Zaugolnova, 1994). Тип онтогенетической структуры ЦП определяли по классификации «дельта-омега» (Zhivotovsky, 2001).
Третий этап заключался в применении экогеографического подхода, который подразумевает выделение экогеографических единиц (ЭГЕ) и экогеографических агрегаций (ЭГА) для определения границ будущих природоохранных территорий необходимых для изучения популяционной структуры S. verticillata . Согласно этому подходу ЦП S. verticillata , произрастающие в относительно однородной среде и связанные друг с другом потенциальными генными потоками, объединяются в экогеографическую единицу. Границей ЭГЕ служит дальность генного потока, то есть условный радиус разноса пыльцы и диаспор. Для определения однородности среды, ЦП S. verticillata охарактеризовали по экологическим 276
шкалам Д.Н. Цыганова (Tsyganov, 1983). Далее с использованием картографического метода провели анализ тематических карт Республики Марий Эл (почвенной, рельефа, карты транспортных путей и особо охраняемых природных территорий РМЭ), на которые нанесли координаты мест произрастания объекта исследования.
Р ЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
Все изученные ЦП S. verticillata располагались на открытых склонах с известняковомергелистыми обнажениями, преимущественно южных и юго-западных экспозиций. Краткие характеристики сообществ, в которых закладывались пробные площадки с участием S. verticillata представлены ниже.
-
1. Верониково-землянично-шалфейная ассоциация. ЦП 1 располагается на склоне южной и юго-западной экспозиции к долине р. Немда и насчитывает 67 особей S. verticillata , среди которых доминируют особи прегенеративного периода, особенно растения ювенильного онтогенетического состояния. Крутизна склона 35-40˚. Выше пробной площади, на вершине склона молодой разреженный 10 летний сосняк. Ниже, в долине реки, высоко-злаковый луг из ежи сборной ( Dactylis glomerata L.) и лисохвоста лугового ( Alopecurus pratensis L.). К югу и к северу по склону расположен сосново-еловый корневищноосоково-разнотравный лес. Общее проективное покрытие (ОПП) – 90 %. Видовая насыщенность – 41 вид. Доминантные виды: S.verticillata , вероника широколистная ( Veronica teucrium L . ), земляника зеленая ( Fragaria viridis (Duchesne) Weston), змееголовник тимьяноцветковый ( Dracocephalum thymiflorum L.), подмаренник северный ( Galium boreale L.). Древостой отсутствует. На склоне есть участки с преобладанием в травяном покрове вейник наземный ( Calamagrostis epigeios (L.) Roth), который образует здесь локус. Почва перегнойно-карбонатная мощностью 10–15 см с выходами мергеля и известняка на поверхность. Ниже мергелистый горизонт.
-
2. Котовниково-шалфейно-девясиловая ассоциация. ЦП 2 располагается на склоне южной и юго-западной экспозиции к долине р. Немда. На площади в 12 м2 произрастает 205 растений S. verticillata разных онтогенетических состояний, но с явным преимуществом виргинильных особей. Генеративные растения единичны. Крутизна склона 35–40˚. Из-за отсутствия сенокошения луг начинает зарастать соснами и елями, которые местами достигают высоты до 5 м. Ниже, в долине реки злаковый луг из D. glomerata и A. pratensis . К югу и к северу по склону сосново-еловый корневищноосоково-разнотравный лес. Почва перегнойно-карбонатная мощностью 10-15 см. Ниже мергелистый горизонт. Местами мергель и известняк выходит на поверхность и занимает до 10% поверхности луга. ОПП травостоя 90 %. Видовая насыщенность составляет 61 вид.
-
3. Васильково-землянично-клеверная ассоциация. ЦП 3 находится на склоне левого берега р. Немда. Крутизна склона 40–45˚. На пробной площади насчитали108 растений преимущественно ювенильных и молодых генеративных. Выше площадки на вершине склона находится зарастающее разнотравьем поле. Ниже, в долине реки, расположен бодяково-васильково-злаковый луг (частично косимый). Почва суглинистая делювиальная. На склоне начинает возобновляться лес. Антропогенного воздействия нет. ОПП травостоя 95–99%. Видовая насыщенность – 35 видов. Доминантные виды: клевер средний ( Trifolium medium L.), F. viridis , василек луговой ( Centaurea jacea L.) и шероховатый ( С. scabiosa L.).
-
4. Подмаренниково-землянично-нивянниковая ассоциация. ЦП 4 находится на верхней части склона южной экспозиции к долине ручья. На пробной площади 10 м2 произрастает 255 растений S. verticillata практически всех онтогенетических состояний. Много молодых вегетирующих и средневозрастных генеративных растений. Крутизна склона 35–40˚. Ниже по склону – мергелистый обрыв с углом склона 60˚. Древостой отсутствует, но есть единичные экземпляры можжевельника обыкновенного ( Juniperus communis L.), ОПП травостоя 75%. Доминанты растительного сообщества: льнянка обыкновенная ( Linaria vulgaris Mill.), F. viridis , подмаренник мягкий ( Galium mollugo L.), тысячелистник обыкновенный ( Achillea millefolium L.). Видовая насыщенность – 37 видов. Луг используется как пастбище для домашних животных.
-
5. В ассоциации молодого сосняка с клёном коротконожково-астрагаловым на средней части крутого склона – известняково-мергелисто-суглинистого обнажения к реке Арборка расположена ЦП 5 . Это наиболее многочисленная (447 особей на 30 м2) и процветающая ценопопуляция. Обнажения свежие, есть ложбинки стока после дождя. Древостой отсутствует.
-
6. ЦП 6 расположена в молодом сосняке астрагаловом на мысе, врезающемся в воду, где заканчиваются известняково-мергелисто-суглинистые обнажения к реке Арборка. S. verticillata здесь представлен единичными генеративными особями, молодые растения обнаружены не были. Обнажения зарастают молодыми соснами. ОПП травостоя 55%. Видовая насыщенность – 21 вид.
-
7. В клеверо-полынно-васильковой ассоциации у подножья склона, вдоль дороги с северной стороны, поднимающейся к бывшей д. Куммер, расположена ЦП 7 . Пробная площадь, на которой растет S. verticillata сильно повреждена скотом. Большинство растений сильно объедены. Древостой отсутствует. ОПП травостоя – 75%. Доминанты растительного сообщества: полынь горькая ( Artemisia absinthium L . ), Melilotus albus Medik.), одуванчик лекарственный ( Taraxacum officinale F.H. Wigg . ), T. medium , C. scabiosa . Видовая насыщенность – 29 видов. Луг используется как пастбище для домашних животных.
-
8. Подмаренниково-землянично-шалфейная ассоциация с участием ЦП 8 расположена по мергелистому склону юго-западной экспозиции Кумерской горы. Подножья склона повреждены скотом – много сорных видов. Иногда весной склоны поджигают, поэтому встречаются голые обнажения суглинка, известняка, мергеля. Крутизна склона – 40º. Древостой отсутствует. Травостой местами с проективным покрытием (ПП) 75%. Видовая насыщенность – 21 вид.
-
9. В разнотравно-лабазниково-клеверной ассоциации по мергелистому склону юговосточной экспозиции лощины, идущей со стороны поля к реке Ноля, в 200 м к западу от края посадок сосны на склонах расположена ЦП 9 с единичными поврежденными вегетирующими растениями S. verticillata низкой жизненности. Древостой: отсутствует. Травяной покров с ПП – 95%. Видовая насыщенность – 28 видов.
-
10. ЦП 10 находится в подмаренниково-землянично-шалфейной ассоциации по мергелистому склону южной экспозиции к р. Ноля. В ЦП отмечено 3 генеративные особи S. verticillata и 10 поврежденных вегетирующих растений низкой жизненности. Древостой отсутствует. Травяной покров с ПП – 85%. Видовая насыщенность – 21 вид.
ОПП травостоя – 55%. Доминанты растительного сообщества: P. sylvestris , B. pinnatum , P. saxifraga . Видовая насыщенность – 39 видов.
В исследуемых сообществах нами были выявлены и другие виды растений, включенные в Красную книгу Республики Марий Эл (Red book, 2013, 2023): астрагал серповидный ( Astragalus falcatus L.), василек ложнопятнистый ( Centaurea pseudomaculosa Dobrocz.), живокость высокая ( Delphinium elatum L.), дремлик темно-красный ( Epipactis atrorubens (Hoffm. ex Bernh.) Besser), котовник венгерский ( Nepeta pannonica L.), осока горная ( Carex montana L.), остролодочник волосистый ( Oxytropis pilosa (L.) DС.), фиалка горная ( Viola montana L.). Василек ложнопятнистый восстановил численность, поэтому в 2023 г. исключен из списка редких видов (Red book, 2023).
Результаты обработки флористических списков показали, что в целом, абиотические условия местообитаний ЦП S. verticillata являются сходными и укладываются в диапазоны, приводимые Д.Н. Цыгановым (Tsyganov, 1983) для данного вида (табл. 1). ЦП S. verticillata произрастают на достаточно богатых (баллы от 7.7 до 8.3), но бедных азотом, слабокислых / нейтральных (от 7.0 до 9.0) почвах с лугово-степным / сухолесолуговым (9.97–11.17) слабо переменным / умеренно переменным увлажнением, предпочитают полуоткрытые пространства и светлые леса. Это подтверждается тем, что все ЦП S. verticillata располагаются на склонах холмов или понижениях к долинам рек, преимущественно южных и юго-западных экспозиций. Поскольку S. verticillata является преимущественно степным видом, фактор освещенности-затенения играет немаловажную роль в расселении и формировании устойчивых ЦП.
-
S. verticillata по омброклиматическому фактору, факторам солевого режима почвы и переменности увлажнения является гемистеновалентным видом (ГСВ), по факторам термоклиматической и криоклиматической шкалы, шкалы увлажнения почвы и освещенности-затенения – мезовалентным (МВ), по фактору богатства почв азотом – гемиэвривалентным (ГЭВ), а по факторам кислотности почв и континентальности климата – эвривалентным (ЭВ). Коэффициент экологической эффективности для климатических факторов варьирует от 13.70 (Kn) до 25.00% (Om); для почвенных факторов – от 10.42 (Hd) до 22.22% (fH). Наибольшее экологическое пространство освоено видом по фактору освещенности-затенения (Lc) – 46.43 %.
Оценка реализованных экологических валентностей (REV) по климатическим и почвенным шкалам выявила, что в условиях РМЭ ЦП S. verticillata в незначительной степени реализуют свои потенциальные возможности.
Таблица 1. Характеристика местообитаний S. verticillata по экологическим шкалам Д.Н. Цыганова
Table 1. Characteristics of habitats of S. verticillata according to ecological scales of D.N. Tsyganov
|
Диапазон экологических шкал (Цыганов, 1983) Range of ecological scales (Tsyganov, 1983) |
Экологическая позиция вида по шкале факторов (Цыганов, 1983) Ecological position of a species on a scale of factors (Tsyganov, 1983) |
Потенциальная экологическая валентность (PEV) Ecological position of a species on a scale of factors (Tsyganov, 1983) |
Реализованная экологическая валентность (REV) Realized Environmenta l Valence (REV) |
Коэффициент экологической эффективности (К ec.eff. , %) Ecological efficiency coefficient (К ec.eff. , %) |
Индекс толерантности (I t ) Tolerance index (I t ) |
|
Tm (1–17) |
5–13 |
0.53 МВ/MV |
0.09 |
16.98 |
It клим. = 0.55 МБ/MB |
|
Kn (1–15) |
3–13 |
0.73 ЭВ/EV |
0.10 |
13.70 |
|
|
Om (1–15) |
5–10 |
0.40 ГСВ/HSV |
0.10 |
25.00 |
|
|
Cr (1–15) |
6–13 |
0.53 МВ/MV |
0.10 |
18.87 |
|
|
Lс (1–9) |
1–5 |
0.56 МВ/MV |
0.26 |
46.43 |
It Lc = 0.56 МБ/MB |
|
Hd (1–23) |
3–13 |
0.48 МВ/MV |
0.05 |
10.42 |
It почв. = 0.53 МБ/MB |
|
Tr (1–19) |
5–11 |
0.37 ГСВ/HSV |
0.08 |
21.62 |
|
|
Rc (1–13) |
5–13 |
0.69 ЭВ/EV |
0.15 |
21.74 |
|
|
Nt (1–11) |
4–10 |
0.64 ГЭВ/HEV |
0.1 |
15.63 |
|
|
fH (1–11) |
5–9 |
0.45 ГСВ/HSV |
0.1 |
22.22 |
Примечание. Экологические шкалы: Tm – термоклиматическая, Kn – континентальности климата, Om – омброклиматическая аридности-гумидности, Cr – криоклиматическая, Hd – увлажнения почвы, Tr – солевого режима почв, Nt – богатства почв азотом, Rc – кислотности почв, Fh – переменности увлажнения, Lc – освещенности-затенения. Экологическая валентность: ГСВ – гемистеновалентная; МВ – мезовалентная; ГЭВ – гемиэвривалентная; ЭВ – эвривалентная. Группы толерантности: МБ – мезобионтная.
Note. Ecological scales: Tm – thermoclimatic, Kn – continentality of climate, Om – ombroclimatic aridity-humidity, Cr – cryoclimatic, Hd – soil moisture, Tr – soil salt regime, Nt – soil nitrogen richness, Rc – soil acidity, Fh – moisture variability, Lc – illumination-shading. Ecological valence: HSV – hemisthenovalent; MV – mesovalent; HEV – hemieurivalent; EV – euryvalent. Tolerance groups: MB – mesobiont.
Максимальное значение REV наблюдается по фактору освещенности-затенения (0.26). По другим факторам значения REV варьируют от 0.08 до 0.26, что в несколько раз ниже значений PEV. Минимальное значение REV отмечено по фактору увлажнения почвы. Судя по всему, узкий реализованный диапазон по данному фактору связан с тем, что S. verticillata, являясь степным и гемистеновалентным по этому фактору видом, приурочен к сухостепному увлажнению, а в данных местообитаниях выявлено лугово-степное/сухолесолуговое увлажнение. Значения потенциальной и реализованной экологической валентности шалфея мутовчатого графически показано на диаграмме (рис. 2).
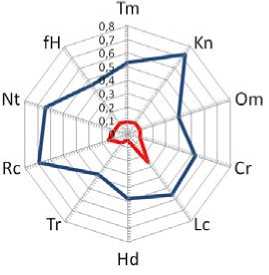
Рис. 2. Экологические валентности ценопопуляций S. verticillata : синяя линия – потенциальная (PEV), красная линия – реализованная (REV)
Fig. 2. Ecological valences of S. verticillata coenopopulations: blue line – potential (PEV), red line – realized (REV)
Диаграмма наглядно иллюстрирует – видом охвачен незначительный диапазон амплитуд по всем экологическим факторам. Экологические позиции изученных ЦП S. verticillata полностью укладываются в рамки возможных экологических потенций (Sinushkina, Osmanova, 2021).
Изучение популяционной структуры и организации ЦП растений, особенно редких и эндемичных видов, является необходимым условием их охраны и сохранения общего биоразнообразия природы. Исследование 10 ассоциаций с участием S. verticillata показало, что численность растений этого вида в ЦП разная, так же, как и онтогенетический состав. Анализ онтогенетических спектров и демографических показателей мы провели для ЦП 1–5 S. Verticillata . К сожалению, провести подобную работу в ЦП 6–10 не представилось возможным из-за небольшой численности и поврежденности шалфея. Следует отметить, что ЦП 6–10 S. verticillata находятся в угнетенном состоянии. Все 5 ЦП S. verticillata нормальные, ЦП 1, 3–5 – полночленные, ЦП 2 – неполночленная, т.к. отсутствуют особи сенильного состояния. В ЦП 1–3 онтогенетический спектр одновершинный левосмещенный с максимумом на ювенильной (ЦП 1), виргинильной (ЦП 2) и молодой генеративной (ЦП 3) группах. В ЦП 3 в прегенеративном периоде доминирует группа ювенильных растений, а в генеративном периоде – молодые генеративные растения (рис. 3).
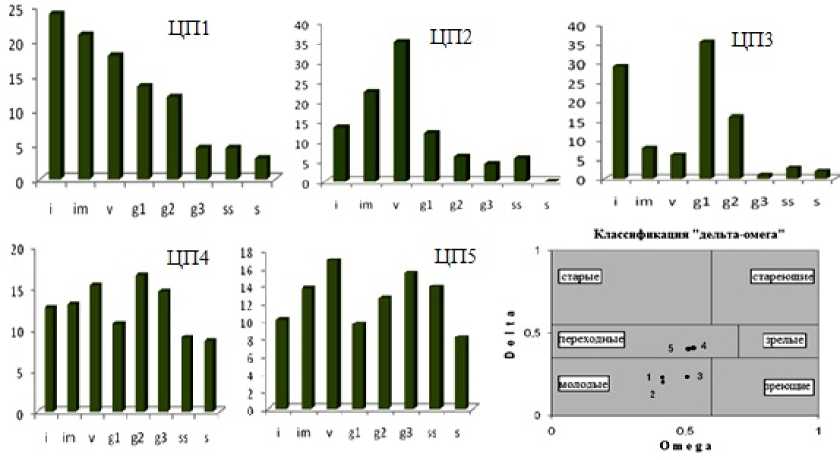
Рис. 3 – Онтогенетический спектр ЦП S. verticillata : по оси абсцисс – онтогенетические состояния: j – ювенильное, im – имматурное, v – виргинильное, g1 – молодое генеративное, g2 – зрелое генеративное, g3 – старое генеративное, ss – субсенильное, s – сенильное; по оси ординат – доля особей, в %
Fig. 3 – Ontogenetic spectrum of the CP of S. verticillata : on the abscissa axis – ontogenetic states: j – juvenile, im – immature, v – virginile, g1 – young generative, g2 – mature generative, g3 – old generative, ss – subsenile, s – senile; on the ordinate axis – the proportion of individuals, in %
Во всех трех ЦП S. verticillata доля особей постгенеративного периода минимальна. Онтогенетический спектр ЦП 4 и 5 S. verticillata бимодальный. Процессы самоподдержания во всех ЦП идут активно, о чем свидетельствуют высокие значения индекса восстановления – I в > 1 (Zhukova, 1995).
Согласно классификации «дельта-омега» (рис. 3), основанной на использовании индекса возрастности (Uranov, 1975) и эффективности (Zhivotovsky, 2001), первые три ЦП S. verticillata относятся к молодым (ЦП 1 ω=0.414, ∆=0.231; ЦП 2 ω=0.416, ∆=0.203; ЦП 3 ω=0.507, ∆=0.234), четвертая и пятая – к переходным (ЦП 4 ω=0.512, ∆=0.405; ЦП 5 ω=0.529, ∆=0.410).
Для всестороннего изучения популяций необходимо сочетание традиционных ботаникогеографических и современных молекулярно-генетических методов.
С целью изучения организации ЦП S. verticillata был применен экогеографический подход, согласно которому, ЦП со схожими экологическими условиями и расположенные территориально близко друг к другу мы объединили в экогеографические единицы. Для определения однородности среды ЦП S. verticillata были охарактеризованы по экологическим шкалам Д.Н. Цыганова (Tsyganov, 1983), а также был проведен анализ тематических карт республики. Результаты экологической оценки местообитаний S. verticillata по шкалам Д.Н. Цыганова показали, что ЦП весьма сходны по экологическим параметрам. В целом, это территории открытых и полуоткрытых пространств, с суббореальным/неморальным термоклиматическим режимом, с умеренной зимой и сухолесолуговым увлажнением. Для оценки сходства или различия экологических параметров были проанализированы тематические карты почвы и рельефа республики (рис. 4, 5).
Анализ почвенной карты РМЭ (рис. 4) и литературных источников (Abramov, 1995) показал, что изученные ЦП S. verticillata располагаются на дерново-подзолистых суглинистых почвах с выходами мергеля и известняка.
Рельеф, в основном, также схож (рис. 5) – все местообитания S. verticillata располагаются на склоновых участках холмов или понижениях к долинам рек. В целом, территория Марийско-Вятского увала, на которой расположены местообитания S. verticillata , представляет собой возвышенную равнину, состоящую из целого ряда увалов, холмов, плато, разделенных между собой понижениями. Большая глубина врезания речной сети придаёт его поверхности большую пересечённость.
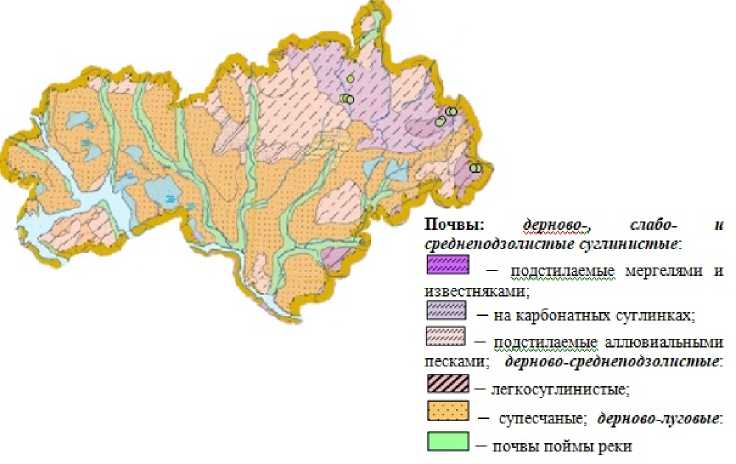
Рис. 4 . Местообитания изученных ценопопуляций S. verticillata, спроецированные на почвенную карту Республики Марий Эл (Ekologo-geograficheskii atlas) (отмечены зелеными кружками)
Fig. 4 . Habitats of the studied coenopopulations of S. verticillata , projected onto the soil map of the Mari El Republic (Ekologo-geograficheskii atlas) (marked with green circles)
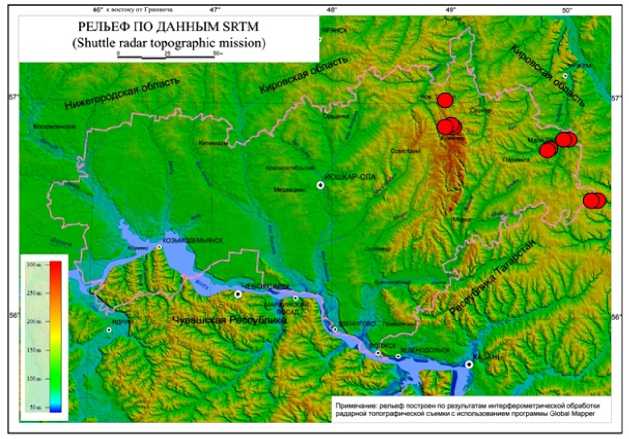
Рис. 5. Местообитания изученных ценопопуляций S. verticillata, спроецированные на карту рельефа Республики Марий Эл (отмечены красными кружками)
Fig. 5. Habitats of the studied coenopopulations of S. verticillata, projected onto the relief map of the Mari El Republic (marked with red circles)
Таким образом, экологическая характеристика местообитаний S. verticillata и анализ тематических карт позволяет сделать вывод о схожести ЦП по экологическим параметрам.
Ценопопуляции S. verticillata , расположенные территориально близко друг к другу и находящиеся в сходных почвенных и иных условиях, были объединены в экогеографические единицы (2 ЭГЕ в Новоторъяльском и 3 ЭГЕ в Мари-Турекском районах). Экогеографические единицы, согласно тем же принципам, объединили в экогеографические агрегации (ЭГА). Мы выделили одну ЭГА на территории Новоторъяльского и две ЭГА в Мари-Турекском районах (рис. 6).
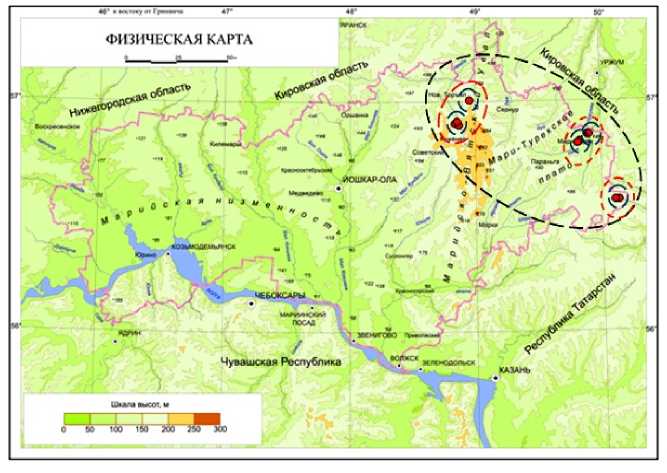
Рис. 6. Экогеографические единицы и экогеографические агрегации S. verticillata , спроецированные на карту Республики Марий Эл (Ekologo-geograficheskii atlas) (синими пунктирными линиями отмечены границы ЭГЕ, красными – границы ЭГА, черным – территория, нуждающаяся в охране)
Fig. 6. Ecogeographic units and ecogeographic aggregations of S. verticillata , projected onto the map of the Mari El Republic (Ekologo-geograficheskii atlas) (blue dotted lines mark the boundaries of the EGE, red lines mark the boundaries of the EGA, black lines mark the area in need of protection) 282
На следующем этапе работы предстоит проведение генетического анализа. Он позволит оценить корректность объединения популяций в ЭГЕ путем анализа изменчивости ДНК-маркеров внутри и между ЭГЕ (по показателям генетического сходства или расстояния между популяциями, генетическим кластерам особей или популяций, оценкам межпопуляционных генных обменов и пр.). Кроме того, анализ карты транспортных путей Республики Марий Эл позволил заметить близкое расположение ЦП S. verticillata к автодорогам, которые соединены между собой (рис. 7).
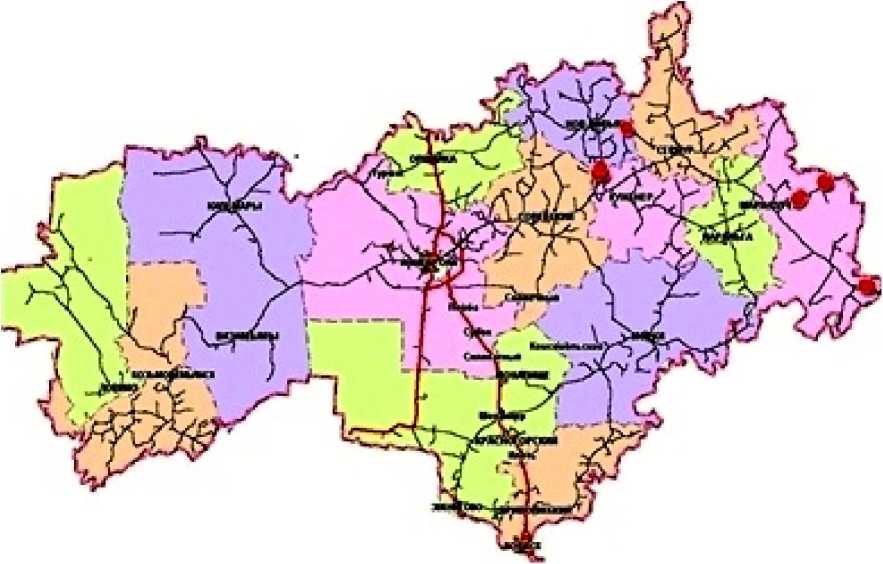
Рис. 7. Местообитания исследованных ценопопуляций S. verticillata, спроецированные на карту транспортных путей Республики Марий Эл (Ekologo-geograficheskii atlas) (исследованные местообитания отмечены красными кружками)
Fig. 7. Habitats of the studied coenopopulations of S. verticillata , projected onto the map of transport routes of the Republic of Mari El (Ekologo-geograficheskii atlas) (the studied habitats are marked with red circles)
Это может служить еще одним доказательством генетического родства особей разных ЦП S. verticillata .
На данном этапе работы генетический анализ не проведен, но собранные данные, их обработка и анализ позволяют нам выдвинуть гипотезу генетического родства изученных ЦП S. verticillata . В дальнейшем стоит задача сбора материала для генетического теста и формирования экогеографических единиц на основе данных экологии и генетики (Sinushkina, Osmanova, 2021).
З АКЛЮЧЕНИЕ
В настоящее время самым инновационным и информативным методом изучения популяционной структуры вида является экогеографический подход, сочетающий в себе как традиционные геоботанические методы исследования, так и современные технологии генетического анализа с помощью ДНК-маркеров. В данной работе экогеографический подход был применен к изучению организации популяций редкого для Республики Марий Эл растения – S. verticillata .
Данный вид не является типичным представителем флоры республики, так как приурочен к степным местообитаниям. Анализ экологической характеристики редких и исчезающих видов растений Республики Марий Эл показал, что по факторам трех климатических шкал (Tm, Om, Cr) краснокнижные виды проявляют большую стенобионтность, чем по фактору континентальности климата (Kn) и освещенности-затенения (Lc). Проанализировав S. verticillata по климатическим факторам, можно сказать, что вид довольно нетребователен к температуре, освещенности и влажности климата. Объект изучения не выходит за рамки фундаментальной экологической ниши, проявляя мезобионтность как по климатическим, так и по почвенным факторам (Itклим.= 0.55; ItLc = 0.56; Itпочв. = 0.53). Поэтому на территории региона S. verticillata не может реализовать все возможные потенции, находясь в узких диапазонах экологических параметров.
Состояние изученных 10 ЦП S. verticillata неоднозначно. ЦП 1–5 нормальные, полночленные (кроме ЦП 2). Значительная доля участия в ЦП 1–5 особей прегенеративного и генеративного периодов указывает на высокую семенную продуктивность S. verticillata и способствует поддержанию устойчивости популяции, но сильная антропогенная и зоогенная (ЦП 6–10) нагрузки могут стать губительными для численности этого вида.
Для оценки устойчивости популяции важно знать связанность соседних ЦП генными потоками. Определив генетическую гомогенность близко расположенных ЦП, можно рассчитать расстояние разноса пыльцы и диаспор. Эти сведения важны не только для изучения структуры популяций S. verticillata, но и для обнаружения путей распространения данного вида на территории Республики Марий Эл. Эта отдельная область исследования, лишь частично затрагиваемая в данной работе, но полученные результаты позволят в дальнейшем более подробно изучить появление и пути распространения S. verticillata в Средней России.
Выделенные ЭГЕ в дальнейшем можно рассматривать как потенциальные объекты для создания новых ООПТ. Эта задача наиболее актуальна, так как большинство ЦП S. verticillata в Республике Марий Эл не входят в ООПТ, исключением являются ЦП Волжского района, находящиеся в национальном парке «Марий Чодра» и ЦП Новоторъяльского района, расположенные на территории Комплексного памятника природы республиканского значения «Остепненные склоны и леса по реке Пича».
Поэтому встает проблема сохранения данного вида на территории Новоторъяльского, Моркинского и Мари-Турекского районов. Мониторинг объектов животного и растительного мира, занесенных в Красную книгу Республики Марий Эл, проведенный в 2013 г., показал, что в местообитаниях S. verticillata в Новоторъяльском и Мари-Турекском районах периодически выпасается скот, что без сомнений, ставит под угрозу способность ЦП к самоподдержанию и самосохранению. Наглядный тому пример – ЦП 6–10 S. verticillata .
Необходимо также подчеркнуть, что в изученных местообитаниях обнаружены и другие редкие виды растений, такие как: астрагал серповидный, живокость высокая, дремлик темнокрасный, лапчатка прямая, котовник венгерский, осока горная, остролодочник волосистый, фиалка горная. Поэтому выделение границ будущих природоохранных территорий позволит сохранить не только ЦП S. verticillata, но и ЦП этих редких видов.
