Организация мозговой активности при мысленном проговаривании слов у мужчин и женщин

Автор: Кривко Е.М., Бахтин О.М., Кирой В.Н., Лазуренко Д.М., Шапошников Д.Г.
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Физиология
Статья в выпуске: 2, 2022 года.
Бесплатный доступ
Цель работы - изучение гендерных различий в паттернах когерентности ЭЭГ, связанных с мысленным проговариванием слов. Материалы и методы. На 10 юношах и 10 девушках в возрасте 21±0,3 года исследовалось влияние гендерных различий на характеристики паттернов пространственной синхронизации ЭЭГ, ассоциированных с мысленным проговариванием слов, обозначающих направления в пространстве: вверх, вниз, влево, вправо, вперед, назад. Результаты. Показано, что у юношей наблюдается доминирование левого, тогда как у девушек -правого полушария, где формируются специфические паттерны когерентности ЭЭГ. Дискриминация мысленно произносимых слов по значениям когерентности ЭЭГ у юношей эффективно реализуется по паттернам когерентности, формирующимся отдельно как в левом, так и правом полушариях, тогда как у девушек - только при учете паттернов когерентности обоих полушарий. Наличие гендерных особенностей в структурно-функциональной организации мозга и характеристиках его биоэлектрической активности нельзя недооценивать, в частности при разработке ИМК на базе ЭЭГ и внутренней речи. Известно, что внутреннее (мысленное) проговаривание слов отражается как в собственно кортикальной, так и в поверхностно регистрируемой биоэлектрической активности мозга. Регистрация активности с использованием метода ЭЭГ имеет очевидные преимущества, однако до настоящего времени практически не известно, как внутренняя речь и ее характеристики проявляются с учетом гендерных различий. Выводы. Полученные результаты подтверждают наличие гендерных различий в ЭЭГ-характеристиках мозга при мысленном произнесения слов, обозначающих направления в пространстве.
Ээг, вербальная активность, когерентность, гендерные особенности, внутренняя речь
Короткий адрес: https://sciup.org/14124542
IDR: 14124542 | УДК: 612 | DOI: 10.34014/2227-1848-2022-2-81-91
Organization of brain activity in men and women during mental pronunciation of words
The purpose of the study is to examine gender differences associated with the mental pronunciation ofwords in EEG coherence patterns. Materials and Methods. The authors studied the impact of gender differences on the characteristics of EEG spatial synchronization patterns associated with the mental pronunciation of words denoting direction: up, down, left, right, forward, backward. The study enrolled 10 boys and 10 girls, aged 21±3. Results. The dominance of the left hemisphere was observed in young men, while the right hemisphere dominated in girls. Specific patterns of EEG coherence are formed in both left and right hemispheres. Discrimination of mentally pronounced words according to the EEG coherence values in young men is effectively implemented according to coherence patterns, which form separately in both left and right hemispheres, whereas in girls the coherence patterns of both hemispheres should be taken into account. Gender peculiarities in the structural and functional organization of the brain and the characteristics of its bioelectrical activity should not be underestimated, in particular, when developing a BCI based on EEG and inner speech. It is known that the internal (mental) pronunciation of words is reflected both in the cortical and superficially recorded bioelectrical activity of the brain. EEG recording of the activity has obvious advantages. However, so far little is known about manifestation of inner speech and its characteristics with respect to gender. Conclusion. The results obtained confirm gender differences in the EEG characteristics of the brain during the mental pronunciation of words denoting directions.
Текст научной статьи Организация мозговой активности при мысленном проговаривании слов у мужчин и женщин
Введение. Человек, лишенный возможности выразить себя вербально, но при этом имеющий сохранную внутреннюю речь, может оказаться в социальной изоляции. Интерфейс «мозг – компьютер» (ИМК) – это технология, способная обеспечить управление активностью мозга, произвольно генерируемой пользователем. Внутреннюю речь в парадигме ИМК можно рассматривать как естественный канал коммуникации для парализованных лиц с сохранным интеллектом и как дополнительный канал коммуникации для лиц, работающих в условиях дефицита времени и информационных перегрузок. Перспективность разработки ИМК на основе паттернов ЭЭГ-актив- ности, сопровождающих внутреннюю речь пользователя, очевидна.
Согласно теории Л.С. Выготского, внутренняя речь формируется в процессе вербального развития, в котором внешний дискурс (голосовая речь) постепенно интернализируется во внутреннюю речь [1]. В соответствии с этими представлениями внутреннюю речь можно рассматривать как мышление в словах. Показано, что внутреннее (мысленное) проговаривание слов отражается как в собственно кортикальной [2, 3], так и в поверхностно регистрируемой биоэлектрической активности мозга [4, 5]. Регистрация этой активности с использованием метода ЭЭГ имеет очевидные преимущества, однако до настоящего времени мало известно, как в ней отражаются внутренняя речь и ее характеристики [6].
Так как в настоящее время в понятие «гендер» вкладывается различное содержание, считаем необходимым уточнить, что в данной статье оно отождествляется с понятием «пол».
На наличие гендерных различий в структурно-функциональной организации мозга, в частности в функциональной специализации полушарий неокортекса, указывалось в целом ряде исследований.
Так, например, клинические наблюдения позволили прийти к выводу о том, что вербальные способности и пространственное восприятие в мозге женщин представлены более билатерально, чем в мозге мужчин [7, 8]. Однако впоследствии вывод о левополушарном доминировании в речевых процессах у мужчин был подвергнут сомнению, поскольку, как отмечалось, для соответствующего обобщения было недостаточно информации [9, 10]. Тем не менее в дальнейших исследованиях было отчетливо продемонстрировано влияние этого фактора не только на цитоархитектонику областей мозга, но и на морфофункциональную организацию и характер их взаимосвязей [11]. Результаты морфологических, электрофизиологических и биохимических исследований мозга человека также указывали на наличие гендерных различий [12]. Гендерные различия обнаруживались также в активности мозга при выполнении одной и той же задачи [13]. Так, было показано, что при реализации когнитивных задач у мужчин более выражена полушарная латерализация [14], межполушарные взаимодействия более выражены у женщин, тогда как внутриполушарные – у мужчин [15, 16].
На основании полученных результатов была сформулирована гипотеза, согласно которой особенности мозга женщин при познавательной деятельности, которые обнаруживаются при выполнении ряда речевых задач, могут быть описаны так называемой моделью двусторонней вербальной репрезентации [17]. Согласно этой модели женщины используют для реализации вербальных функций оба полушария, тогда как мужчины – преимуще- ственно левое. Результаты, полученные с использованием метода дихотического прослушивания при анализе коэффициентов латера-лизации (LQ), действительно свидетельствуют о выраженном левополушарном доминировании в процессе речи у мужчин [18]. Мужской мозг более асимметрично функционирует также при обработке пространственных ориентиров в качестве навигационных средств и при выполнении визуальных тестов [19, 20].
Гендерные различия обнаруживаются в функциональной организации мозга при реализации процессов внимания и памяти [21, 22]. Механизмы внимания при распознавании речи характеризуются сенсомоторным торможением в мозге женщин и прогностической сенсомоторной активацией в мозге мужчин [23]. Усилению функциональных связей между корковыми структурами могут способствовать, как это было показано экспериментально, женские половые гормоны эстрадиол и прогестерон [24]. Исследования c использованием метода ЭЭГ также демонстрируют гендерные различия в анализе фонетических сигналов как во время восприятия речи, так и при ее воспроизведении [13, 25].
Таким образом, имеется достаточное количество доказательств существования гендерных различий в структурно-функциональной организации мозга и его биоэлектрической активности при реализации речевых процессов. Однако до настоящего времени согласованность развития ритмической активности мозга в пределах полушарий и между ними, адекватным показателем которой является когерентность, была изучена недостаточно.
Цель исследования. Изучение гендерных различий в паттернах когерентности ЭЭГ, связанных с мысленным проговариванием слов, обозначающих направления в пространстве.
Материалы и методы. В исследованиях приняли участие 10 юношей и 10 девушек (студенты ЮФУ, средний возраст – 21±0,3 года). Все они ранее участвовали в психофизиологических экспериментах, были правшами и не имели отклонений в состоянии здоровья. Эксперименты проводились в соответствии с Хельсинкской декларацией, от всех студентов было получено письменное согласие на участие в исследовании.
Во время обследования участники располагались в свето- и звукоизолированной камере в кресле на расстоянии 1 м от экрана монитора, на котором предъявлялось стилизованное изображение циферблата часов с бегущей стрелкой. Элементарное смещение стрелки соответствовало одному делению, т.е. одной «секунде». Изображение имело размер 7 угловых градусов и предъявлялось на фоне серого экрана монитора. Положения стрелки, выделенные красным цветом, соответствовали моментам, когда обследуемый должен был произносить слово, указанное в центре циферблата. При совпадении стрелки с первой меткой слово следовало произнести вслух, с последующей – мысленно и т.д.
При совпадении стрелки с позицией, соответствующей 12 часам, в центре циферблата появлялось новое слово. Каждому обследуемому было предъявлено 6 слов, обозначающих направления в пространстве: вверх, вниз, влево, вправо, вперед, назад. Каждое слово в процессе эксперимента произносилось вслух и мысленно не менее 27 раз (9×3). Параллельно осуществлялась регистрация фонограммы через микрофон чувствительностью 2,24 мВ/Па. Для записи фонограммы использовался анализатор речи Speech Analyzer (version 3.0.1. 1996-2007 SIL International / All Rights Reserved). Как регистрация фонограммы, так и проговаривание слов вслух осуществлялись исключительно для контроля выполнения задания, анализ особенностей мужского и женского голосов не проводился.
Перед основным экспериментом проводился обучающий сеанс, в течение которого обследуемые должны были в аналогичной ситуации произносить указанные слова вслух и мысленно. Это позволяло адаптировать обследуемых к условиям эксперимента и установить уровень записи их голосов.
ЭЭГ регистрировалась непрерывно, монополярно от 14 отведений (f3, f4, f7, f8, Fp1, Fp2, c3, c4, t3, t4, t5, t6, p3, p4) по международной системе 10×20 с использованием электроэнцефалографа-анализатора «ЭНЦЕФАЛАН 131» производства «МЕДИКОМ-МТД» (г. Таганрог, Россия). Расположение электродов позволяло, согласно литературным данным [26], регистрировать активность зон Брока и Вернике, а также ряда других неокортикальных регионов, связанных с разными аспектами речевой деятельности. Референтные электроды располагались на мочках ушей. Частота дискретизации электрограмм составляла 250 Гц по каждому каналу, полоса пропускания частотных фильтров – 1–70 Гц, использовался режектор-ный фильтр 50 Гц. По отдельному каналу записывались метки, соответствующие моментам предъявления на циферблате слов, а также информация о том, как следовало произнести слово – вслух или мысленно.
Для анализа электрограмм в апостериорном режиме программно по меткам отбиралось не менее 25 безартефактных ЭЭГ-эпох, соответствующих произнесению каждого слова вслух или мысленно. Для всех отобранных фрагментов ЭЭГ и пар отведений рассчитывались коэффициенты когерентности (КоГ) на частотах гамма-1 (30–45 Гц) и гамма-2 (55– 70 Гц) ритмов. Выбор для анализа высокочастотного диапазона был обусловлен тем, что, как это было показано ранее [4, 27, 28], наиболее информативные признаки для разделения паттернов активности мозга, связанных с речью, регистрируются именно в области высоких (30–200 Гц) ЭЭГ-частот.
Массивы вычисленных значений КоГ подвергались Z-преобразованию для приближения к нормальному распределению. Сравнительный анализ массивов полученных значений, соответствующих мысленному произнесению слов, проводился с использованием многофакторного дисперсионного анализа MANOVA FACTORIAL в рамках генеральных линейных моделей с использованием программного пакета Statistica 8. Дизайн анализа включал факторы «Гендер» (уровни: юноши, девушки), «Обследуемые» (уровни: 20 чел.), «Состояние» (уровни: покой, вверх, вниз, вправо, влево, вперед, назад), «Тип произнесения» (уровни: вслух, мысленно), «Локализация» (уровни: ЛП, ПП). Зависимыми переменными являлись значения КоГ для всех пар отведений. Равенство дисперсий оценивалось с использованием Levene’s test (F=506,5±10,5 при p<(0,002±0,01)).
Результаты и обсуждение. Проведенные нами ранее исследования с использованием аналогичной методики показали, что при мыс- ленном проговаривании слов в ЭЭГ практически не регистрируются мышечные артефакты [29]. Это позволяет анализировать и трактовать наблюдаемые в ЭЭГ гендерные различия, в т.ч. в области гамма-частот, исключительно в связи с речевой деятельностью.
В состоянии покоя средние значения КоГ ЭЭГ, рассчитанные для гамма-частот и всех пар отведений, составили 0,61±0,02. Статистически значимые различия между обследуемыми отсутствовали, причем как при попарном сравнении, так и при сравнении гендерных групп. Различия отсутствовали и между показателями когерентности гамма-1- и гам-ма-2-частот, причем практически по всем парам отведений (рис. 1) как у девушек, так и у юношей.
С учетом отсутствия различий в показателях КоГ гамма-1- и гамма-2-частот дальнейший анализ проводился совместно для обоих частотных диапазонов.
Вербальная деятельность сопровождалась достоверным повышением уровня когерент- ности (df=82, t=3,7, p<0,001), причем в ЭЭГ юношей более выраженно, чем в ЭЭГ девушек: 0,77±0,02 и 0,67±0,01 соответственно (F(21, 1372)=79,7, p<0,00).
Анализ внутриполушарной когерентности показал наличие различий как по фактору «Гендер» (F(21, 980)=80,1, p<0,00), так и по фактору «Латеральность» (F(21, 980)=50,3, p<0,00), а также на уровне их взаимодействия (F(21, 980)=42,2, p<0,00). Указанные различия были связаны с тем, что при мысленном произнесении слов (по сравнению с состоянием покоя) уровень когерентности ЭЭГ у юношей был достоверно выше в парах отведений левого полушария (F(21, 1372)=30,5, p<0,00), тогда как у девушек – в парах отведений правого полушария (F(21, 1372)=26,7, p<0,00) (рис. 1). Гендерные различия наблюдались и во внут-риполушарной когерентности, причем в левом полушарии они были несколько более выражены (левое полушарие – F(21, 1372)=60,03, p<0,00, правое полушарие – F(21, 1372)=36,3, p<0,00).
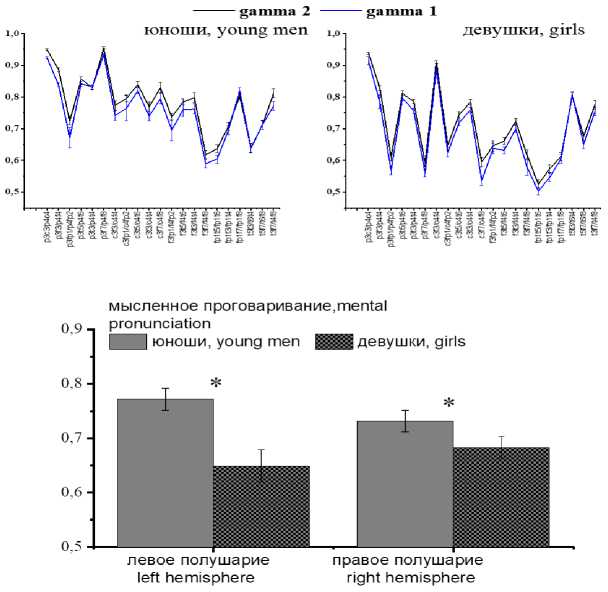
Рис. 1. Средние значения КоГ ЭЭГ юношей и девушек в парах отведений левого и правого полушарий при мысленном произнесении слов (по оси Y – значения КоГ, вертикальные линии – ошибки средних, * – достоверные различия (p<0,05))
Fig. 1. Average CoG EEG values in boys and girls in pairs of leads of the left and right hemispheres during mental pronunciation of words (Y axis – CoG values; vertical lines – mean errors, * – the differences are significant (p<0.05))
Анализ значений КоГ ЭЭГ, зарегистрированной при мысленном произнесении разных слов, проводился с использованием процедур попарного (Planed Comps) и множественного (Tukey’s test) сравнения. Различия в паттернах когерентности ЭЭГ, ассоциированных с разными словами, у юношей имели место для пар отведений как левого, так и правого полушарий, тогда как у девушек – только правого. При этом как у девушек, так и у юношей достоверно различались одни и те же пары слов
(рис. 2): вверх – назад (девушки – F(21, 940)=22,6, p<0,00; юноши – F(21, 940)=2,2, p<0,002), вниз – назад (девушки – F(21, 940)=8,5, p<0,003; юноши – F(21, 940)=1,7, p<0,02), влево – назад (девушки – F(21, 940)=16,2, p<0,00006; юноши – F(21, 940)=3,2, p<0,00003), вверх – вправо (девушки – F(21, 940)=5,8, p<0,01; юноши – F(21, 940)=1,8, p<0,001). Кроме того, у девушек различия обнаруживались в паре слов «вправо – назад» (F(21, 940)=5,4, p<0,02).
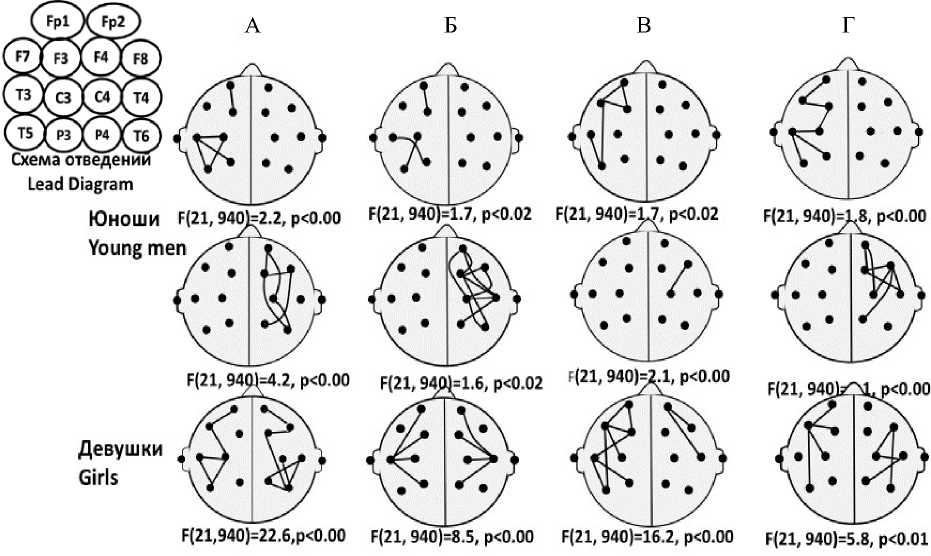
Рис. 2. Топография пар отведений ЭЭГ, позволяющих дискриминировать слова, обозначающие направления в пространстве, при их мысленном произнесении юношами и девушками: А) вверх – назад; Б) вниз – назад; В) влево – назад; Г) вверх – вправо
Fig. 2. Topography of EEG derivation pairs, which makes it possible to discriminate words denoting direction.
The words are mentally pronounced by boys and girls:
А – “Up-Back”, B – “Down-Back”, C – “Left-Back”, D – “Up-Right”
Анализ пространственной структуры пар отведений, позволяющих дискриминировать мысленно произносимые слова, показал, что в левом полушарии она включает ряд речевых зон, а именно: IFG (inferior frontal gyrus), STG (superior temporal gyrus) и MTG (middle temporal gyrus), а в правом – как гомологичные области, так и ряд других [30]. При этом если у юношей паттерн отведений, дискрими- нирующий слова, включал лишь 15 % пар отведений одного из полушарий, то у девушек – 27 % пар отведений двух полушарий.
Раннее нами [4] была продемонстрирована возможность идентификации паттернов ЭЭГ, специфичных для речевой деятельности, в частности для внутренней речи. На такую возможность указывают и другие авторы [31]. Однако нами была исследована лишь группа юношей, а вопрос о гендерных различиях оставался открытым. Анализ литературы указывает на наличие гендерных различий как в реализации вербальных функций структурами мозга, так и в возникающей при этом его биоэлектрической активности.
Приведенные выше результаты подтверждают наличие гендерных различий при мысленном произнесении слов, обозначающих направления в пространстве. Во-первых, повышение когерентности в этих условиях (по сравнению с состоянием покоя) было более выражено на гамма-частотах в ЭЭГ юношей. Во-вторых, мысленное произнесение слов девушками требовало синхронизации активности связанных с речевыми процессами областей не только левого полушария (что характерно для юношей), но и правого. Наконец, различение мысленно произносимых слов у юношей имело место при анализе когерентности ЭЭГ пар отведений как левого, так и правого полушарий, тогда как у девушек – только на основе одновременного учета пар отведений из разных полушарий.
Изучение взаимосвязи топологии и взаимодействия сетей мозга с различными аспектами вербальной деятельности в последнее десятилетие активно ведется с использованием метода фМРТ и технологий нейровизуализации [32, 33], которые, однако, достаточно сложно использовать в системах ИМК, ориентированных на широкий круг конечных пользователей. Результаты этих исследований показали, что области неокортекса, связанные с воспроизведением и пониманием реальной речи, локализованы как в левом, так и правом полушариях мозга [33]. При этом произнесение слов требует активации не только моторной коры, но также областей, связанных со слуховым восприятием, семантической обработкой и др., вследствие чего складывающаяся интеграция включает целый ряд неокорти-кальных регионов, в частности верхнюю и среднюю височные извилины, височный полюс, угловую извилину, височно-теменное соединение, нижнюю лобную извилину, премоторную и медиальную префронтальную кору, островок, предклинье, таламус и хвостатое ядро [33]. Контроль воспроизведения речи реализуется сетями, ответственными за соб- ственно произнесение и понимание речи, – бороздой Хешля, теменной и дополнительной моторной корой, за собственно воспроизведение – сетями левой задней и вентральной передней теменной доли [32], а также специфической левосторонней лобно-височно-теменной сетью, которая активируется во время повествовательной речи [34]. В целом есть основания полагать, что функциональные связи между областями неокортекса, которые находят отражение в показателях когерентности, обеспечивают формирование не только состояния бодрствования (в т.ч. покоя, resting-state network, RSN), но и речевых процессов (speech production network, SPN), в частности внутренней речи. Показано, что первые из них преимущественно связаны с зонами правого полушария, тогда как вторые – с зонами левого [35], что, по-видимому, и определяет сдвиг связности процессов, регистрируемых в ЭЭГ, от правого полушария в покое к левому в условиях речевой деятельности [34], а также сложные взаимодействия глобальных мозговых сетей, контролирующих воспроизведение речи [35].
В рамках указанных представлений полученные нами результаты могут быть интерпретированы следующим образом. В левом полушарии юношей-правшей при мысленном проговаривании слов активируется нейронная сеть, контролирующая воспроизведение речи (SPN). При этом в правом полушарии сохраняются активными сети, контролирующие уровень бодрствования (RSN). Структура обеих сетей позволяет различать ряд мысленно произносимых юношами слов, обозначающих направления в пространстве. У девушек, для которых, как известно, в значительной степени характерна образная стратегия мышления [12], реализация которой требует привлечения ресурсов обоих полушарий, такое различение возможно только на основе анализа как лево-, так и правополушарной когерентности гамма-частот. При решении практических задач, связанных с формированием в технологии ИМК команд управления на основе паттернов когерентности ЭЭГ, ассоциированных с внутренней речью, паттерн отведений, таким образом, должен учитывать гендерную принадлежность пользователя.
Выводы. Экспериментально показано наличие гендерных различий в показателях когерентности ЭЭГ, регистрируемой при мысленном проговаривании юношами и девушками слов, обозначающих направления в пространстве.
-
1. Уровень когерентности гамма-частот при мысленном произнесении слов, обозначающих направления в пространстве, более существенно повышается в ЭЭГ юношей.
-
2. Специфические паттерны когерентности ЭЭГ, позволяющие осуществлять дискриминацию мысленно произносимых слов, у
- юношей формируются как в левом, так и в правом полушарии, тогда как у девушек такая дискриминация возможна только на основе когерентности гамма-частот, регистрируемых от отведений как левого, так и правого полушария.
-
3. Обнаружение паттернов когерентности ЭЭГ, позволяющих с высокой достоверностью различать мысленно произносимые слова, указывает на перспективность использования ассоциированных с внутренней речью паттернов когерентности в системах ИМК.
Список литературы Организация мозговой активности при мысленном проговаривании слов у мужчин и женщин
- Выготский Л.С. Мышление и речь: сборник. Москва; 2020. 250.
- Martin S., Millân José delR., KnightR., Brian T., Pasley N. The use of intracranial recordings to decode human language: Challenges and opportunities. Brain & Language. 2019; 193: 73-83.
- Cooney C., Folli R., Coyle D. Neurolinguistics research advancing development of a direct-speech brain-computer interface. IScience. 2018; 8: 103-125.
- Кирой В.Н., Бахтин О.М., Миняева Н.Р., Лазуренко Д.М., Асланян Е.В., Кирой Р.И. Электрографические корреляты внутренней речи. Журн. высш. нерв. деят. 2015; 65 (5): 616-625.
- Kiroy V.N., Bakhtin O.M., Krivko E.M., Lazurenko D.M., Aslanyan E.V., Shaposhnikov D.G., Shcher-ban I. V. Spoken and Inner Speech-related EEG Connectivity in Different Spatial Direction. Biomedical Signal Processing and Control. 2022; 71: 103-224. DOI: https://doi.org/10.1016/j.bspc.2021.103224.
- De Lima Xavier L., Hanekamp S., Simonyan K. Sexual dimorphism within brain regions controlling speech production. Frontiers in neuroscience. 2019; 13: 795.
- McGlone J. Sex differences in functional brain asymmetry. Cortex. 1978; 14 (1): 122-128.
- McGlone J. Sex differences in human brain asymmetry: A critical survey. Behavioral and brain sciences. 1980; 3 (2): 215-227.
- Obleser J. Gender differences in functional hemispheric asymmetry during processing of vowels as reflected by the human brain magnetic response. Neuroscience letters. 2001; 314 (3): 131-134.
- Frost J.A. Language processing is strongly left lateralized in both sexes: Evidence from functional MRI. Brain. 1999; 122 (2): 199-208.
- Gong G., He Y., Evans A.C. Brain connectivity: gender makes a difference. The Neuroscientist. 2011; 17 (5): 575-591.
- Боголепова И.Н., Агапов П.А., Малофеева И.Г., Пилецкая И.А. Межполушарная асимметрия передней лимбической области коры мозга человека. Международный журнал прикладных и фундаментальных исследований. 2021; 5: 7-11.
- Moctezuma L.A., MolinasM. Sex differences observed in a study of EEG of linguistic activity and resting-state: Exploring optimal EEG channel configurations. 7th International Winter Conference on Brain-Computer Interface (BCI). 2019; 6: 1-6.
- Hirnstein M., Hugdahl K., Hausmann M. Cognitive sex differences and hemispheric asymmetry: A critical review of 40 years of research. Laterality: Asymmetries of Body, Brain and Cognition. 2019; 24 (2): 204-252.
- IngalhalikarM. Reply to Joel and Tarrasch: On misreading and shooting the messenger. Proceedings of the National Academy of Sciences. 2014; 111 (6): E638.
- Scheuringer A., Harris T.A., Pletzer B. Recruiting the right hemisphere: Sex differences in inter-hemispheric communication during semantic verbal fluency. Brain and Language. 2020; 207.
- Gur R.C., Alsop D., Glahn D., Petty R., Swanson C.L., Maldjian J.A. An fMRI study of sex differences in regional activation to a verbal and a spatial task. Brain Lang. 2000; 74: 157-170.
- Packheiser J. A large-scale estimate on the relationship between language and motor lateralization. Scientific reports. 2020; 10 (1): 1-10.
- Lenroot R.K., Giedd J.N. Sex differences in the adolescent brain. Brain and cognition. 2010; 72 (1): 46-55.
- Hiscock M. Is there a sex difference in human laterality? II. An exhaustive survey of visual laterality studies from six neuropsychology journals. Journal of Clinical and Experimental Neuropsychology. 1995; 17 (4): 590-610.
- Вольф Н.В. Половые различия функциональной организации процессов полушарной обработки речевой информации. Ростов-на-Дону: ЦВВР; 2000. 240.
- Langrova J. Gender impact on electrophysiological activity of the brain. Physiological research. 2012; 61.
- Thornton D. Sex differences in early sensorimotor processing for speech discrimination. Scientific reports. 2019; 9 (1): 1-13.
- Peper J.S. Sex steroids and connectivity in the human brain: a review of neuroimaging studies. Psycho-neuroendocrinology. 2011; 36 (8): 1101-1113.
- Sato M. The neurobiology of sex differences during language processing in healthy adults: A systematic review and a meta-analysis. Neuropsychologia. 2020; 140.
- Qureshi M.N.I. Multiclass classification of word imagination speech with hybrid connectivity features. Transactions on Biomedical Engineering. 2017; 65 (10): 2168-2177.
- Martin S. Decoding spectrotemporal features of overt and covert speech from the human cortex. Frontiers in neuroengineering. 2014; 7: 14.
- Muller L. Spatial resolution dependence on spectral frequency in human speech cortex electrocorticogra-phyю. Journal of neural engineering. 2016; 13 (5): 056013.
- Бахтин О.М., Кривко Е.М., Кирой В.Н. Электромиографические компоненты, ассоциированные с внутренней речью. Журнал медико-биологических исследований. 2020; 8 (2): 111-120.
- FriedericiA.D. The brain basis of language processing: from structure to function. Physiological reviews. 2011; 91 (4): 1357-1392.
- Vorontsova D. Silent EEG-Speech Recognition Using Convolutional and Recurrent Neural Network with 85 % Accuracy of 9 Words Classification. Sensors. 2021; 21 (20): 6744.
- SimmondsA.J. Parallel systems in the control of speech. Human brain mapping. 2014; 35 (5): 1930-1943.
- Silbert L.J. Coupled neural systems underlie the production and comprehension of naturalistic narrative speech. Proceedings of the National Academy of Sciences. 2014; 111 (43): E4687-E4696.
- Geranmayeh F. Overlapping networks engaged during spoken language production and its cognitive control. Journal of Neuroscience. 2014; 34 (26): 8728-8740.
- Simony an K., Fuertinger S. Speech networks at rest and in action: interactions between functional brain networks controlling speech production. Journal of neurophysiology. 2015; 113 (7): 2967-2978.
