Осмотический потенциал как фактор адаптации зостеры морской (Zostera marina L.) к условиям местообитанияя
 к условиям местообитанияя")
Автор: Корзунина Анастасия андреевнА., Марковская Евгения федоровнА.
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 3 (94), 2008 года.
Бесплатный доступ
В статье рассматривается влияние условий произрастания на величину осмотического потенциала клеток листа зостеры морской (Zostera marina L.), произрастающей в Кандалакшском заливе Белого моря. Соленость воды в местах исследования варьируется от 12 до 22 ‰. Диапазон значений осмотического потенциала в этих условиях составил 14-34 кПа. Показано увеличение осмотического потенциала с увеличением солености воды. В статье также обсуждается роль осмотического потенциала в адаптации зостеры к условиям произрастания на литорали.
Осмотический потенциал, соленость
Короткий адрес: https://sciup.org/14749435
IDR: 14749435 | УДК: 581.522.4
Текст научной статьи Осмотический потенциал как фактор адаптации зостеры морской (Zostera marina L.) к условиям местообитанияя
В настоящее время все больший интерес исследователей вызывает жизнедеятельно сть организмов, обитающих на стыке морской и континентальной сред. На побережьях морей формируется совершенно особая категория местообитаний, характерной чертой которых является нестабильная соленость. Это маршевые луга, эстуарии рек и ручьев, супралито-ральные ванны и ванны верхнего и среднего горизонтов литорали. Нестабильность солености, а также некоторых других гидрохимических показателей в таких водоемах обусловлена главным образом приливно-отливными и штормовыми движениями морской воды, с одной стороны, и влиянием атмосферных процессов – с другой [1]. В результате комбинации этих факторов возникают уникальные, крайне изменчивые во времени условия для развития различных организмов. В отличие от животных организмов, растения не могут изолироваться от этих перемен или поменять свое пространственное положение, а поэтому вы
нуждены функционировать в переменно ритмичных условиях среды, которые имеют место на литорали. Одним из таких объектов является вторичноводное высшее сосудистое растение – зостера. Широкий спектр ее местообитаний: от литорали (на обсушке береговой линии) до сублиторали на глубине 2–4 м [2] свидетельствует о большом адаптационном потенциале. Однако пути адаптации этого вида к различному уровню солености в Кандалакшской губе Белого моря и переменной суточной солености на литорали во время отлива до настоящего времени не изучены.
Целью работы было оценить роль осмотического потенциала в адаптации зостеры ( Zostera marina L.) к условиям засоления и градиенту солености в условиях литорали.
МАТЕРИАЛ И МЕТОДИКА
Объектом исследования является зостера (Zostera marina L.), взморник морской – погру- женное многолетнее растение. На Белом море зостера произрастает на литорали, выходит также и на сублитораль. В зависимости от рельефа может занимать пояс до 30 м [2].
РАЙОН ИССЛЕДОВАНИЯ
Работа выполнена в точках, расположенных в Кандалакшском заливе Белого моря между Кандалакшским и Карельским берегами. Длина залива – 185 км, ширина у входа – 67 км. В Кандалакшский залив впадают реки Нива, Ковда, а также множество ручьев, которые способствуют опреснению водоема. Температура воды на поверхности летом в среднем до 18,5 ºС, в мелких, защищенных от ветров, хорошо прогреваемых губах и бухтах вода прогревается сильнее – до 25 ºС. Зимой – от –1,0 до 1,5 ºС. С первой декады октября по май большая часть Кандалакшского залива покрыта льдом: от вершины почти до Турьего мыса и до Кемь-луд, в мягкие зимы только до Порьей губы.
Соленость воды зимой – 27–29 ‰, летом – от 0 до 20–25 ‰. Это объясняется тем, что в Белое море с речным стоком поступает большое количество пресной воды, которая снижает концентрацию солей в верхних водных слоях. У поверхности воды, особенно вблизи устьев рек, наблюдаются резкие сезонные колебания солености в связи с изменением объема речных стоков. Наибольшее опреснение происходит в разгар половодья [7].
Исследования проводились в 3 точках Кандалакшского залива: остров Горелый, губа Долгая, остров Двойной, на растениях зостеры, произрастающих в различных условиях: на литорали, сублиторали и на опресняемых участках.
ОПИСАНИЯ ИССЛЕДОВАННЫХ МЕСТ ПРОИЗРАСТАНИЯ ЗОСТЕРЫ
На острове Двойном сообщество зостеры небольшое: примерно 2×1,5 м в литоральной ванне. Грунт песчано-илистый. При отливе не обсыхает, уровень воды во время отлива примерно 15–20 см, вода на отливе сильно нагревается. Находится с защищенной стороны острова.
Губа Долгая – это небольшой залив, глубоко вдающийся в материк. Сильные ветры и волно-бой, идущие с моря, наблюдаются редко или совсем отсутствуют. В губу впадает множество небольших пресных ручейков. Грунт илистый или песчаный. Сообщества зостеры здесь сильно разрежены, располагаются на песчаной литорали, хорошо защищены от действия волн.
На острове Горелом зостера образует сплошной «ковер» на литорали и сублиторали. Грунт песчано-илистый. Характерно наличие многочисленных «ключей» пресной воды. Зостера на этом острове исследовалась в различных условиях произрастания: на литорали, сублиторали и возле ручья, то есть в условиях опреснения.

Рис. 1. Побережье о. Горелый
Литоральные растения зостеры во время отлива сверху покрыты значительным слоем нитчатых водорослей, что в некоторой степени защищает зостеру от высыхания. На участке у ручья побеги располагаются несколько реже, на отливе практически не обсыхают, так как постоянно омываются водой из ручья. Температура воды в ручье ниже, чем на литорали во время прилива. В солнечные дни в июне на литорали вода может прогреваться до 20 °С и выше, а температура в ручье значительно ниже – около 10 °С, поэтому на отливе растения сильно охлаждаются (рис. 1).
МЕТОДИКА СБОРА
И ОБРАБОТКИ МАТЕРИАЛА
Материал был собран в течение двух летних периодов: в 2005 и 2006 годах в июне – июле. Сбор проб проводился в 3 различных точках Кандалакшского залива: о. Горелый, губа Долгая, о. Двойной. На о. Горелый пробы были взяты в различных горизонтах побережья (литораль и сублитораль), а также в опресняемых участках. Также были собраны пробы зостеры, произрастающей на разной глубине. Для этого на о. Горелый была сделана трансекта, начинающаяся с верхней границы произрастания зостеры и на 12 метров уходящая в море. Пробы были взяты с каждого метра трансекты. Для определения осмотического потенциала клеток листа собирали растения зостеры в местах с различными условиями произрастания: на литорали, на сублиторали и на опресняемых участках. При этом также брались пробы воды, плотно сть воды измеряли с помощью ареометра и затем по таблицам переводили плотность в соленость.
Осмотический потенциал клеток листа зос-теры определяли плазмолитическим методом (рис. 2) [4].

Рис. 2. Плазмолиз в клетках Zostera marina :
А – нормальные клетки, Б – плазмолизированные клетки, 1 – хлоропласты, 2 – клеточная оболочка
РЕЗУЛЬТАТЫ
В таблице приведены данные по осмотическому потенциалу клеток листа зостеры, произрастающей в акватории Кандалакшской губы: на разной глубине и при различной солености воды.
Максимальные значения (32.1–34.7 кПа) осмотического потенциала в обоих годах исследования были отмечены у растений зостеры, произрастающих на литорали, а минимальные (14.8–23 кПа) – на опресняемых участках острова Горелый. Диапазон варьирования солености воды (12–22 ‰) уже, чем диапазон варьирования осмотического потенциала (14–34 кПа), что может свидетельствовать о большом адаптивном потенциале этого вида. Исследование показало, что с увеличением солености возрастает осмотический потенциал клеток листа (рис. 3).
Однако на графике выделяются (они отмечены квадратами) точки, в которых при одинаковом значении солености величины осмотического потенциала различаются: меньшие значения соответствуют значениям осмотического потенциала в клетках листьев растений сублиторали, большие – клеткам листьев растений на литорали. Соленость воды во время прилива на литорали и сублиторали одинаковая, но во время отлива условия резко изменяются, и это различие может составлять 10 ‰ [1]. Также изменяются и другие факторы – температура, влажность, освещенность. Если на сублиторали они варьируют незначительно, то на литорали, в пределах приливно-отливной зоны, изменения этих факторов даже в течение суток очень существенны.
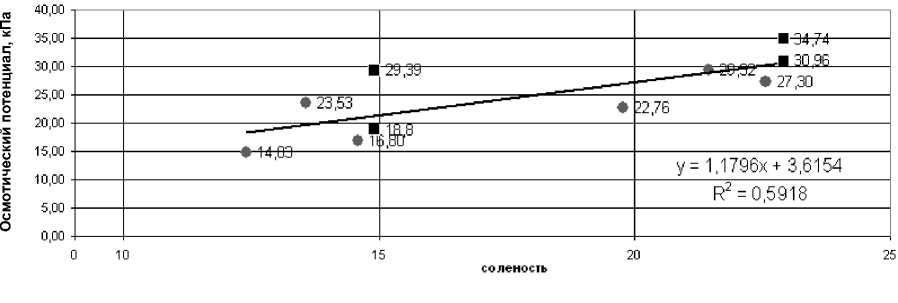
Рис. 3. Зависимость осмотического потенциала листа Zostera marina от солености воды
Осмотический потенциал листьев зостеры, произрастающей в различных условиях засоления
|
Места взятия проб |
Год |
№ пробной площадки |
Экологические условия, соленость, ‰ |
Величина осмотического потенциала вегетативного листа, кПа |
|
О. Горелый литораль |
2005 |
- |
22,90 |
34,7±2,1 |
|
О. Горелый сублитораль |
- |
22,90 |
30,9±1,2 |
|
|
О. Горелый опреснение |
- |
13,57 |
23,5±1,3 |
|
|
О. Двойной |
- |
22,56 |
27,3±2,3 |
|
|
Губа Долгая литораль |
- |
21,46 |
29,3±1,3 |
|
|
О. Горелый опреснение |
2006 |
- |
12,39 |
14,8±1,4 |
|
О. Горелый литораль |
1 |
14,91 |
29,4±2,5 |
|
|
3 |
32,1±1,3 |
|||
|
5 |
23,7±2,7 |
|||
|
О. Горелый сублитораль |
7 |
14,91 |
22,7±4,1 |
|
|
10 |
18,8±1,4 |
|||
|
Губа Долгая литораль |
- |
14,59 |
16,8±1,4 |
Исследования динамики изменения осмотического потенциала в листьях зостеры во время действия приливно-отливной волны (рис. 4) показали, что на отливе уже через 15 минут отмечается увеличение осмотического потенциала (с 15,9 до 25,7 кПа), которое сохраняется в листьях зос-теры и после прилива. Снижение потенциала происходит медленнее, чем его повышение.
Изучение осмотического потенциала растений в зависимости от глубины произрастания показало, что эта связь описывается линейным уравнением с достаточно высоким уровнем значимости
(рис. 5). Более высокий осмотический потенциал во время отлива у литоральных растений позволяет им лучше приспособиться к условиям среды.
Растения на литорали во время отлива обсыхают, в их тканях может изменяться температура, а повышенное осмотическое давление клеток позволяет им дольше удерживать воду и благодаря этому оставаться жизнеспособными. Таким образом, диапазон осмотического потенциала у зостеры можно рассматривать как интегральный показатель степени акклимированности вида к условиям произрастания.
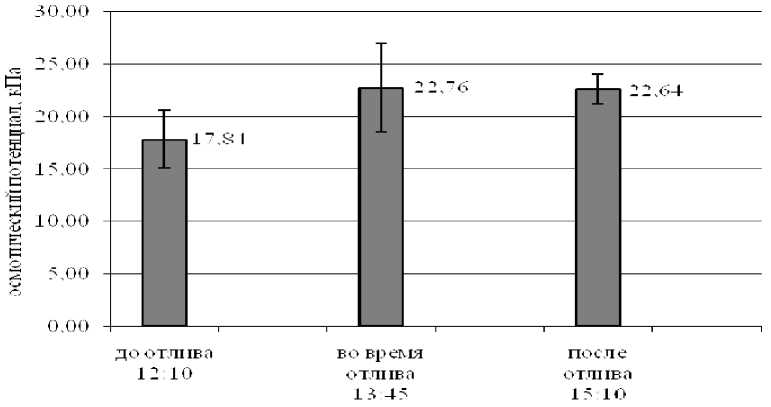
Рис. 4. Динамика осмотического потенциала клеток листьев зостеры (о. Горелый, соленость воды – 19,77 ‰, 2006 г.)
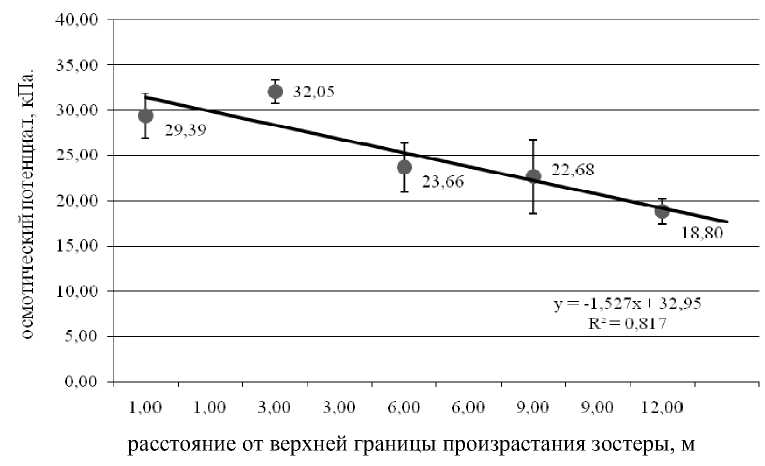
Рис. 5. График зависимости осмотического потенциала от глубины произрастания
ОБСУЖДЕНИЕ
Зостера обитает в широком диапазоне солености воды. По данным Т. Мейера и С. Неринга, в Балтийском море зостера может произрастать при солености от 3 до 30 ‰ [14], по данным Амельзадеха с соавторами, в Японском море – от 5 до 33 ‰ [11]. Диапазон солености воды в местах произрастания зостеры в Кандалакшском заливе Белого моря более узкий и варьируется от 14 до 23 ‰, при этом диапазон осмотического потенциала клеток листа зостеры включает значения от 14,8 до 32,1 кПа.
В условиях повышенной солености почвенного слоя и воды одним из путей наиболее широко распространенной адаптации является изменение осмотического потенциала клетки, который может повышаться как за счет накопления в клетках повышенного количества осмотически активных гидрофильных ионов солей, так и низкомолекулярных органических соединений, в том числе углеводов [10]. Осмотический потенциал клеток листа зостеры на отливе увеличивается с 15,9 до 25,7 кПа. Быстрое изменение осмотического потенциала у зостеры во время отлива и суточная цикличность этих изменений дает основание предположить, что в этой реакции должны быть задействованы вещества основного обмена. Такой механизм был обнаружен у другого литорального растения – Ruppia maritime L. [15]. При экспериментальном погружении растений этого вида в раствор, соленость которого в 2 раза превышает соленость, в которой были выращены растения, осмотический потенциал клеток за одну минуту увеличивался почти на 50 %. Это происходило за счет растворимых углеводов (их концентрация увеличилась на 34 %) и растворимого пролина (его концентрация увеличилась на 18 %). Однако известно, что синтез и распад пролина в листьях зостеры происходит медленно, при изменении солености воды [12]. Как и руппия, зостера относится к группе галофитов, и можно предположить, что для поддержания осмотического потенциала этого вида также могут использоваться осмотически активные сахара. Увеличение содержания низкомолекулярных углеводов на отливе может происходить за счет распада крахмала [16], сахарозы [17], синтеза глюкозы de novo [16]. В работе У. Карстена на антарктических зеленых водорослях литорали было показано, что синтез глюкозы наблюдается при солености воды в 68 ‰, а содержание крахмала в это время остается постоянным, то есть это увеличение может быть связано с синтезом de novo. Важно отметить, что у зостеры были получены высокие значения фотосинтеза при произрастании в условиях высокой солености (до 33 ‰) [13]. Это означает, что в этих условиях синтезируются углеводные продукты, которые могут выполнять осморегуляторную функцию. Однако эти предположения нужно проверить экспериментально.
В заключение мы выражаем глубокую благодарность руководству Кандалакшского заповедника за возможность экспериментальной работы на его территории и Г. А. Шкляревич за помощь в организации экспедиций.
Список литературы Осмотический потенциал как фактор адаптации зостеры морской (Zostera marina L.) к условиям местообитанияя
- Бурковский И. В. Структурно-функциональная организация и устойчивость морских донных сообществ. М.: Изд-во МГУ, 1992. 208 с..
- Вехов В. Н. Зостера морская (ZOSTERA MARINA L.) Белого моря. М.: Изд-во МГУ, 1992. 144 с.
- Вехов В. Н. Некоторые вопросы экологии зостеры на Белом море//Биология Белого моря. Т. 4. М.: Изд-во МГУ, 1974. С. 65-79..
- Викторов Д. П. Малый практикум по физиологии растений. М.: Высшая школа, 1969. 135 с.
- Жизнь растений/Под ред. А. Л. Тахтаджяна. Т. 6. М.: Просвещение, 1982. С. 39-41..
- Зубов Н. Н. Океанологические таблицы. Л.: Гидрометеорологическое изд-во, 1957. 406 с..
- Карпович В. Н. «Кандалакшский заповедник», Заповедники европейской части РСФСР. I. М.: Мысль, 1988. С. 20-60..
- Малый практикум по физиологии растений/Под ред. Т. В. Гусева М.: Изд-во МГУ, 1982. 192 с..
- Тахтаджян А. Л. Система и филогения цветковых растений. М.; Л.: Наука, 1966. С. 468.
- Физиология растений: Учебник для студентов ВУЗов/Под ред. И. П. Ермакова. М.: Академия, 2005. С. 514-552..
- Alemzadeh A. et al. ZMVHA-B1, the gene for subunit B of vacuolar H-ATPase from the Zostera marina L. is able to replace vma2 in yeast null mutant//Journal of bioscience and bioengineering. 2006. 102(5). P. 390-395..
- Diggelen J., Rozema J., Broekman R. Mineral composition of and proline accumulation by zostera marina L. in response to environmental salinity//Aquatic Botany. 1987. 27 (2). P. 169-176..
- Hellblom F., Björk M. Photosynthetic responses in Zostera marina to decreasing salinity, inorganic carbon content and osmolality//Aquatic Botany. 1999. 65 (1-4). P. 97-104..
- Meyer T., Nehring S. Anphlanzung von Seegraswiesen (Zostera marina L.) als interne Massnahme zur restaurierung der Ostsee//Rostok Meersbiologie beitrag. Heft 15. 2006. S. 105-119..
- Murphyb L. R, Kinseya S. T. Duroko M. J. Physiological effects of short-term salinity changes on Ruppia maritime//Aquatic Botany. 2004. 76 (4). Р. 329-337..
- Karsten U. Okologische Untersuchungen zur Salinitats -und Temperaturtolerz antarktischer Grunalgen unter besonderer Beruksichtigung des b-Dimethylsulfoniunpropionat (DMPS) -Stofwechsels//ePIC (electronic Publication Information Center) of Alfred Wegener Institute for Polar and Marine Research (AWI) [Electronic resource]. 1991. 120 p. Mode access:http://epic.awi.de/Publications/BerPolarforsch199179.pdf>..
- Тouchettea B. W., Burkholderb Jo A. M. Carbon and nitrogen metabolism in the seagrass, Zostera marina L.: Environmental control of enzymes involved in carbon allocation and nitrogen assimilation//Journal of Experimental Marine Biology and Ecology. 2007. 350 (1-2). Р. 216-233.
