Основные закономерности распределения хирономид в донных сообществах центрального района Воткинского водохранилища в многолетнем аспекте

Автор: Алексевнина Маргарита Степановна, Преснова Елена Владимировна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 2, 2019 года.
Бесплатный доступ
Обобщены данные мониторинговых наблюдений за изменениями структуры хирономидных сообществ центрального района Воткинского водохранилища. Стационарные исследования проводились на базе Камской биологической станции с 1975 по 2014 гг. Зообентос собирали в течение вегетационного сезона на правобережном и левобережном мелководье, а также в глубоководной зоне водоема. Установлен таксономический состав, выявлены доминантные комплексы, рассмотрены количественные показатели развития личинок хирономид на разных биотопах водохранилища. Высокие качественные и количественные показатели развития хирономидоценозов отмечены на залитой пойме левобережного мелководья, здесь складываются благоприятные условия для развития личинок хирономид. Изучение сезонной динамики численности и биомассы личинок хирономид показало, что большинство популяций, обитающих на правобережном мелководье, являются моноцикличными, на левобережье распространены как моноцикличные, так и дицикличные виды. Наблюдения позволили установить многолетние периоды, в течение которых изменялась продуктивность и видовой состав хирономидоценозов, что связано с поступлением в водохранилище органических загрязнений.
Хирономиды, сообщество, структура, мониторинг, воткинское водохранилище, биотоп
Короткий адрес: https://sciup.org/147227082
IDR: 147227082 | УДК: 574.587 | DOI: 10.17072/1994-9952-2019-2-197-205
The main regularities of distribution chironomids in benthic communities the central area of the Votkinsk reservoir in long-term perspective
The data of monitoring observations of changes in the structure of chironomid communities of the central region of the Votkinsk reservoir are summarized. Stationary studies were conducted on the basis of the Kama Biological Station from 1975 to 2014. Zoobenthos was harvested during the growing season in the right-bank and left-bank shallow waters, as well as in the deep-water zone of the water. The taxonomic composition has been established, dominant complexes have been identified, quantitative indicators of the development of chironomid larvae on different biotopes of the reservoir have been considered. High qualitative and quantitative indicators of the development of chironomidocenoses are noted on the flooded floodplain of the left-bank shallow water, favorable conditions for the development of chironomid larvae develop here. The study of the seasonal dynamics of the abundance and biomass of chironomid larvae showed that the majority of the chironomid populations living in the right-bank shoal are monocyclic, and both monocyclic and dicyclic species are common on the left bank. Monitoring observations made it possible to establish long-term periods during which the productivity and species composition of chironomidocenoses changed, which is connected with the entry of organic pollutants into the reservoir.
Текст научной статьи Основные закономерности распределения хирономид в донных сообществах центрального района Воткинского водохранилища в многолетнем аспекте
В пресноводных экосистемах наиболее разнообразной в видовом отношении группой являются личинки комаров звонцов (Diptera,Chironomidae).
Составляя 30–40% видового состава донных животных, они обеспечивают устойчивость и стабильность бентоценозов. Высокая скорость продуцирования органического вещества личинками хи-рономид определяет большую долю их в общей
Таблица 1
Видовой состав и частота встречаемости* личинок хирономид на разных биотопах центрального района Воткинского водохранилища в период 1975–2014 гг.
|
Таксон |
Правобережное мелководье |
Русло |
Левобережное мелководье |
|
подсемейство Tanypodinae |
4 |
4 |
4 |
|
Ablabesmyia (Ablabesmyia) monilis (Linnaeus, 1758) |
– |
– |
+ |
|
Anatopynia plumipes (Fries, 1823) |
– |
+ |
– |
|
Procladius (Holotanypus) choreus (Meigen, 1804) |
+ |
+ |
+ |
|
Procladius (Holotanypus) ferrugineus (Kieffer, 1918) |
+ |
+++ |
++ |
|
Psectrotanypus varius (Fabricius, 1787) |
– |
+ |
+ |
|
Tanypus (Tanypus) punctipennis Meigen, 1818 |
+ |
– |
– |
|
Thienemannimyia geijskesi (Goetghebuer, 1934) |
+ |
– |
– |
Продолжение табл. 1
|
Таксон |
Правобережное мелководье |
Русло |
Левобережное мелководье |
|
подсемейство Diamesinae |
0 |
1 |
2 |
|
Potthastia gaedii (Meigen, 1838) |
– |
+ |
+ |
|
Potthastia longimanus Kieffer, 1922 |
– |
– |
+ |
|
подсемейство Prodiamesinae |
1 |
1 |
1 |
|
Monodiamesa bathyphila (Kieffer, 1918) |
+ |
++ |
++ |
|
подсемейство Orhtocladiinae |
6 |
7 |
9 |
|
Cricotopus (Cricotopus) bicinctus (Meigen, 1818) |
– |
– |
+ |
|
Cricotopus (Isocladius) sylvestris (Fabricius, 1794) |
+ |
+ |
+ |
|
Cricotopus (Paratrichocladius) sp . |
++ |
+ |
++ |
|
Nanocladius (Nanocladius) dichromus (Kieffer, 1906) |
+ |
+ |
– |
|
Orthocladius (Pogonocladius) consobrinus (Holmgren, 1869) |
– |
– |
+ |
|
Parakiefferiella bathophila (Kieffer, 1912) |
– |
– |
+ |
|
Parakiefferiella triquetra (Pankratova, 1970) |
+ |
+ |
+ |
|
Psectrocladius (Psectrocladius) psilopterus (Kieffer, 1906) |
++ |
+ |
++ |
|
Psectrocladius (Psectrocladius) simulans (Johannsen, 1937) |
+ |
+ |
+ |
|
Psectrocladius sp . |
– |
+ |
+ |
|
подсемейство Chironominae |
25 |
22 |
28 |
|
Триба Chironomini |
20 |
18 |
21 |
|
Chernovskiia orbicus (Townes, 1945) |
– |
+ |
– |
|
Chironomus (Chironomus) cingulatus Meigen, 1830 |
+ |
++ |
++ |
|
Chironomus (Chironomus) muratensis Meigen, 1830 |
+ |
++ |
+ |
|
Chironomus (Chironomus) plumosus (Linnaeus, 1758) |
+ |
++ |
+ |
|
Cladopelma viridulum (Linnaeus, 1767) |
+ |
– |
– |
|
Cryptochironomus (Cryptochironomus) defectus (Kieffer, 1913) |
++ |
+ |
++ |
|
Cryptochironomus (Cryptochironomus) ussouriensis (Goetghebuer, 1933) |
– |
+ |
– |
|
Demicryptochironomus (Demicryptochironomus) vulneratus (Zetterstedt, 1838) |
+ |
+ |
+ |
|
Dicrotendipes nervosus (Staeger, 1839) |
++++ |
+ |
++ |
|
Dicrotendipes tritomus (Kieffer, 1916) |
– |
+ |
+ |
|
Endochironomus albipennis (Meigen, 1830) |
– |
+ |
+ |
|
Endochironomus tendens (Fabricius, 1775) |
+ |
– |
– |
|
Fleuria lacustris Kieffer, 1924 |
+ |
+ |
++++ |
|
Glyptotendipes (Glyptotendipes) barbipes (Staeger, 1839) |
+ |
– |
+ |
|
Glyptotendipes (Glyptotendipes) glaucus (Meigen, 1818) |
+ |
– |
– |
|
Harnischia curtilamellata (Malloch, 1915) |
+++ |
++ |
++ |
|
Harnischia fuscimanus Kieffer, 1921 |
+ |
– |
+ |
|
Lauterborniella agrayloides (Kieffer, 1911) |
– |
– |
+ |
|
Microchironomus tener (Kieffer, 1918) |
– |
– |
+ |
|
Parachironomus gracilior (Kieffer, 1918) |
– |
– |
+ |
|
Parachironomus vitiosus (Goetghebuer, 1921) |
– |
+ |
+ |
|
Paralauterborniella nigrohalteralis (Malloch, 1915) |
++ |
+ |
+ |
|
Paratendipes albimanus (Meigen, 1818) |
– |
– |
+ |
|
Polypedilum (Pentapedilum) exsectum (Kieffer, 1916) |
+ |
– |
– |
|
Polypedilum (Pentapedilum) sordens (van der Wulp, 1875) |
– |
+ |
– |
|
Polypedilum (Polypedilum) nubeculosum (Meigen, 1804) |
+ |
+ |
+ |
|
Polypedilum (Tripodura) bicrenatum Kieffer, 1921 |
++ |
+++ |
++ |
|
Polypedilum (Tripodura) scalaenum (Schrank, 1803) |
++ |
++ |
+ |
|
Polypedilum (Uresipedilum) convictum (Walker, 1856) |
+ |
+ |
+ |
|
Stictochironomus sticticus (Fabricius, 1781) |
+ |
– |
– |
|
Synendotendipes impar (Walker, 1856) |
+ |
– |
– |
|
триба Tanytarsini |
5 |
4 |
7 |
|
Cladotanytarsus (Cladotanytarsus) mancus (Walker, 1856) |
++ |
++ |
++ |
Окончание табл. 1
|
Таксон |
Правобережное мелководье |
Русло |
Левобережное мелководье |
|
Cladotanytarsus (Cladotanytarsus) pallidus Kieffer, 1922 |
– |
– |
+ |
|
Micropsectra junci (Meigen, 1818) |
– |
– |
+ |
|
Stempellina bausei (Kieffer, 1911) |
+ |
– |
+ |
|
Stempellinella edwardsi Spies & Saether, 2004 |
+ |
– |
– |
|
Tanytarsus gregarius Kieffer, 1909 |
+ |
+ |
+ |
|
Tanytarsus pallidicornis (Walker, 1856) |
+ |
+ |
+ |
|
Zavrelia pentatoma Kieffer & Bause, 1913 |
– |
+ |
+ |
|
Всего |
37 |
36 |
45 |
Примечание. *«–»– вид отсутствует, «+» – частота встречаемости менее 20%, «++» – 20–50%, «+++» – 50– 80%,«++++» –более 80%.
продукции зообентоса. Общеизвестно большое практическое значение личинок, как основного корма для многих бентосоядных рыб и водных беспозвоночных. Велика роль хирономид в процессах самоочищения водоемов, так как в сильно загрязненных и эвтрофных водоемах развиваются эврибионтные виды, которые в процессе питания используют загрязняющие вещества, прежде всего органические. Поэтому исследованиям этой группы донных животных посвящены многие монографии российских специалистов [Боруцкий, 1963; Яблонская, 1968; Соколова, 1973; Шилова, 1976; Тодераш, 1984; Балушкина, 1987; Поздеев, 2006; Зинченко, 2011 и др.].
Целью нашего сообщения является выявление закономерностей пространственного и временного распределения отдельных видов хирономид и хи-рономидных сообществ в мелководной и глубоководной зонах центрального района Воткинского водохранилища. Исследования проведены на базе Камской биостанции (г. Оханск) в 1975–2014 гг. и являются итогом мониторинговых наблюдений за зообентическими сообществами водохранилища.
Материал и методы исследования
С 1975 по 2014 гг. в отдельные годы с мая по ноябрь через каждые 10–15 дней на базе Камской биостанции проводились стационарные сборы зообентоса дночерпателем Петерсена (250 см²) в право- и левобережной литорали и глубоководной зоне водохранилища по стандартной сетке станций. За каждый вегетационный сезон отбирали около 60 бентосных проб. Методы сбора и анализа материала достаточно унифицированы [Методика …, 1975]. Для определения личинок хирономид использовали определители А.А. Черновского [1949], В.Я. Панкратовой [1970, 1977, 1983], а в последние годы – Е.А. Макарченко [1999].
Для каждого вида хирономид на разных биотопах в течение вегетационного периода подсчитаны средние значения численности ( N ) и биомассы ( B ), определена частота встречаемости ( P ), выражен-
ные в процентах. Используя эти показатели, вычисляли величину доминантного индекса [Щербина, 2001]:
Id = 3 N • B • P , что позволяет оценить их роль в бентоценозах.
В сборе и обработке материала активное участие принимали студенты-гидробиологи университета, выполняя дипломные работы и магистерские диссертации. Всем им выражаем большую благодарность.
Результаты исследований
За время наших исследований в центральном районе Воткинского водохранилища (1975– 2014 гг.) отмечено 59 видов личинок хирономид, которые представлены подсемействами Tanypodi-nae, Diamesinae, Prodiamesinae, Orthocladiinae и Chironominae . Наибольшее число видов (39) относится к подсемейству Chironominae , что составляет 66% общего числа зарегистрированных видов, или 61–66% видового богатства на разных биотопах. Роль остальных подсемейств хирономид в составе бентофауны отдельных биотопов водохранилища весьма ограничена: среди Orthocladiinae отмечено 10 видов, что обеспечивает 17–21% разнообразия хирономид на разных биотопах, из Tanypodinae зарегистрировано 7 видов (9–11%). Подсемейства Diamesinae и Prodiamesinae насчитывали в совокупности 3 вида (не более 4% видового богатства хирономид). По своим экологическим характеристикам – это эврибионтный комплекс видов, адаптированных к илистым грунтам и изменениям содержания кислорода в воде и, прежде всего, в грунтах. Особенно показателен отбор эврибионт-ных видов среди представителей Orthocladiinae как более оксифильной группы животных. Их фауна на исследованном участке представлена исключительно фитофильными и фитопелофильными видами, среди которых наиболее распространены хирономиды родов Psectrocladius , Cricotopus и Paratrichocladius (табл. 1).
Как видно из табл. 1, наибольшее видовое богатство хирономидофауны отмечено в левобережной литорали (45 видов), отличающейся пологим рельефом дна, малыми глубинами, развитием высшей водной растительности и относительно быстрым прогревом воды в течение вегетационного сезона. В русловой зоне и правобережной литорали видовое богатство хирономид не так велико (36–37 видов). Коэффициент видового сходства Чекановского-Серенсена свидетельствует о достаточно высокой степени различий между хироно-мидофаунами отдельных биотопов (33–38%). При этом б о льшие различия видового состава зарегистрированы между левобережной и правобережной литоралями водохранилища.
Структура сформировавшихся в водохранилище хирономидоценозов обусловлена не только их видовым составом, но и количественной представленностью в них отдельных видов, адаптированных к специфическим условиям биотопов. На их развитие в течение вегетационного периода большое влияние оказывают факторы среды, значительно различающиеся на разных биотопах, главные из которых, динамика вод, температурный режим, характер грунта и др. Поэтому зональное распределение личинок хирономид имеет следующие тенденции.
В 2014 г. (это последний год систематических стационарных наблюдений за бентофауной водохранилища) в правобережье водохранилища, где основной грунт – галька с песком, глиной и илом, складывается бедный в количественном отношении хирономидоценоз. Это открытое прибрежье подвержено волнобою, поэтому динамика грунтов достаточно интенсивна. На данном биотопе в разные годы отмечено 15–28 видов. Основу хироно-мидоценоза в прежние годы определяли Procladius ferrugineus и Dicrotendipes nervosus , в настоящее время, и в частности в 2014 г., доминирует Poly-pedilum nubeculosum , а в целом доминантный комплекс обеспечивает 57% численности и 75% биомассы всех личинок хирономид (рис. 1).
В составе бентоценоза русловой части водохранилища, где преобладает заиленный песок, зарегистрировано 16–26 видов личинок хирономид. В 1970–1990 гг. ярким доминантом в сообществе был Procladius ferrugineus , в последние годы лидируют виды рода Polypedilum (в основном Poly-pedilum bicrenatum и Polypedilum scalaenum ). В 2014 г. они обеспечивали более 61% биомассы хи-рономид (рис. 1).
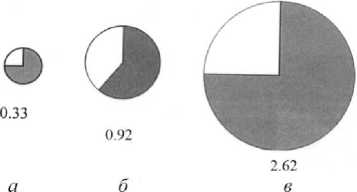
Рис. 1 . Распределение средней биомассы хиро-номид (г/м²) и доля в ней видов доминантного комплекса (заштриховано) на разных биотопах центрального района Воткинского водохранилища в 2014 г.:
а – правобережное мелководье, б – русло, в – левобережное мелководье
Хирономидное сообщество, сформированное на левобережном мелководье водохранилища, большую часть которого занимает залитая пойма, количественно и качественно более богато, здесь в разные годы отмечено от 18 до 36 видов. Ярким доминантом в последние годы является Fleuria lacustris , в доминирующий комплекс в разные периоды исследования входили виды Polypedilum nubeculosum , Chironomus muratensis , C ladotanytar-sus mancus и Procladius ferrugineus . Виды-доминанты в 2014 г. составляли 75% биомассы и 60% численности хирономидных сообществ биотопа. Обилие растительности и детрита обеспечивают благоприятные пищевые условия для личинок хирономид и прежде всего фито- и пелофиль-ных (рис. 1).
За время стационарных наблюдений была прослежена сезонная динамика численности и биомассы личинок хирономид в литоральной и глубо- ководной зонах центрального района водохранилища. Характер сезонной динамики хирономид определяют, во-первых, их жизненные циклы, во-вторых – факторы среды [Шилова, 1976; Поздеев, Островская, 2006]. Ранее было показано, что в условиях Воткинского водохранилища большинство видов хирономид моно- и дицикличны [Алексев-нина, Гореликова,1988; Алексевнина, 1992, 2001].
Сезонная динамика развития личинок хироно-мид в правобережном мелководье имеет следующие особенности (рис. 2). В мае – начале июня численность и биомасса личинок хирономид низкие, в конце июня – середине июля происходит увеличение численности при незначительном повышении биомассы, что связано с появлением младшевозрастных личинок в популяциях хироно-мид. В августе численность и биомасса продолжают расти, а в сентябре–октябре при увеличении биомассы численность снижается, что свидетельствует о присутствии в популяциях старшевозраст-ных особей и частичном вылете имаго. Общий характер динамики развития личинок хирономид в правобережье определяет доминантный вид Dicro-tendipes nervosus, который в условиях Воткинского водохранилища является моноцикличным.
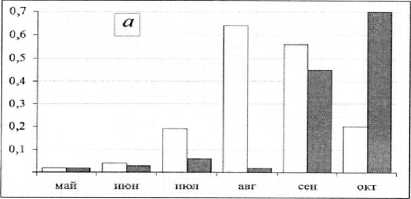
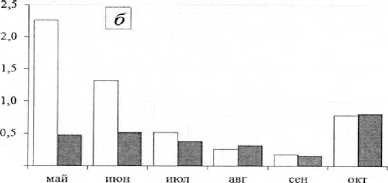
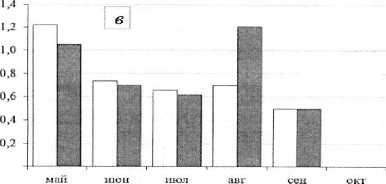
Рис. 2. Сезонная динамика численности (тыс. экз./м², столбцы без штриховки) и биомассы (г/м², столбцы со штриховкой) хирономид центральной части Воткинского водохранилища с мая по октябрь 2014 г.: а – правобережное мелководье; б – русло; в– левобережное мелководье сентябрь происходит снижение численности и биомассы личинок хирономид (рис. 2), что, как правило, связано с вылетом имаго весенне-летней генерации и выеданием их рыбой. Значительное увеличение численности и биомассы в октябре происходит за счет роста личинок и миграции личинок хирономид из прибрежных в глубоководную зону с более постоянными условиями жизни. Характер динамики численности и биомассы личинок в прежние годы определялся развитием Procladius ferrugineus [Колина, Преснова, 1980], а в настоящее время – Polypedilum bicrenatum, который в этих условиях является моноцикличным.
Сезонная динамика численности и биомассы личинок хирономид в левобережной литорали имеет 2 подъема биомассы (май и август) и высокую численность в конце весны и начале лета. Это свидетельствует о том, что наряду с моноциклич-ными видами большое значение имеют частично или полностью дицикличные. В 2014 г. характер динамики развития хирономид определяет Fleuria lacustris (рис. 2).
Таким образом, в условиях правобережья (осушаемое мелководье) большинство популяций хи-рономид моноцикличны, на бывшем русле Камы и залитой пойме наряду с моноцикличными видами распространены дицикличные, среди которых некоторые виды адаптированы к частичному вылету имаго осенью, а другая часть – весной.
Анализ многолетней динамики структуры хи-рономидоценозов центрального района Воткинского водохранилища показал, что с 1976 по 2014 гг. можно выделить периоды, значительно различающиеся по уровню продуктивности и соотношению видов в сообществах [Алексевнина, Преснова, 2007, 2017]. Так, с 1976 по 1992 гг. биомасса личинок хирономид в центральном районе Воткинского водохранилища была достаточно стабильной и составляла 0.60–0.90 г/м², в среднем – 0.73 г/м². В дальнейшем произошло снижение биомассы личинок хирономид и самые низкопродуктивные хи-рономидоценозы (0.21–0.52 г/м²) отмечены в 1993–2004 гг., в среднем – 0.4 г/м². Начиная с 2006 г., биомасса личинок комаров-звонцов увеличивается, и в 2014 г. составила 1.30 г/м², в среднем за этот период –1.1 г/м² (рис. 3).
Периодические многолетние изменения биомассы личинок хирономид, по нашему мнению, связаны с характером поступления в водохранилище органических загрязнений. В 70–80-е гг. прошлого столетия сообщества хирономид формировались в условиях постоянного поступления в водоем сбросов Краснокамского ЦБК, богатых отходами переработки целлюлозы, что привело к значительному обеднению бентоценозов, особенно в русловой зоне водохранилища. Хирономидоце-
В глубоководной зоне водохранилища с мая по нозы прибрежных мелководий были более разнообразными и продуктивными (табл. 2).
В 90-е гг. прошлого столетия поступление органических загрязнений значительно снизилось, что привело к увеличению в донных биоценозах численности двустворчатых моллюсков, которые в процессе фильтрации использовали в пищу органические взвеси. Это послужило причиной уменьшения количества животных детритофагов-собирателей и глотателей, прежде всего личинок хирономид. Так, в 1999 г. была зарегистрирована самая низкая биомасса личинок хирономид за все время существования Воткинского водохранилища
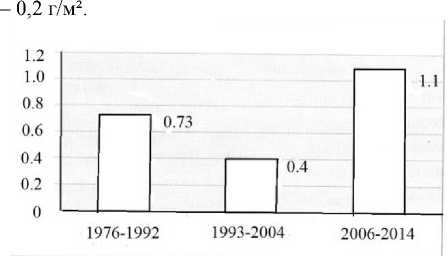
Рис. 3 . Изменение средней биомассы ( В , г/м²) хирономидных сообществ центрального района Воткинского водохранилища в период 1976–2014 гг.
С начала XXI в. продуктивность хирономидо-ценозов в водохранилище постепенно увеличивалась, особенно заметно в русловой зоне и левобережье (табл. 2).
За время исследований в структуре хирономи-доценозов в разные периоды изменились не только их весовые соотношения, но и представленность видов. При достаточном постоянстве видового разнообразия хирономидных сообществ, приуроченных к ранее описанным биотопам, произошла смена доминантных комплексов, показанная в табл. 2. В первые годы существования Воткинского водохранилища в центральном районе на всех биотопах ярким доминантом был типичный пело-фил и облигатный хищник-полисапроб Procladius ferrugineus . Видами-субдоминантами являлись Chironomus рlumosus , Dicrotendipes nervosus , Cladotanytarsus mancus , Polypedilum bicrenatum. Во второй период среди личинок хирономид доминировали виды рода Polypedilum , главным образом, Polypedilum bicrenatum .
В последние годы наблюдается некоторая специализация хирономидных сообществ по биотопам. На «залитой пойме» левобережного мелководья ярким доминантом стал Fleuria lacustris , субдоминантами можно считать Polypedilum nubecu-losum , Cladotanytarsus mancus , Endochironomus al-bipennis . На правобережном мелководье (бывшей рипали Камы) сформировалось сообщество, в котором доминируют Polypedilum nubeculosum и Di-crotendipes nervosus , а в русловой глубоводной зоне среди личинок хирономид «лидирует» Polypedi-lum bicrenatum, и только здесь сохраняет свой доминантный статус Procladius ferrugineus .
Таблица 2
Изменение структуры хирономидных сообществ ( D –доминантные виды, Sd – субдоминантные виды, B –биомасса, г/м²) на разных биотопах центрального района Воткинского водохранилища в период 1975–2014 гг.
|
Период |
Параметр |
Правобережное мелководье |
Русло |
Левобережное мелководье |
|
1975–1992 |
D |
Procladius ferrugineus |
Procladius ferrugineus |
Procladius ferrugineus |
|
Sd |
Polypedilum bicrenatum , Dicrotendipes nervosus |
Dicrotendipes nervosus |
Chironomus plumosus , Cladotanytarsus mancus |
|
|
B |
0.67 |
0.25 |
1.35 |
|
|
1993–2004 |
D |
Dicrotendipes nervosus |
Polypedilum nubeculosum |
Polypedilum bicrenatum |
|
Sd |
Polypedilum bicrenatum , Cryptochironomus defectus |
Polypedilum bicrenatum , Procladius ferrugineus |
Polypedilum nubeculosum , Chironomus muratensis , Fleuria lacustris , Cladotanytarsus mancus |
|
|
B |
0.16 |
0.40 |
0.75 |
|
|
2005–2014 |
D |
Polypedilum nubeculosum |
Polypedilum bicrenatum |
Fleuria lacustris |
|
Sd |
Dicrotendipes nervosus |
Polypedilum scalaenum, Procladius ferrugineus |
Polypedilum nubeculosum , Cladotanytarsus mancus , Endochironomus albipennis |
|
|
B |
0.37 |
0.75 |
2.20 |
По нашему мнению, в последние годы структу- ра хирономидоценозов достаточно стабилизирова- лась. В видовом составе преобладают пело- и фи-топелофильные виды, главным образом детрито-фаги-собиратели, присутствие которых позволяет считать центральный район водохранилища как β-мезосапробный.
Заключение
Анализ многолетних данных позволил установить видовой состав личинок хирономид центрального района Воткинского водохранилища (59 видов) и выявить основные тенденции их распределения по биотопам в течение периода исследования. Русловую зону, правобережное и левобережное мелководья занимают хирономидоценозы, разные по количественным показателям развития и представленности отдельных видов, прежде всего составляющих доминантный комплекс. Наиболее благоприятные условия за весь период исследования для развития комаров-звонцов складывались на залитой пойме левобережья, где сформировались разнообразные и более продуктивные сообщества. В последние годы значительно улучшились условия существования хирономидоценозов глубоководной зоны водохранилища, что привело к увеличению их видового богатства и количественных показателей развития. На характер многолетней динамики численности и биомассы хиро-номид в Воткинском водохранилище в период исследований большое влияние оказывали поступающие в водоем органические загрязнения.
Список литературы Основные закономерности распределения хирономид в донных сообществах центрального района Воткинского водохранилища в многолетнем аспекте
- Алексевнина М.С. Структура и распределение хирономидоценозов Воткинского водохранилища в районе Оханска // Биологические ресурсы камских водохранилищ и их использование: межвуз. сб. науч. тр. Пермь, 1992. C. 163-171.
- Алексевнина М.С. Состав и распределение донных сообществ центральной части Воткинского водохранилища в 1997-1998 г.г. // Рыбные ресурсы Камско-Уральского региона и их рациональное использование: материалы науч.-практ. конф. Пермь, 2001. С. 8-10.
- Алексевнина М.С., Гореликова Н.М. Зообентос // Биология Воткинского водохранилища. Иркутск: Иркут. ун-т, 1988. С. 65-97.
- Алексевнина М.С., Преснова Е.В. Бентофауна как элемент мониторинга экосистемы Воткинского водохранилища // Современные проблемы водохранилищ и их водосборов: тр. междунар. науч.-практ. конф. Пермь, 2007. Т. 2. С. 188-192.
- Алексевнина М.С., Преснова Е.В. Изменение структуры бентоценозов Воткинского водохранилища за время его существования (1964-2014 гг.) // Вестник Пермского университета. Сер. Биология. 2017. Вып. 3. С. 328-332.
- Балушкина Е.В. Функциональное значение личинок хирономид в континентальных водоемах. Л.: Наука, 1987. 179 с.
- Боруцкий Е.В. Вылеты Chironomidae (Diptera) континентальных водоемов разных климатических поясов как фактор обеспеченности рыб пищей // Зоологический журнал. 1963.Т. 42, вып. 2. С. 233-247.
- Зинченко Т.Д. Эколого-фаунистическая характеристика хирономид (Diptera, Chironomidae) малых рек бассейна Средней и Нижней Волги (Атлас). Тольятти: Кассандра, 2011. 258 с.
- Колина Т.Н., Преснова Е.В. Динамика популяций Procladius ferrugineus K. (Diptera, Chironomidae) в Воткинском водохранилище // Биологические ресурсы водоемов Западного Урала. Пермь, 1980. С. 46-52.
- Макарченко Е.А. Комары-звонцы (Chironomidae) // Определитель пресноводных беспозвоночных России и сопредельных территорий / под ред. С.Я. Цалолихина. СПб., 1999. Т. 4. Высшие насекомые, двукрылые. С. 210-216.
- Методика изучения биогеоценозов внутренних водоемов // под ред. Ф.Д. Мордухай-Болтовского. М.: Наука, 1975. 240 с.
- Панкратова В.Я. Личинки и куколки комаров подсемейства Orthocladiinae фауны СССР (Diptera, Chironomidae = Tendipedidae). Л.: Наука, 1970. 344 с.
- Панкратова В.Я. Личинки и куколки комаров подсемейства Podonominae и Tanypodinae фауны СССР (Diptera, Chironomidae = Tendipedidae). Л.: Наука, 1977. 154 с.
- Панкратова В.Я. Личинки и куколки комаров подсемейства Chironominae фауны СССР (Diptera, Chironomidae = Tendipedidae). Л.: Наука, 1983. 296 с.
- Поздеев И.В. Роль личинок хирономид в структуре донных сообщнеств рек бассейна Верхней и Средней Камы: дис. … канд. биол. наук. СПб., 2006. 198 с.
- Поздеев И.В., Островская Ю.В. Сезонная динамика и продукция массовых видов хирономид предгорной реки Среднего Урала (р. Чусовая) // Вестник Башкирского ун-та. 2006. № 4. С. 55-56.
- Соколова Н.Ю. Особенности биологии массовых видов хирономид Учинского водохранилища // Комплексные исследования водохранилищ. М.: Изд-во МГУ. 1973. Вып. 2. С. 101-122.
- Тодераш И.К. Функциональное значение хирономид в экосистемах водоемов Молдавии. Кишинев: Штиинца, 1984. 172 с.
- Черновский А.А. Определитель личинок комаров семейства Tendipedidae. М.; Л.: Изд-во АН СССР, 1949. 187 с.
- Шилова А.И. Хирономиды Рыбинского водохранилища. Л.: Наука, 1976. 251 с.
- Щербина Г.Х. Сравнительный анализ структуры макрозообентоса на участках верхнего и нижнего бьефов Рыбинского гидроузла // Биология внутренних вод, 2002, № 3. С. 44-45.
- Яблонская Е.А. Опыт применения метода Е.В. Боруцкого для определения продукции хирономид // Методы определения продукции водных животных. Минск: Вышэйш. школа, 1968. С. 204-218.
