Особенности адаптации вторичной ксилемы кустарников и кустарничков к условиям Альпийского пояса (Северный Кавказ)
")
Автор: Чавчавадзе Евгения Савельевна, Умаров Мухади Умарович, Сизоненко Ольга Юрьевна, Волкова Светлана Борисовна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 3, 2017 года.
Бесплатный доступ
Изучено семь видов аэроксильных и простратных кустарников и кустарничков из альпийского пояса (Северный Кавказ), определен комплекс ксилотомических признаков, способствующих адаптации этих растений к суровым условиям высокогорий.
Вторичная ксилема, древесина, водопроводящая ткань, ксилотомические признаки, альпийский пояс, адаптация, северный кавказ
Короткий адрес: https://sciup.org/146279444
IDR: 146279444 | УДК: 581.522.5:
Adaptation of the secondary xylem of shrubs and semifrutexs of the Alpine zones (North Caucasus)
Seven species of aeroxylic and prostrate shrubs and semifrutexs of the Alpine zones (North Caucasus) were studied. The xylotomycal feature complex facilitating the adaptation of wood plants to the severe conditions of the high mountains was defined.
Текст научной статьи Особенности адаптации вторичной ксилемы кустарников и кустарничков к условиям Альпийского пояса (Северный Кавказ)
Введение. Альпийским называют высокогорный пояс, расположенный между субальпийским и нивальным поясами в горах умеренных и субтропических широт. В районах наших исследований он лежит выше 2300-2500 м. Климат высокогорий достаточно суровый с частыми снегопадами и сильными ветрами, а также интенсивной солнечной радиацией. Растительный покров развивается в условиях обильного, долго не сходящего снега, отсутствия многолетнего промерзания почвы (несмотря на низкие температуры), а также достаточного увлажнения при хорошем дренаже (Энциклопедия географических терминов, 1968). Он представлен низкотравными лугами и зарослями, как правило, стелющихся кустарников и кустарничков, распространенных небольшими пятнами на скелетных горно-луговых почвах, осыпях и моренах. Видовой состав древесных растений очень скуден (Галушко, 1975). Насколько известно, при хорошо исследованной систематике, ксилотомические особенности альпийских растений этого района Кавказа в комплексе не рассматривались, хотя отдельные таксоны из других горных мест обитаний были описаны (Яценко-Хмелевский, 1946, 19546; Умаров, 1992; Чавчавадзе, Сизоненко, 2002; Чавчавадзе и др., 2004; Бенькова, Швейнгрубер, 2004; Carlquist 1988).
Представленная статья посвящена выявлению комплекса признаков, специфичных для структуры вторичной ксилемы видов, произрастающих в суровых условиях альпийского пояса восточных территорий Северного Кавказа.
Материал и методика. Изучена структура водопроводящей ткани (древесины) 7 видов, относящихся к 7 родам 5 семейств двудомных покрытосеменных растений, собранных в природе на склонах Андийского хребта (2300-2500 м н.у.м.) и перевала Харами (2420 м н.у.м.), а также отобранных в Гербарии БИН РАН: Dry as caucasica Juz. - Дриада кавказская (сем. Rosaceae), Pentaphylloides fruticosa (L.) O. Schwarz. - Курильский чай кустарниковый (сем. Rosaceae), Salix kazbekensis A. Skvorts. - Ива казбекская (сем. Salicaceae), Empetrum caucasicum (V.Vassil.) Juz. - Водяника кавказская (сем. Empetraceae), Rhododendron caucasicum L. - Рододендрон кавказский (сем. Ericaceae), Vaccinium myrtillus L. - Черника обыкновенная (сем. Ericaceae), Ribes orientate Desf. - Смородина восточная (сем. Grossulariaceae), В основном это небольшие, как правило, простратные кустарнички (представители p.p. Dryas, Pentaphylloides, Empetrum, Vaccinium) и кустарники (p.p. Rhododendron, Ribes и Salix) (Галушко, 1978; Умаров и др., 2008).
Образцы каждого вида взяты в 3-х-кратной повторности. Срезы для микропрепаратов приготовлены на замораживающем микротоме фирмы Reichert (Австрия) в трех плоскостях - поперечной, тангентальной, радиальной. Ксилотомические описания и микрометрия выполнены по традиционным методикам (Яценко-Хмелевский, 1954а; Metcalfe., Chalk, 1983), с использованием терминологии, предложенной Международной Ассоциацией анатомов древесины - 1AWA (Weeler, Baas, 1998) и словаря терминов, приведенного коллективом авторов в Атласе древесины и волокон для бумаги (Атлас древесины и волокон для бумаги, 1992). Структурный анализ вторичной ксилемы взятых видов проведен с применением фотонного микроскопа Axio Scope Al Zeiss и методики кодирования признаков, включающей 20 групп признаков и более 160 их вариаций (Умаров и др., 2007).
Результаты и обсуждение. Состав древесины рассмотренных видов довольно однообразен. Помимо постоянных элементов -члеников сосудов, клеток радиальных лучей и аксиальной паренхимы, она может включать только волокнистые трахеиды (p.p. Empetrum, Rhododendron, Vaccinium) или только волокна либриформа (р. Salix), виды родов Dryas, Pentaphylloides, отличаются более богатым составом, включающим сосудистые трахеиды, a Ribes orientate - волокна либриформа, часто септированные. В древесине некоторых исследованных видов спорадически наблюдаются трахеидоподобные сосуды с одной перфорацией на боковой стенке.

Рис. 1 . Salix kazbekensis - рассеяннососудистый тип древесины (поперечный срез)

Рис. 2 . Rhododendron caucasicum - древесина полукольцесосудистого типа (поперечный срез)
Вторичная ксилема рассмотренных видов преимущественно рассеяннососудистая или только рассеяннососудистая (р. Salix), часто в сочетании с полукольцесосудистой (p.p. Dryas, Empetrum, Rhododendron, Ribes, Pentaphylloides, Vaccinium, Ribes) (рис. 1, 2), в очень узких слоях прироста - кольцесосудистая, с одним рядом более крупных просветов вдоль границы в ранней части годичного кольца.
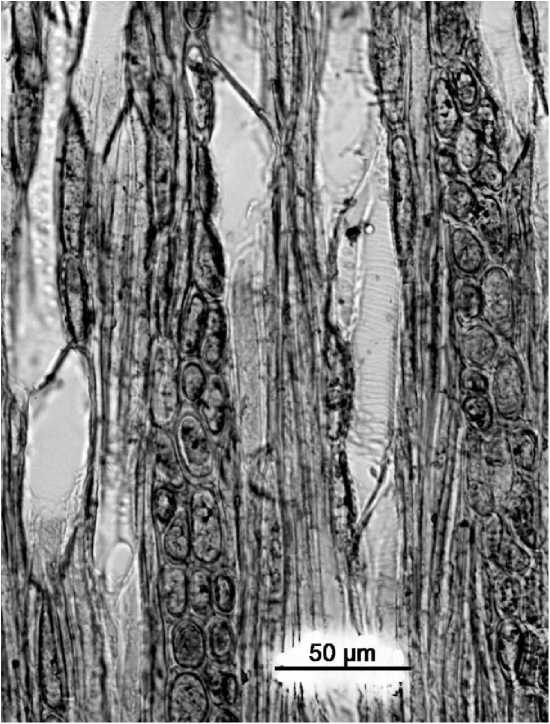
Рис. 3 . Vaccinium myrtillus — спиральные утолщения на стенках члеников сосудов (тангентальный срез)
Членики сосудов чрезвычайно малого или малого диаметра - от 20 до 60 мкм, исключение - сосуды у представителей рода лапчатка с просветами до 80-100 мкм. Форма члеников сосудов волокновидная и цилиндрическая (длина превышает диаметр в 8-11 и в 2-7 раз соответственно), или, в основном, цилиндрическая (рр. Dryas, Ribes, Salix); членики сосудов с короткими и длинными клювиками, иногда лишены их. Стенки сосудов со спиральными утолщениями или без них (рис. 3).
Перфорационные пластинки лестничные (p.p. Empetrum, Rhododendron), лестничные, лестнично-сетчатые (р. Ribes), лестничные в сочетании с простыми (p.p. Dryas, Vaccinium) и только простые (р.р. Pentaphylloides, Salix) (рис. 4). Межсосудистая поровость тоже представлена несколькими типами, как примитивными (лестничной, переходной в сочетании с супротивной), так и более специализированными (супротивной, очередной и смешанной) в различных сочетаниях у отдельных видов; а представители р.р. Empetrum, Vaccinium и Ribes обладают всеми перечисленными типами поровости (рис. 5); исключительно очередной - сомкнутой, сближенной и свободной
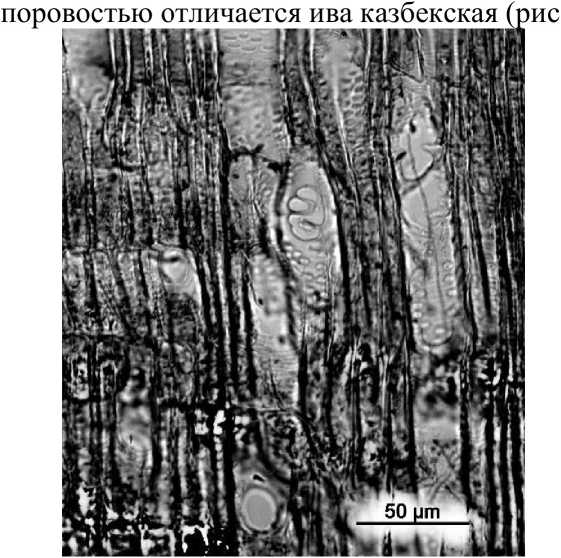
Рис. 4 . Vaccinium myrtillus - простая и нерегулярная перфорации

Рис. 5 . Ribes orientate — членики сосудов с лестничными перфорациями (с бифуркациями) и с межсосудистой поровостью: лестничной, переходной в сочетании с супротивной, супротивной, очередной и смешанной (радиальный срез)

Рис. 6. Salix kazbekensis - членики сосудов с простыми перфорациями, клювиками и очередной сближенной и сомкнутой межсосудистой поровостью (радиальный срез)
Волокнистые трахеиды, так же как и сосуды, в основном тонкостенные и занимают наибольший объем древесины. Окаймленные поры на их стенках с овальными и овально-вытянутыми апертурами. Встречаются также более толстостенные волокнистые трахеиды со слабо выраженными окаймлениями и щелевидными апертурами. У Ribes orientale встречаются волокна с порами, имеющими неясное окаймление и узкие отверстия, волокнистые трахеиды иногда образуют скопления в поздней части годичного кольца.
Аксиальная паренхима в основном скудная, апотрахеальная диффузная. Отдельные ее клетки или несколько клеток в коротких цепочках разбросаны в беспорядке среди волокнистых элементов. Лишь представители родов Dryas и Pentaphylloides имеют, кроме того, скудную паратрахеальную вазицентрическую паренхиму. Подавляющее большинство рассмотренных видов имеет гомогеннопалисадные и гетерогенные лучи (рис. 7).

Рис. 7 . Ribes orientate - гетерогенный луч (радиальный срез)
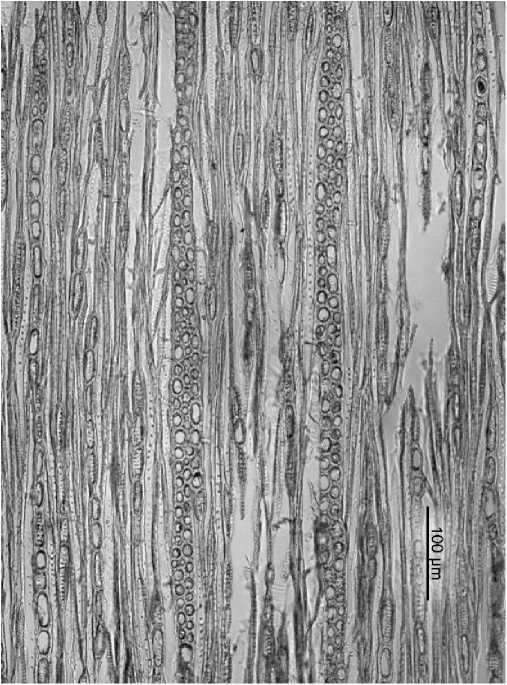
Рис. 8 . Rhododendron caucasicum - однорядные, двурядные и трехрядные лучи, последние - слившиеся короткими однорядными окончаниями (тангентальный срез)
Ширина (рядность) лучей исследованных видов достаточно разнообразна: от однорядных (р. Salix} и однорядных с несколькими двурядными слоями (р. Empetrum) или однорядных и двурядных (р. Pentaphylloides} до однорядных в сочетании с двурядными, трехрядными и многорядными (p.p. Dryas, Ribes, Vaccinium, Rhododendron} (рис. 8).
Высота лучей тоже демонстрирует значительный диапазон, являясь нестабильным признаком даже в пределах образца: наименьшей слойностью отличаются виды р. Empetrum - до 10 клеток, редко выше; у p.p. Dryas, Ribes, Vaccinium, Salix - до 30-35 клеток. Отмечено, что у видов, имеющих узкие лучи, часто происходит слияние их концами, что значительно увеличивает их слойность. У двутрехрядных и многорядных лучей имеются длинные и короткие однорядные окончания (p.p. Dryas и Pentaphyllo ides}; у p.p. Vaccinium и Ribes - лучи с короткими однорядными окончаниями или без них.
Эколого-ксилотомический анализ аэроксильных и простратных кустарников и кустарничков альпийского пояса восточных районов Северного Кавказа показал невысокий уровень специализации их вторичной ксилемы, имеющей ряд примитивных признаков. В гидросистеме этих растений еще нет четкого структурно-функционального деления на водопроводящие, механические и запасающие элементы. Об этом свидетельствует присутствие в древесине всех исследованных видов (за исключением Salix kazbekensis} волокнистых трахеид, выполняющих одновременно проводящую и механическую функции. На эту же черту указывают и отмеченные у подавляющего числа видов рассеяннососудистый и полукольцесосудистый типы древесины. Наиболее специализированный кольцесосудистый тип, с его повышенной способностью к проведению воды, по-видимому в особо холодных местах обитания не требовался. К тому же для формирования такой ксилемы необходимо значительно больше времени и энергетических ресурсов, чем двум другим типам. (Лобжанидзе., Габуния, 2008; Чавчавадзе, Сизоненко, 2002; Чавчавадзе и др., 2004).
Для экстремальных условий высокогорий характерно отложение в стеблях растений очень узких, иногда множественных («ложных») слоев прироста, часто с неясными границами. Это явление обусловлено периодичностью работы камбия в течение короткого вегетационного сезона, низкими температурами, неустойчивой погодой с частыми снегопадами и сильными холодными ветрами. Сужение годичных колец связано с некоторыми адаптивными перестройками в их структуре. Чаще всего оно вызывает увеличение густоты сосудов и некоторое уменьшение их диаметров, что сохраняет водопроводимость на том же уровне. Вторичной ксилеме альпийских растений присущи частые сочетания одиночных и сгруппированных сосудов очень малого и малого диаметра с количественным перевесом более примитивных -одиночных (до 75-80 %). Узкие просветы сосудов усиливают связь водных нитей со стенками, снижая опасность эмболии, а их полигональная форма увеличивает число контактов с окружающими клетками древесины (Carlquist, 1984; Чавчавадзе, Сизоненко, 2002).
Для нормального функционирования водопроводящей системы наиболее существенное значение имеют структурные особенности перфорационных пластинок и межсосудистой поровости. У изученных видов кустарников и кустарничков можно встретить все существующие типы перфораций - от наиболее архаичных (лестничных с большим количеством бифуркаций и сетчатых) до наиболее специализированных - простых. Одним типом перфораций обладают виды (p.p. Empetrum, Rhododendron, Pentaphylloides и Salix). Одновременное присутствие у члеников сосудов лестничных и простых перфораций отмечается у р.р. Dryas и Vaccinium, а р. Ribes обладает различными типами перфораций и их комбинациями. Такое сочетание физиологически выгодно альпийским растениям, так как специализированные простые перфорации повышают эффективность водопроведения, а примитивные - лестничные и сетчатые - задерживают и рассеивают пузырьки воздуха, образующиеся при таянии снега и оттаивании почвы, защищая этим сосуды от закупорки.
Достаточная абсолютная и относительная влажность альпийских биотопов позволяет существовать растениям с довольно примитивной лестничной, переходной и точечной - супротивной поровостью члеников сосудов (например, виды р.р. Rhododendron, Vaccinium). Одновременное присутствие на стенках сосудов лестничной и точечной - супротивной, очередной и смешанной поровости значительно расширяет экологический диапазон растений, позволяя им продвигаться выше, к границе нивального пояса (р.р. Dryas, Pontaphylloides, Empetrum), а также осваивать осыпи и маренные субстраты (р.р. Dryas, Ribes). Ива казбекская, обладающая самой специализированной межсосудистой поровостью (очередной -сомкнутой, сближенной и свободной) в условиях альпийского пояса особых преимуществ не имеет.
Аксиальная паренхима альпийских растений, выполняющая в основном запасающую функцию, скудная. Некоторое исключение представляют виды р.р. Dryas и Pontaphylloides, имеющие как мало специализированную апотрахеально-маргинальную паренхиму, так и более эволюционно продвинутую - паратрахеально-вазицентрическую. Однако, обильной тяжевой паренхимы у альпийских древесных видов нет. Частично запасающую функцию берут на себя радиальные лучи и, в первую очередь, гомогенно-палисадного типа, которые наблюдаются во вторичной ксилеме всех исследованных видов. Они предназначены преимущественно для взаимодействия с осевыми элементами древесины и необходимы видам с особенно скудной аксиальной паренхимой. Характерные для арборифлоры высокогорий гетерогенные лучи выполняют одновременно проводящую и запасающую функции.
Заключение. Приведенные выше структурные особенности вторичной ксилемы, характеризуют альпийские кустарники и кустарнички восточной части Северного Кавказа как типичных мезофитов. Можно предположить, что их предки, относящиеся к тем же родам и семействам, сформировались в мезофильных лесных ценозах и оказались преадаптированными к условиям высокогорий. Дальнейшие приспособления к суровой обстановке альпийского пояса были связаны с некоторыми перестройками в их вегетативной сфере, в том числе, в гидросистеме, которые носили в основном количественный характер, не затрагивая существенно типичное строение их древесины.
У всех рассмотренных видов, независимо от их систематического положения, в структуре водопроводящей ткани отмечено присутствие признаков разного уровня специализации (гетеробатмия), что указывает скорее на их физиологическую необходимость, чем на эволюционный уровень таксона (Чавчавадзе, Сизоненко, 2002). Этот вывод подтверждается исследованиями некоторых ученых, считающих, что господство различных семейств цветковых растений в экстремальных условиях крайне холодных регионов объясняется не их положением в системе, а некоторыми географическими и биологическими причинами (Хохряков, 1981; Хохряков, 1984).
Авторы статьи выражают искреннюю признательность д.б.н. Гайсумову М.А. и Трухмановой Г.Р.
ВОЛКОВА Светлана Борисовна - научный сотрудник отдела Ботанический музей, ФГБУН Ботанический институт им. В. Л. Комарова РАН, 197376, Санкт-Петербург, ул. проф. Попова, д. 2, e-mail: vsb 105 @yandex.ru
Чавчавадзе Е.С. Особенности адаптации вторичной кислемы кустарников и кустарничков Альпийского пояса (Северный Кавказ) / Е.С. Чавчавадзе, М.У. Умаров, О.Ю. Сизоненко, С.Б. Волкова // Вести. ТвГУ. Сер.: Биология и экология. 2017. № 3. С. 100-111.
Список литературы Особенности адаптации вторичной ксилемы кустарников и кустарничков к условиям Альпийского пояса (Северный Кавказ)
- Атлас древесины и волокон для бумаги. 1992/ред. Чавчавадзе Е.С. М.: Ключ. 336 с.
- Бенькова В.Е., Швейнгрубер Ф.Х. 2004. Анатомия древесных растений России. Берн-Штутгарт-Вена: Поль Хаупт. 456 с.
- Галушко А.И. 1975. Растительный покров Чечено-Ингушетии. Грозный: Чечено-Ингушское книжное изд-во. 118 с.
- Галушко А.И. 1978. Флора Северного Кавказа. Грозный: Изд-во Ростовского ун-та. 320 с.
- Лобжанидзе Э.Д., Габуния М.Д. 2008. Экология камбиальной активности и формирования древесины. Тбилиси: Изд-во «Ламари». 513 с.
- Умаров М.У. 1992. Пути адаптации водопроводящей ткани древесно-кустарничковых растений к горным местообитаниям: дис. … д-ра. биол. наук. Л. 317 с.
- Умаров М.У., Чавчавадзе Е.С., Волкова С.Б. 2007. К методике создания информационного банка структуры древесины покрытосеменных//Горные экосистемы и их компоненты. Нальчик. 13-18 августа 2007 г.: труды международной конф. М.: Товарищество науч. изд. КМК. С. 143-147.
- Умаров М.У., Чавчавадзе Е.С., Сизоненко О.Ю. 2008. Арборифлора Чеченской республики (Восточный Кавказ), рациональное использование и охрана//Экологическая ситуация на Северном Кавказе, проблемы и пути их решения: материалы Всероссийской науч.-практич. конф. Нальчик. 2008. С. 84-103.
- Хохряков А.П. 1981. Эволюция биоморф растений. М.: Наука. 168 с.
- Хохряков А.П. 1984. Таксономический состав и основные направления эволюции растений Арктики. -в кн.: Адаптация организмов к условиям Крайнего Севера. СССР, Таллин, изд-во АН ЭССР. 1984. С. 188-192.
- Чавчавадзе Е.С., Сизоненко О.Ю. 2002. Структурные особенности древесины кустарников и кустарничков Арктической флоры России. СПб.: Росток. 272 с.
- Чавчавадзе Е.С., Умаров М.У., Сизоненко О.Ю. 2004. Пути адаптации покрытосеменных России к условиям высокогорий и высоких широт//Труды IV международного симпозиума «Строение, свойства и качество древесины». СПб. С. 143-145.
- Энциклопедия географических терминов. 1968. М. Советская энциклопедия. 435 с.
- Яценко-Хмелевский А.А 1946. Строение древесины кавказских представителей сем.Ericaceae и его систематическое и филогенетическое значение//Изв. АН Арм. ССР. № 9. С. 33-58.
- Яценко-Хмелевский А.А. 1954а. Основы и методы анатомического исследования древесины. М. 337 c
- Яценко-Хмелевский А.А. 1954б. Древесины Кавказа. Т. 1. Ереван. 674 с.
- Carlquist S. 1984. Vessel grouping in dicotyledon wood: significance and reliantionship to imperfotate tracheary elements//Aliso. V. 10. № 4. P. 505-525.
- Carlquist S. 1988. Comparative wood anatomy: systematic, ecological and evolutionary aspects of dicotyledon wood. Berlin; Heidelberg; New York; London; Paris: Tokyo. 436 p.
- Metcalfe C. R., Chalk L. 1983. Anatomy of the Dicoyledons. Wood structure and conclusion of general introduction. Oxford. V. 2. 297 p.
- Weeler E.A., Baas P. 1998. Wood identification -a review//IAWA J. V. 19. P. 241-264.
