Особенности антэкологии и семенной продуктивности пустырника сизого и пустырника пятилопастного

Автор: Демьянова Евгения Ивановна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 1, 2012 года.
Бесплатный доступ
Рассмотрены особенности антэкологии и семенной продуктивности пустырника сизого (Leonurus glaucescens) и п. пятилопастного (L. quinquelobatus) в уральском регионе. Оба вида обладают высокой семенной продуктивностью и могут быть рекомендованы для выращивания в культуре как ценные лекарственные растения.
Антэкология, семенная продуктивность, системы скрещивания, лекарственные растения
Короткий адрес: https://sciup.org/147204576
IDR: 147204576 | УДК: 582.4/.9-15
Peculiarities of anthecology and seed production of leonurus glaucescens and Leonurus quinquelobatus
Anthecology and seed production of two species of Leonurus L. have studied. We found that the real production of both species is very high. The breeding systems combines the cross-pollination and self-pollination.
Текст научной статьи Особенности антэкологии и семенной продуктивности пустырника сизого и пустырника пятилопастного
Оба исследованных вида пустырника ( Leonurus glaucescens Bunge и L. quinquelobatus Gilib., La-miaceae ) относятся к растениям с обширным ареалом [Куприянова, 1954]. Указанные виды несколько различаются по характеру местообитаний: L. quinquelobatus присуще большее тяготение к сорным местообитаниям в сравнении с L. glaucescens , произрастающим по кустарникам, оврагам и степям и, редко, как сорное. Виды пустырника являются лекарственными, медоносными и жирномасличными растениями [Дикорастущие…, 2001; Зи-эп, Жохова, 2007 и др.]. Для получения лекарственного сырья используются верхние цветущие части побегов как у дикорастущих, так и культивируемых растений. Для практических целей у последних легче организовать заготовку сырья, а также сбор семян для дальнейшего расширения плантаций видов пустырника.
Задача исследования – изучение систем скрещивания у видов пустырника как важных офици-нальных растений для рекомендации их выращивания в уральском регионе. Литературные сведения о подобных опытах в отношении видов пустырника единичны [Паутова, Джумаев, 1986].
Организация исследований
Наблюдения за экологией цветения, опыления и семенной продуктивностью пустырника сизого проведены в 1986–1987 и 2010 гг. в Троицком лесостепном заказнике (Челябинская обл.) в зарослях ивняка по берегу оз. Кукай. Пустырник пятилопастной исследовался в 1995, 2004–2005, 2010–2011 гг. в Пермском крае (Нытвенский р-н), где он про- израстает на сорных местах, и на коллекционном участке лекарственных растений Учебного ботанического сада Пермского университета.
Антэкологические наблюдения проведены по методике А.Н. Пономарёва [1960]. Они сопровождались измерением температуры и влажности воздуха на уровне соцветий аспирационным термометром Асмана. Фертильность пыльцы определялась ацето-карминовым методом [Паушева, 1980; Справочник…, 2004]. Семенная продуктивность изучена по методике И.В. Вайнагия [1973, 1974] с раздельным определением потенциальной и реальной (фактической) семенной продуктивности (ПСП и РСП) в расчёте на 1 цветок, на 1 генеративный побег или на 1 особь. Поскольку число генеративных побегов у изучаемых растений зависит от их возраста и условий местообитания, такое положение вызывает затруднения при выборе расчётной единицы. Для удобства интерпретации полученного материала расчёт семенной продуктивности производился не на 1 особь, а только на генеративный побег. Под ПСП, следуя И.В. Вайнагию [1973, 1974], понимали число семяпочек в расчёте на 1 цветок. В семействе губоцветные этот показатель строго детерминирован и равен 4. РСП – число семян (эремов) в расчёте на 1 цветок. В дальнейшем эти показатели семенной продуктивности в расчёте на 1 цветок перемножались на 1 генеративный побег с учётом на нём числа мутовок и числа цветков в них. Для характеристики эффективности завязывания семян использовали показатель ПС – коэффициент семенификации, вычисляемый как РСП/ПСП∙100. Определение семенной продуктивности проводилось в двух вариантах: при свободном опылении и изоляции генеративных побегов.
Последние в стадию бутонизации покрывались марлевыми мешочками во избежание посещений насекомыми. Проращивание семян проводилось в лабораторных условиях на увлажнённой фильтровальной бумаге в чашках Петри [Николаева, Разумова, Гладкова, 1985; Николаева, Лянгузова, Поздова, 1999; Ишмуратова, Ткаченко, 2009] при температуре 20– 22°С, на свету. Энергия прорастания учитывалась на 3–5-й день. Семена проращивали до 30–40 дней, а затем подводились итоги по следующим параметрам: проросшие, твёрдосемянные (непроросшие, но не заплесневевшие) и сгнившие семена. Цифровой материал подвергнут статистической обработке [Лакин, 1980].
Результаты и их анализ
Leonurus glaucescens Bunge – пустырник сизый
Начало цветения этого вида в условиях лесостепного Зауралья в 1986–1987 гг. отмечено 10–14 июля, а конец фиксирован последними числами августа. В зависимости от габитуса растения, что в значительной степени определяется его жизненной формой (двулетник или многолетний поликарпик) и условиями местообитания, цветение особи составляет 1.5–2 мес.
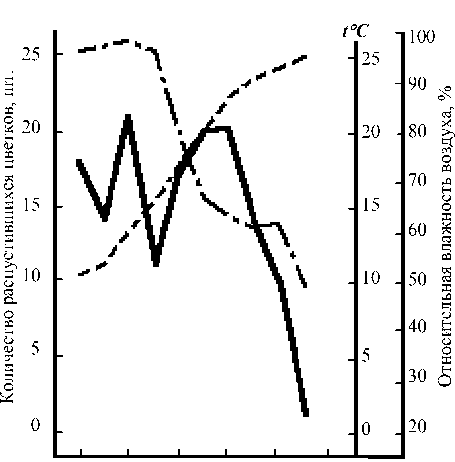
Наблюдения за суточной ритмикой цветения проводили трижды – 4, 6 и 7 июля 1988 г. (рис. 1–3). В эти дни погода была умеренно прохладной, ветреной, довольно облачной. В утренние часы отмечалась обильная роса. Начало распускания цветков очевидно было приурочено к раннеутренним часам, поскольку ко времени наших наблюдений (5 ч.) око-
Время 5.00 7.00 9.00 11.00 13.00 15.00
Рис. 1. Суточный ритм распускания цветков Leonurus glaucescens 4 июля 1987 г.
Условные обозначения:
– кол-во распустившихся цветков
– температура воздуха
– относительная влажность воздуха
5.00 7.00 9.00 11.00 13.00
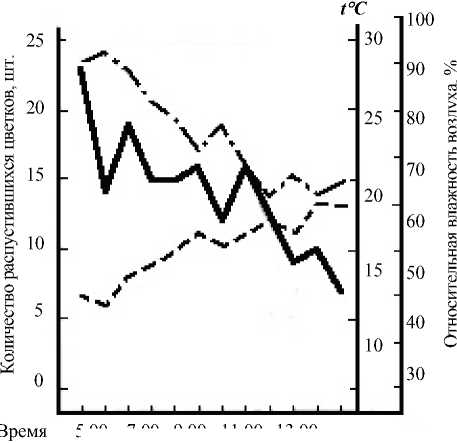
Рис. 2. Суточный ритм распускания цветков Leonurus glaucescens 6 июля 1987 г.
Условные обозначения те же, как на рис. 1
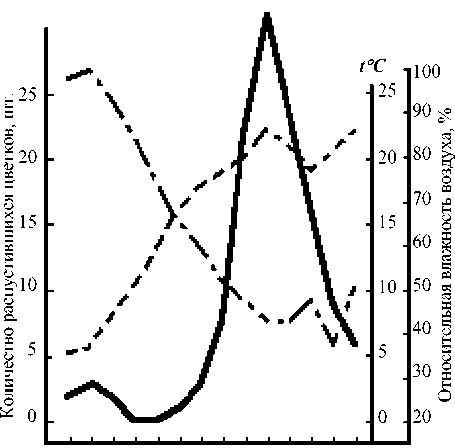
Время 5.00 7.00 9.00 11.00 13.00 15.00 17.00
Рис. 3. Суточный ритм распускания цветков Leonurus glaucescens 7 июля 1987 г.
Условные обозначения те же, как на рис. 1
ло 12–13% цветков от суммы цветков, распустившихся за день, оказывались уже раскрытыми. Цветение прекращалось к 14 ч. при сравнительно ясной погоде (4 июля) и продолжалось до 16–17 ч. – при пасмурной (6 июля). В последнем случае пик цветения не был ярко выражен и распускание цветков происходило достаточно равномерно. Иной оказалась динамика цветения 7 июля, что, вероятней всего, было связано с низкой утренней температурой. В этот день раскрывание цветков началось несколько позже обычного и продолжалось до 18 ч. Массовое распускание цветков наблюдалось между 13–16 ч.
(раскрылось около 70% цветков от общей суммы всех цветков, распустившихся за день). Пик цветения был приурочен к 14 ч. На основании наших наблюдений можно констатировать наличие у пустынника сизого дневного типа распускания цветков.
Наблюдения за экологией цветка позволили выявить следующие закономерности. Распускание цветков в пределах главного соцветия идёт в акропетальном порядке. Цветки дихогамны, протанд-ричны. 4 тычинки расположены парами: у верхних тычинок короткие тычиночные нити, у нижней пары, напротив, тычиночные нити длинные. Вскрывание пыльников интрорзное. Обычно первыми растрескиваются пыльники нижней пары тычинок, а затем через короткий промежуток времени – верхние. В это время рыльце едва заметно в зеве цветка, поскольку столбик ещё короткий. Оно находится в слабо выраженной вильчатой стадии. Воспринимающая сосочковая поверхность рыльца в это время ещё плохо выражена. Едва заметные сосочки покрывают лишь кончики лопастей рыльца. При дальнейшем выдвижении из околоцветника рыльце, находясь уже в отчётливо выраженной вильчатой стадии и поднимаясь к верхней губе, минует вскрытые пыльники верхних тычинок, не приходя с ними в соприкосновение. Однако вероятность спонтанного попадания пыльцы на рыльце достаточно велика, и она связана с наложением рыльцевой фазы на ещё не закончившуюся тычиночную. Физиологическая зрелость рыльца, определяемая возможностью прорастания на нём пыльцы, наступает примерно через 14–16 ч. после раскрывания цветков. В это время на рыльце хорошо просматриваются сосочки, а его лопасти широко расставлены. Тычиночная стадия продолжается примерно в течение 28–30 ч., что и гарантирует совмещение обеих фаз цветения. Самоопылению весьма способствуют и многочисленные насекомые (шмели и пчёлы), интенсивно посещающие нектароносные цветки пустырника [Карташова, 1965; Глухов, 1974; Пельменев, 1985, и др]. Передвигаясь по густому соцветию, они неизбежно способствуют самоопылению в форме гейтоногамии (при наличии самосовместимости у этого вида и высокой фертильности пыльцы; последняя во все годы исследований была более 90%).
После засыхания тычинок лопасти рыльца приобретают ещё более выраженную вильчатую форму, и при этом столбик начинает отклоняться к отогнутой верхней губе. Рыльцевая стадия продолжается около 30 ч., т.е. её длительность сопоставима с тычиночной. Наши наблюдения за антэ-кологическими особенностями пустырника сизого позволяют говорить о возможности самоопыления у этого вида, что значительно облегчается совпадением тычиночной и рыльцевой стадий цветения. При изоляции генеративных побегов отмечается массовое завязывание семян.
Семенная продуктивность . Как указывалось ранее, изучение семенной продуктивности пустырника сизого проводилось в двух вариантах опыта: при свободном опылении и изоляции. В первом варианте опыта обнаружена высокая семенная продуктивность этого вида в расчёте на 1 цветок и на 1 генеративный побег, что объясняется высокой фертильностью пыльцы и хорошей посещаемостью цветков насекомыми (табл. 1, 2; рис. 4). На эффективность завязывания семян указывает и высокий процент семенификации. В структуре семенной продуктивности решительно преобладают цветки, завязавшие по 4 эрема (85.4%).
Таблица 1
Семенная продуктивность пустырника сизого при свободном опылении в 1986 г.
|
Кол-во исследованных цветков |
Показатели семенной продуктивности |
||||
|
ПСП (семяпочки), шт. |
РСП (семена), шт. |
ПС, % |
СП на 1 цветок, шт. |
||
|
М ± m |
сv, % |
||||
|
500 |
2000 |
1949 |
97.43 |
3.89 ± 0.02 |
11.81 |
Иная картина наблюдается в опыте с изоляцией генеративных побегов (табл. 2). Здесь все показатели семенной продуктивности уступают варианту свободного опыления. Прежде всего, при изоляции обнаруживается большое количество цветков, не завязавших ни одного эрема (рис. 4). В этом варианте обнаружен и большой процент неполноценных зачатков двух типов, которые обозначаются как «невыполненные» и «пустые». К первой категории отнесены мелкие и щуплые зачатки, резко отличающиеся по внешнему виду от полноценных эремов. Однако обращает на себя внимание тот факт, что количество невыполненных семян примерно одинаково при обоих вариантах скрещива- ния. В такой ситуации можно говорить о видоспецифичной особенности пустырника сизого, определяемой внутренними факторами, и не зависящей от способов опыления.
Наличие «пустых» семян у губоцветных впервые отмечено Р.Е. Левиной [1970] для Salvia pra-tensis L. и Stachys recta L. «Пустые» зачатки по размерам не отличимы от полноценных соответствующей фазы спелости. Только при разрушении околоплодника можно убедиться, что они лишены содержимого. Вместо семени в них находится сухая бурая плёнка или личинка насекомого. Подобные зачатки наблюдали и мы. При поражении семян насекомыми в околоплоднике имелось не- большое отверстие. Такие зачатки учитывались нами как «повреждённые». Следует отметить, что количество «повреждённых» эремов в большей степени наблюдается при свободном опылении.
По нашему мнению, это связано с тем, что при изоляции затруднён доступ вредных насекомых, повреждающих семена.
Таблица 2
Семенная продуктивность пустырника сизого при разных вариантах опыления в 1987 г. (исследовано по 500 цветков)
|
Показатель |
Вариант опыта |
|
|
свободное опыление |
изоляция |
|
|
ПСП в расчете на 1 цветок |
4 |
4 |
|
РСП в расчете на 1 цветок |
3.58 ± 0.05 |
2.84 ± 0.08 |
|
сv, % |
31.90 |
62.73 |
|
td, % |
7.78 |
|
|
кол-во мутовок на 1 ген. побег, шт.1 |
13.56 ± 0.44 |
13.56 ± 0.44 |
|
кол-во цветков в мутовке, шт.1 |
15.15 ± 0.19 |
15.15 ± 0.19 |
|
кол-во цветков на 1 ген. побег, шт.1 |
205.43 ± 7.15 |
205.43 ± 7.15 |
|
ПСП в расчете на 1 ген. побег, шт1 |
821.74 ± 23.07 |
821.74 ± 23.07 |
|
РСП в расчете на 1 ген. побег, шт. |
735.85 ± 27.67 |
584.24 ± 26.14 |
|
td, % |
3.98 |
|
|
ПС в расчете на 1 ген. побег, % |
89.55 |
71.10 |
Примечание. 1 Эти показатели мы рассматривали как общие для обоих вариантов опыта.
В 2010 г. в этом же районе и в тех же местообитаниях у 250 цветков при свободном опылении
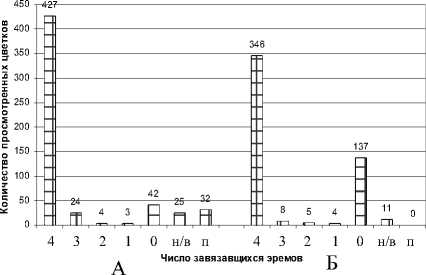
Рис. 4. Структура семенной продуктивности Leonurus glaucescens при разных вариантах опыления в 1987 г.: А – при свободном опылении, Б – при изоляции.
Условные обозначения: н/в – невыполненные эремы, п – поврежденные эремы исследовалась семенная продуктивность пустырника сизого в расчёте на 1 цветок. Несмотря на крайне засушливый летний сезон этого года, она оказалась весьма высокой (3.45 ± 0.03) и близкой к величине, полученной нами ранее.
Всхожесть семян у данного вида определялась в 1986–1987 гг., причём в последний год она изучена для семян, полученных при свободном опылении и при изоляции генеративных побегов (табл. 3). Результаты опытов свидетельствуют о высокой энергии прорастания и всхожести семян в обоих вариантах опыта, при этом у изолированных растений отмечены даже более высокие показатели. Вероятно, это связано с отсутствием повреждения семян насекомыми при изоляции генеративных побегов.
Таблица 3 Всхожесть семян Leonurus glaucescens , полученных при свободном опылении и изоляции в 1986–1987 гг.
|
Вариант опыта |
Показатели всхожести семян |
||||||
|
энергия прорастания, % |
среднее значение всхожести семян на 30-й день, % |
td, % |
среднее значение непроросших эремов, % |
среднее значение заплесневевших семян, % |
среднее значение “пустых” семян, % |
||
|
на 3-й день |
на 5-й день |
||||||
|
1986 г. |
|||||||
|
свободное опыление |
80.21 |
9.42 |
93.41 ± 0.92 |
0 1 |
2.43 |
4.16 |
0 |
|
1987 г |
|||||||
|
свободное опыление |
16.22 |
54.21 |
76.61 ± 2.41 |
2.08 |
0 |
9.61 |
13.78 |
|
изоляция |
48.24 |
36.62 |
85.63 ± 3.62 |
2.24 |
8.01 |
4.12 |
|
Leonurus quinquelobatus Gilib. – пустырник пятилопастной
По антэкологическим характеристикам (дневному суточному ритму цветения, экологии цветка, высокой фертильности пыльцы и другим призна- кам) этот вид пустырника практически не отличается от предыдущего. По габитусу особи п. пятилопастного (многолетнего поликарпика), как правило, представляют собой более крупные растения с большим количеством боковых побегов и цветков на особи в сравнении с п. сизым. Оба вида весьма близки и по семенной продуктивности при свободном опылении (табл. 4; рис. 5).
Таблица 4
Семенная продуктивность Leonurus quinquelobatus при свободном опылении в разные годы наблюдений
|
Показатель семенной продуктивности |
Годы наблюдений |
||||
|
1995 |
2004 |
2005 |
2010 |
2011 |
|
|
Среднее число мутовок на 1 ген. побег, шт. |
17.14 ± 1.33 |
12.63 ± 0.91 |
18.33 ± 0.54 |
17.93 ± 2.43 |
18.14 ± 1.34 |
|
Среднее число цветков в мутовке, шт. |
28.71 ± 2.23 |
22.72 ± 0.53 |
26.63 ± 0.31 |
23.13 ± 1.51 |
25.71 ± 2.14 |
|
Среднее число цветков на 1 ген. побег, шт. |
489.61 ± 0.13 |
287.15 ± 20.12 |
486.13 ± 24.03 |
414.03 ± 0.23 |
465.52 ± 0.13 |
|
ПСП на 1 цветок, шт. |
4 |
4 |
4 |
4 |
4 |
|
ПСП на 1 ген. побег, шт. |
1958.91 ± 0.43 |
1146.14 ± 80.23 |
1944.21 ± 97.24 |
1656.11 ± 0.62 |
1862.31 ± 0.43 |
|
РСП на 1 цветок, шт. |
3.95 ± 0.01 |
3.97 ± 0.01 |
3.97 ± 0.01 |
3.99 ± 0.02 |
3.91 ± 0.01 |
|
РСП на 1 ген. побег, шт. |
1939.91 ± 0.03 |
1137.52 ± 0.07 |
1929.62 ± 77.24 |
1655.32 ± 0.03 |
1820.23 ± 0.03 |
|
ПС на 1 цветок, % |
98.80 |
99.30 |
99.30 |
99.50 |
97.80 |
|
ПС на 1 ген. побег, % |
99.10 |
99.20 |
99.20 |
99.90 |
97.80 |
|
ПП на 1 ген. побег, % |
100 |
98.40 |
98.80 |
100 |
99.20 |
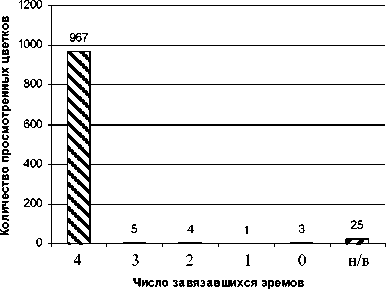
Рис. 5. Структура семенной продуктивности Leonurus quinquelobatus при свободном опылении в 2005 г. Условные обозначения те же, как на рис. 4
Всхожесть семян пустырника пятилопастного, полученных при свободном опылении, примерно такая же, как и для п. сизого (ср. табл. 3 и 5). При сравнении всхожести семян при двух вариантах опыления (табл. 5) обращают на себя внимание более низкие показатели, свойственные варианту изоляции: здесь семена прорастали практически в два раза хуже в сопоставлении с вариантом свободного опыления. Показатель энергии прорастания семян свидетельствует о более дружном их прорастании на 3-й день наблюдений у семян, полученных при свободном опылении. Семена, полученные при изоляции, в большей степени подвержены гниению (заплесневению). Что касается непроросших семян, то их большее количество также в варианте самоопыления.
Таблица 5
Всхожесть семян Leonurus quinquelobatus , полученных при свободном опылении и изоляции в разные годы наблюдений
|
Вариант опыта |
Показатели всхожести семян |
|||||||
|
энергия прорастания, % |
среднее значение всхожести семян на 30-й день, % |
td, % |
среднее значение непроросших эремов, % |
td, % |
среднее значение заплесневевших семян, % |
среднее значение «пустых» семян, % |
||
|
на 3-й день |
на 5-й день |
|||||||
|
2006 г. |
||||||||
|
свободное опыление |
83.62 |
5.63 |
89.31 ± 1.72 |
0 1 |
10.69 ± 0.01 |
0 1 |
0 1 |
0 |
|
2010 г. |
||||||||
|
свободное опыление |
63.33 |
15.12 |
78.31 ± 1.21 |
0 1 |
21.71 ± 1.62 |
0 1 |
0 1 |
0 |
|
2011 г |
||||||||
|
свободное опыление |
51.33 |
26.26 |
83.0 ± 1.72 |
7.55 |
17.0 ± 1.52 |
6.47 |
0 |
0 |
|
изоляция |
8.67 |
25.33 |
40.33 ± 1.78 |
30.0 ± 1.32 |
29.67 |
0 |
||
Заключение
Оба изученных растения (Leonurus glaucescens и L. quinquelobatus) оказались весьма близки по антэ-кологическим особенностям. Это энтомофильные растения с дневной ритмикой распускания цветков. Последние дихогамны, протандричны, с высокой фертильностью пыльцы. Тем не менее, протандрия не предотвращает совмещение тычиночной и рыль-цевой стадий, что способствует самоопылению в пределах цветка и соцветия и обеспечивает высокую семенную продуктивность как при свободном опылении, так и при изоляции. Подобное положение, вероятно, связано с их приуроченностью к рудеральным местообитаниям.
Согласно литературным сведениям, у сорных растений широко представлено самоопыление в разных вариантах: автогамия, гейтоногамия и клейстогамия [Anderson, 1989; Butterfield, Meshaka, Trauth, 1989; Travers, Mena-Ali, Stephenson, 2004, и др.]. По мнению указанных авторов, у сорняков са-монесовместимость обычно отсутствует, поскольку нарушенные местообитания требуют частой реколонизации. К тому же такие местопроизрастания кратковременны, а размеры популяций весьма невелики. Оба исследованных нами сорных вида имеют вполне устойчивую систему скрещивания, успешно сочетающую самоопыление и перекрёстное опыление. Они легки в культуре и могут быть рекомендованы в качестве официнарных растений для выращивания в культуре в уральском регионе.
Список литературы Особенности антэкологии и семенной продуктивности пустырника сизого и пустырника пятилопастного
- Вайнагий И.В. Методика статистической обработки материала по семенной продуктивности растений на примере Potentilla aurea L.//Раст. ресурсы. 1973. Т. 9, № 2. С. 287-296.
- Вайнагий И.В. О методике изучения семенной продуктивности растений//Бот. журн. 1974. Т. 59, № 6. С. 826-831.
- Глухов М.М. Медоносные растения. М.: Колос, 1974. 303 с.
- Дикорастущие полезные растения России/под ред. А.Л. Буданцева, Е.Е. Лесиовской. СПб.: Изд-во СПХФА, 2001. 663.
- Зиэп Т.Т., Жохова Е.В. Продукты вторичного метаболизма видов рода Leonurus (Lamiaceae)//Раст. ресурсы. 2007. Т. 43, № 4. С. 125-140.
- Ишмуратова М.М., Ткаченко К.Г. Семена травянистых растений. Особенности латентного периода, использование в интродукции и размножении in vitro. Уфа: Гилем, 2009. 115 с.
- Карташова Н.Н. Строение и функция нектарников цветка двудольных растений. Томск: Изд-во Томск. ун-та, 1965. 192 с.
- Куприянова Л.А. Род Leonurus L.//Флора СССР. 1954. Т. 21. С. 145-157.
- Лакин Г.Ф. Биометрия. М.: Высш. школа, 1980. 291 с.
- Левина Р.Е. К изучению ритма плодоношения некоторых губоцветных (Salvia pratensis L. и Stachys recta L.)//Бюл. МОИП. Отд. биол. 1970. Т. 75, № 3. С. 53-64.
- Николаева М.Г., Лянгузова И.В., Поздова Л.М. Био логия семян. СПб.: НИИ химии СПбГУ, 1999. 233 с.
- Николаева М.Г., Разумова М.В., Гладкова В.Н. Спра вочник по проращиванию покоящихся семян. Л.: Наука, 1985. 268 с.
- Паутова И.А., Джумаев Х.К. Некоторые особенности цветения и плодоношения Leonurus quinquelobatus Gilib.//Тр. I Молодеж. конф. ботаников г. Ленинграда. Л., 1986. С. 108-116.
- Паушева З.П. Практикум по цитологии растений. М.: Наука, 1980. 250 с.
- Пельменев В.К. Медоносные растения. М.: Россель-хозиздат, 1985. 142 с.
- Пономарёв А.Н. Изучение цветения и опыления растений//Полевая геоботаника. М.; Л., 1960. Т. 2. С. 9-19.
- Справочник по ботанической микротехнике. Основы и методы/состав. Р.П. Барыкина, Т.Д. Веселова, А.Г. Девятов, Х.Х. Джалилова, Г.М. Ильина, Н.В. Чубатова. М.: изд-во МГУ, 2004. 311 с.
- Флора СССР. М.; Л.: Изд-во АН СССР, 1954. Т. 21. 703 с.
- Andersson St. The evolution of self-fertility in Crepis tectorum (Asteraceae)//Plant Syst. and Evol. 1989. Vol. 168, № 3-4. P. 227-236.
- Butterfield B., Meshaka W.E., Trauth S.E. Evidence for predominant autogamy in Youngia japonica (Asteraceae)//Southwest. Natur. 1989. Vol. 34, № 4. P. 557-579.
- Mulligan G.A. Autogamy, allogamy and pollination in some Canadian weeds//Can. J. Bot. 1972. Vol. 50, № 8. P. 1767-1771.
- Travets S. E., Mena-Ali J., Stephenson A. Plasticity in the self-incompatibility system of Solanum carolinense//Plant Species Biol. 2004. Vol. 19, № 3. P. 127-135.
