Особенности биологии цветения Origanum vulgare L. (Lamiaceae) в Московской области
 в Московской области")
Автор: Годин Владимир Николаевич, Куранова Наталия Геннадиевна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 4, 2018 года.
Бесплатный доступ
Изучены структура синфлоресценций, особенности цветения общего и частного соцветий, а также обоеполых и пестичных цветков Origanum vulgare L. (Lamiaceae). Наблюдения проведены в 2015-2018 гг. по общепринятым методикам на территории Московской обл. Синфлоресценция O. vulgare - фрондозно-брактеозная щитковидная метёлка, состоящая из флоральных единиц - кистевидных тирсов. Обоеполые цветки O. vulgare строго протандричны, длительность их цветения составляет 2-3 дня. В пестичных цветках тычиночная стадия отсутствует, и они функционируют 1.5-2 дня. В кистевидных тирсах раскрывание цветков происходит акропетально, продолжительность цветения зависит от числа цветков в них. Зацветание флоральных единиц в пределах главного побега и паракладиев, а также самих паракладиев в составе синфлоресценции происходит в базипетальном порядке.
Синфлоресценция, биология цветения, протандрия, гинодиэция
Короткий адрес: https://sciup.org/147227039
IDR: 147227039 | УДК: 581.461: | DOI: 10.17072/1994-9952-2018-4-343-350
Flowering biology of Origanum vulgare L. (Lamiaceae) in Moscow region
The structure of synflorescences, the flowering of the main-florescences, co-florescences and partial florescences, as well as the perfect and pistillate flowers of Origanum vulgare L. (Lamiaceae) have been studied. Observations were conducted in 2015-2018 according to generally accepted methods in Moscow region. Synflorescence O. vulgare is a frondose-bracteate umbel-like panicle consisting of florale units are racemose thyrsus. The perfect flowers of O. vulgare are strictly protandrous, the duration of their flowering is 2-3 days. In the pistillate flowers, the staminate stage is absent, they function 1.5-2 days. In racemose thyrsus the opening of flowers occurs acropeately, the duration of their flowering depends on the number of flowers on them. Blooming of the floral units within the main axis and paraclades, as well as the paraclades in the synflorescence occurs in the basipetal order.
Текст научной статьи Особенности биологии цветения Origanum vulgare L. (Lamiaceae) в Московской области
Изучение особенностей цветения и опыления, выяснение типов и механизмов цветения цветка и соцветия, способов и характера опыления является необходимым условием для всестороннего исследования антэкологии любого вида растений [Пономарев, 1960]. Порядок раскрывания цветков как признак редко используется при описании соцветий, но изучать его необходимо, так как он имеет большое значение для познания закономерностей морфогенеза и эволюции соцветий [Sell, 1980; Кузнецова, 1991]. С другой стороны, последовательность цветения цветков в соцветии или синфлоресценции имеет и практическое значение для выявления возможности гейтоногамии в пределах как частного соцветия, так и со- цветий паракладиев и в целом объединённого соцветия.
В качестве объекта наших исследований выбрана душица обыкновенная ( Origanum vulgare L.) – коротко- или длиннокорневищный многолетний травянистый поликарпик, гемикриптофит или травянистый хамефит, обладающий евразиатским ареалом, распространённый преимущественно в различных вариантах лесостепи [Нухимовский, Черкасов, 1987]. Для O. vulgare , как и для многих представителей сем. Lamiaceae , характерна гино-диэция, проявляющаяся в образовании двух типов цветков – обоеполых и пестичных – на разных
особях в пределах популяции [Годин, 2011; Годин, Демьянова, 2013].
W. Troll [1964] показал, что все представители сем. Lamiaceae характеризуются полителическим (открытым) тирсом, составные части которого представляют собой цимоиды – цветоносные системы, состоящие из терминального цветка и 1 или 2 симподиально нарастающих боковых ветвей. Дальнейшие исследования представителей данного семейства продемонстрировали огромное разнообразие строения их флоральных единиц, выражающееся в варьировании числа паракладиев, порядке ветвления синфлоресценции, числа субъединиц в тирсе и т. д. [Буданцев, 1992; Байкова, 2006].
Данные в литературе о структуре соцветий O. vulgare и ее антэкологических особенностях фрагментарны, зачастую неоднозначны и иногда противоречивы. Так, Л.Б. Зимина [2002] характеризует цветоносные побеги данного вида как полители-ческую множественную кисть, состоящую из колосков. Е.Л. Нухимовский и О.А. Черкасов [1987] считают, что парциальное соцветие у O. vulgare представляет собой брактеозную кисть. Т.В. Кузнецова, Н.И. Пряхина, Г.П. Яковлев [1992] определяют синфлоресценцию O. vulgare как метелку из колосьев щитковидной или овальной формы. Согласно данным И.С. Шарыгиной [1959], продолжительность цветения обоеполых цветков O. vulgare составляет около 2–2.5 сут., однако по результатам исследований Л.Б. Зиминой и Н.Т. Ко-нон [1999] оно продолжается 7–9 сут., а у пестичных цветков – 4–6 сут. Перечисленные авторы к тому же не указывают наличие протандрии, характерной для многих представителей сем. Lamiaceae и для O. vulgare , в частности [Knuth, 1899]. В связи с этим цель нашей работы – изучение особенностей цветения O. vulgare.
Материал и методы исследования
Изучение антэкологических особенностей O. vulgare проводили в 2015–2018 гг. в окр. Агробиостанции МПГУ (пос. Павловская Слобода, Истринский р-он Московской обл.). Выявление особенностей цветения проводили по плану, предложенному А.Н. Пономаревым [1960].
В основе наших исследований лежали наблюдения, которые должны были ответить на поставленную цель. Прежде всего, нами изучены морфология и биология цветков. Анализ цветков проводили с учётом следующих особенностей: форма и окраска околоцветника и изменение этих признаков в течение жизни цветка, особенности строения андроцея и гинецея и их взаимное расположение, движение тычинок и рылец в процессе развития и функционирования цветка, время созревания рылец и пыльников, строение, положение и доступность нектарников для насекомых, специальные образования и структуры в цветке, способствующие опылению.
Для выявления морфологических адаптаций цветка к способу опыления подробно изучен морфогенез цветка, начиная от плотного бутона до увядания околоцветника и генеративных структур. В этом отрезке функционирования цветка выделены определённые стадии. В связи с наличием у O. vulgare двух типов цветков (обоеполых и пестичных) изучение биологии цветения проводили на особях разного полового статуса.
Длительность тычиночной фазы определяли визуально на 20 этикетированных цветках ежегодно, в течение четырех лет, на разных растениях двух половых форм. Началом тычиночной фазы считали момент растрескивания пыльников у 1–2 самых первых тычинок. Окончание фиксировали по времени раскрывания пыльников последней тычинки и высеивания из пыльников пыльцы.
Степень зрелости рыльцевой поверхности определяли как визуально по морфологическим признакам, так и с помощью химических методов. Окрашивание рыльца проводили по методу Робинсона [Robinsohn, 1924]. Суть данной реакции заключается в том, что воспринимающая поверхность зрелых рылец при нанесении на неё слабого раствора перманганата калия окрашивается в коричневый или бурый цвет. Незрелые рыльца перманганатом калия не окрашиваются. Позитивную реакцию зрелых рылец мы наблюдали также при окрашивании метиленовой синей в концентрации 1:20 000 (водный раствор).
По линии отдельного цветка отмечали кратность его раскрывания и продолжительность жизни раскрывшегося цветка при различных погодных условиях. Для изучения продолжительности жизни раскрывшегося цветка на маркированных растениях было взято не менее 50 цветков ежегодно, в течение четырех лет наблюдений.
При изучении цветения флоральных единиц и синфлоресценций в целом проводили ежедневный учет раскрывшихся и отцветших цветков на пяти особях O. vulgare , одновременно отмечали характер цветения флоральных единиц в разных частях синфлоресценции. Наблюдения проводили по схеме, представленной на рис. 1. Учет раскрывшихся цветков проводили на одной из симметричных сторон синфлоресценции. Полученные данные наблюдений сведены в виде графиков (рис. 2 и 3), соответствующих указанной схеме (рис. 1).
Результаты и их обсуждение
Структура синфлоресценции. Генеративные побеги O. vulgare имеют типичную структуру, включающую зоны возобновления, торможения и обогащения. В зоне возобновления развиваются почки возобновления и озимые побеги, в зоне тор- можения – короткие вегетативные силлептические побеги (в нижней части зоны они отсутствуют), зона возобновления состоит из главного соцветия, развитого на верхушке побега, и боковых соцветий, развитых на верхушках силлептических боковых побегов (паракладиев). У генеративных побегов O. vulgare отмечается разная степень развития зон торможения и обогащения. Чаще зона тормо- диев. В тех случаях, когда в зоне обогащения генеративных побегов имеется лишь главное соцветие (а оно нередко состоит только из одного парциального кистевидного тирса), зона торможения охватывает 10–15 узлов. В тех случаях, когда у генеративных побегов почти не выражена зона торможения (1–3 узла), в зоне обогащения развивается 8– 10 пар паракладиев. Таким образом, она охваты- жения охватывает 6–12 узлов, в зоне обогащения вает почти весь побег. развивается от 1 до 10 пар супротивных паракла-
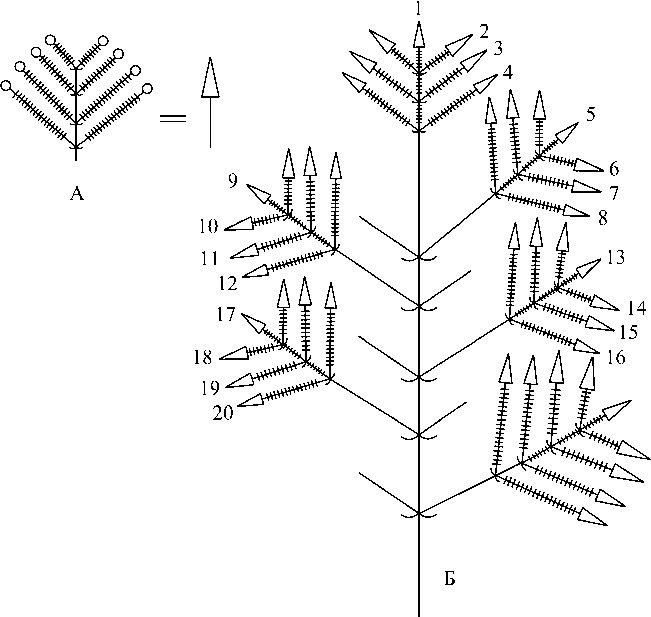
Рис. 1 . Схема синфлоресценции O. vulgare :
А – флоральная единица (кистевидный тирс), Б – верхняя часть синфлоресценции. 1–20 – порядковые номера флоральных единиц
Главная ось синфлоресценции и все её боковые ответвления (паракладии) завершаются группировкой цветоносных осей определённого строения – флоральной единицей, которая является основной структурной единицей соцветия. Флоральная единица у O. vulgare представлена открытым брактеозным тирсом, редуцированным до кистевидного соцветия (рис. 1). Все парциальные кистевидные тирсы (в главном соцветии и соцветиях паракладиев) у O. vulgare являются открытыми, поскольку они не заканчиваются верхушечным цветком, а их апекс засыхает в фазу плодоношения вместе с прикрывающими его зачатками нескольких пар новых брактей. Обычно их неточно называют колосками [Кузнецова, Пряхина, Яковлев, 1992], но в отличие от колосков, в парциальных кистевидных тирсах данного вида каждый пазушный цветок имеет короткую цветоножку, которая особенно хорошо выражена в фазе плодоношения.
Субъединицы тирса – цимоиды – у многих представителей данного семейства представляют собой вариант многоярусного дихазия (дихазий из монохазиев) [Troll, 1964]. У O. vulgare дихазий редуцирован до одного цветка на цветоножке и представляет собой одноцветковый цимоид. Таким образом, тирс в результате редукции трансформируется в открытую кисть (кистевидный тирс). Кистевидные флоральные единицы у этого вида составляют синфлоресценцию метёлковидного типа (рис. 1). Следовательно, тип синфлоресценции O. vulgare – фрондозно-брактеозная метёлка из кистевидных тирсов. За счёт уменьшения длины междоузлий как на главной оси, так и на паракладиях, происходит агрегация кистевидных тирсов, которые образуют соцветие щитковидной формы (рис. 1).
Цветение обоеполых и пестичных цветков O. vulgare. Обоеполые цветки O. vulgare строго протандричны. У таких цветков первой наступает мужская фаза цветения: через 1–2 ч. после раскрывания венчика, когда пыльники начинают растрескиваться. В период раскрывания венчиков и прохождения мужской фазы цветения в цветках происходят существенные изменения: в момент начала цветения тычинки начинают распрямляться, но пестик к этому времени ещё не развит. Только после раскрывания цветка постепенно происходит удлинение столбика пестика, который через 10–12 ч. доходит до половины трубки венчика, а через 24 ч. достигает пыльников, через 30–35 ч. он выходит из венчика и лопасти рыльца начинают отклоняться в разные стороны. Именно с этого момента начинается женская фаза цветения. Мужская фаза заканчивается усыханием пыльников. Продолжительность этой фазы около 35–38 ч.
Пыльники сохраняют пыльцу 6–8 ч. после раскрывания цветка. Опорожнение пыльников осуществляется в течение немногих часов, особенно у цветков, распустившихся утром, когда они посещаются насекомыми-опылителями. Фертильность пыльцы варьирует от 76.3 до 91.1%.
После раскрывания цветка рыльце бывает сначала игольчатым или сразу имеет вильчатую форму, направлено несколько косо кверху и не выступает из-под верхней губы. Подобные рыльца не готовы к опылению: при микроскопировании с ацетокармином видна не прорастающая на рыльце пыльца. Спустя некоторое время (3–4 ч. и более) вильчатое рыльце обычно несколько спускается вниз, и нередко его нижняя лопасть оказывается уже усеянной, особенно на самом кончике, пыльцевыми зёрнами. При микроскопировании в ацетокармине на рыльцах видны прорастающие пыльцевые зерна. Позднее, к концу дня или уже на следующий день, рыльца становятся якоревидными. Пыльца обычно локализована у них уже на обеих лопастях. Продолжительность женской фазы составляет 25–30 ч. Следовательно, обоеполый цветок O. vulgare функционирует 60–78 ч.
Продолжительность жизни пестичного цветка меньше, чем у обоеполых. Тычиночная стадия у них отсутствует в связи с редукцией тычинок до стаминодиев [Годин, Евдокимова, 2017]. Рыльце в своём развитии проходит те же несколько стадий, что и у обоеполых цветков: 1) рыльце игольчатое, скрытое под верхней губой венчика; 2) слегка выдвинутое, с вильчато разошедшимися лопастями (на данной стадии иногда уже выявляется пыльца при микроскопировании); 3) столбик длинный, рыльце выставляется далеко вперёд и немного вниз относительно верхней губы венчика, лопасти его якоревидно изогнуты. На данной стадии развития рыльца хорошо видна прорастающая пыльца. Полная жизнеспособность рыльца в пестичных цветках наступает через 5–7 ч. после его раскры- вания. Пестичные цветки увядают к концу второго дня цветения.
Полученные нами результаты согласуются с данными И.С. Шарыгиной [1959] о продолжительности цветения обоеполых цветков O. vulgare и Х. К. Джумаева и Р. М. Мухамеджановой [1990] об особенностях и длительности функционирования обоеполых и пестичных цветков у другого вида данного рода – O. tyttanthum . Имеющиеся в литературе сведения о более длительном функционировании обоеполых и пестичных цветков O. vulgare [Зимина, Конон, 1999] объяснить можно лишь неблагоприятными условиями для цветения, продолжающимися длительное время. Как продемонстрировала Е.И. Демьянова [1990], пестичная фаза цветков может пролонгироваться при отсутствии опыления из-за неблагоприятных условий окружающей среды для работы насекомых-опылителей.
Биологическим и визуальным показателем окончания цветения является засыхание пыльников или рылец [Левина, 1981]. Другими словами, цветение цветка заканчивается, когда пыльники и рыльца перестают функционировать.
Наблюдения за продолжительностью жизни раскрывшегося цветка показали, что она зависит от погодных условий, местообитания и эффективности опыления. У каждого вида растения период цветения отдельного цветка имеет свою определённость во времени. Однако эта определённость не всегда является стабильной и может изменяться [Демьянова, 1990].
Главными факторами, регулирующими продолжительность цветения раскрывшегося цветка, являются погодные условия – температура, влажность воздуха и освещённость. В тёплую сухую солнечную погоду цветки O. vulgare цвели меньшее число дней (обычно 2 дня), чем в прохладную и дождливую (цветение составляло 3–4 дня, но могло быть и более продолжительным при сохранении неблагоприятной погоды).
Погодные условия, в свою очередь, влияют на активность насекомых-опылителей, которая в значительной степени сказывается на продолжительности жизни раскрывшегося цветка. В холодную погоду или при длительных дождях активность насекомых наименьшая, поэтому неопылённые цветки долго сохраняют функционирующими тычинки и плодолистики. Удлинение периода цветения является прекрасным приспособлением к энтомофильному опылению при неблагоприятных условиях.
Продолжительность жизни раскрывшегося цветка также находится в зависимости от количества пыльцы в цветках, от числа и расположения цветков на особи или в соцветии, от достаточности опыления. Во время наших наблюдений для O.
vulgare было характерно обилие пыльцы в обоеполых цветках, большое число цветков на особях разного полового статуса. Совершенно очевидно, что все эти факторы действовали позитивно на растения, повышая эффективность опыления и стабилизируя продолжительность жизни раскрывшегося цветка.
Местообитание – тоже один из факторов разной продолжительности жизни раскрывшегося цветка при одних и тех же погодных условиях. Выяснилось, например, что цветки данного вида в тени (среди кус- тарников) цвели на 1–2 сут. дольше, чем цветки растений открытых местообитаний.
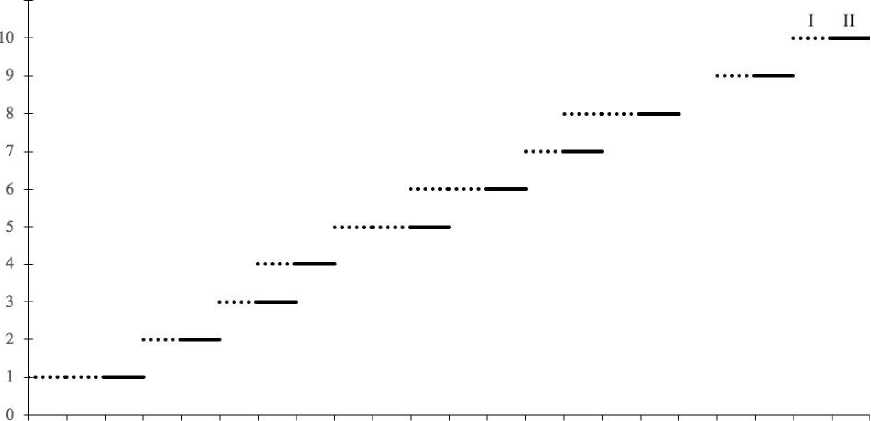
Цветение флоральной единицы . Цветение в кистевидных тирсах O. vulgare происходит в акропетальной последовательности. Последовательность цветения цветков в парциальных соцветиях представлена на рис. 2. Цветение начинается с раскрывания нижней пары цветков (рис. 2) и постепенно переходит на последующие, верхние пары.
s и
7.7 8.7 9.7 10.7 11.7 12.7 13.7 14.7 15.7 16.7 17.7 18.7 19.7 20.7 21.7 22.7 23.7 24.7 25.7 26.7 27.7 28.7 29.7
даты наблюдений
Рис. 2 . Последовательность и продолжительность цветения обоеполых цветков O. vulgare в пределах одного кистевидного тирса:
I – тычиночная фаза развития цветка, II – рыльцевая фаза
Начало цветения каждой последующей пары цветков наступает либо после отцветания предыдущей пары, либо немного раньше, так что в период цветения в колосовидных тирсах в определённые моменты может и не быть раскрывшихся цветков, но может находиться сразу две пары. Тем не менее, наиболее часто в парциальных соцветиях находится под одной паре раскрывшихся цветков. На рис. 2 очень четко видно, что довольно часто пестичная фаза в ниже расположенной паре цветков не совпадает с тычиночной фазой в выше расположенной паре цветков. Но иногда эти фазы совпадают. Следовательно, внутрицветковая протандрия не всегда обеспечивает ксеногамное опыление цветков во флоральных единицах O. vulgare. Возможно завязывание семян и в результате гей-тоногамного опыления за счет не всегда строго выраженной межцветковой протандрии. Продолжительность цветения цветков в парциальных соцветиях находится в прямой зависимости от числа пар цветков: если в соцветии имеется четыре пары цветков, продолжительность его цветения состав- ляет 8–9 дней, при наличии двенадцати пар – 27 дней.
Цветение синфлоресценции . Характер цветения синфлоресценций O. vulgare обусловлен спецификой их строения. Кистевидные тирсы в объединённом соцветии зацветают не одновременно: в начале наблюдается раскрывание цветков в кистевидных тирсах верхней части синфлоресценции, позднее – в нижней части. Зацветание флоральных единиц (частных соцветий) в пределах главного побега и паракладиев, а также самих паракладиев в составе синфлоресценции происходит в базипетальном порядке.
Цветение кистевидных тирсов начинается с верхушечной части главного соцветия и постепенно переходит на его нижние боковые ветви. Разница во времени цветения флоральных единиц верхних и нижних частей синфлоресценции составляет 3–5 дней. Такая же закономерность наблюдается и в цветении флоральных единиц в пределах главного соцветия и паракладиев первого порядка: цветение конечных флоральных единиц начинается на
2–3 дня раньше, чем цветение самых нижних фло-ральных единиц (рис. 3). Как следует из приведенных данных, одновременно в синфлоресценции O. vulgare может цвести большое число цветков, находящихся в тычиночных или пестичных фазах развития, что указывает на возможность завязыва- семян в результате гейтоногамного опыления.
ния
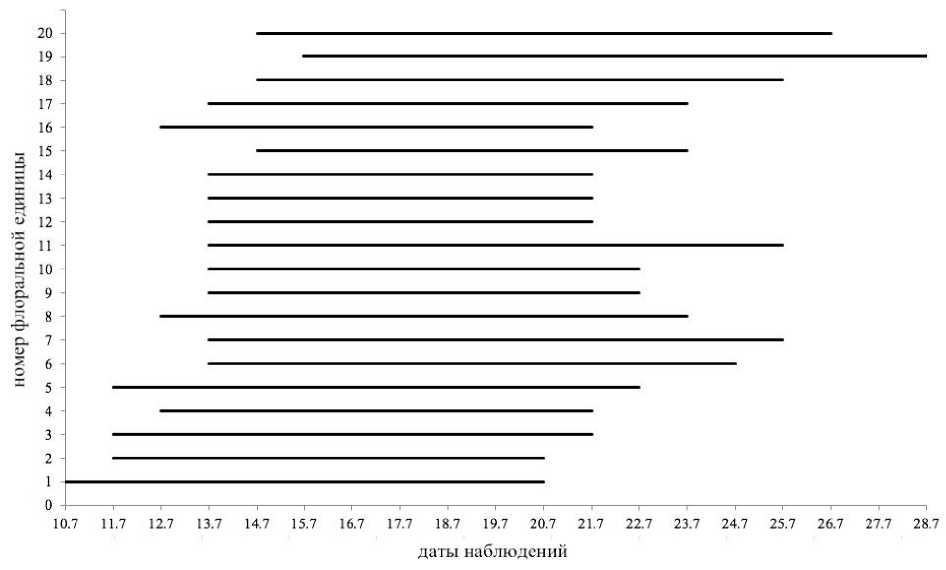
Рис. 3 . Последовательность и продолжительность цветения кистевидных тирсов в верхней части синфлоресценции O. vulgare (порядковые номера флоральных единиц как на рис. 1)
Как следует из приведенных данных, характер раскрывания и отцветания флоральных единиц, соцветий паракладиев и синфлоресценции в целом зависит от числа в них цветков. В соцветиях с большим числом цветков этот процесс идет более равномерно и значительно растянут. В соцветиях же с меньшим числом этот процесс протекает более интенсивно и сравнительно быстро заканчивается. Однако наблюдая различие в цветении син-флоресценций различных особей, не мешает отметить, что максимум открытых цветков в синфло-ресценции приходится на 6–10-й день после начала цветения. Это совпадает со временем цветения 2–3-й пары цветков в кистевидных тирсах верхней части синфлоресценции и раскрыванием 1–2-й пары цветков в кистевидных тирсах нижних частей синфлоресценции. Это состояние и следует считать за полное цветение особи. В дальнейшем число отцветших цветков превышает число раскрывающихся цветков. По мере отцветания нижних пар цветков во флоральных единицах и раскрывания последующих верхних пар кистевидные тирсы заметно удлиняются.
В результате проведенных исследований нами не выявлены структурные отличия составных час-
Полученные данные свидетельствуют также о том, что продолжительность цветения синфлорес-ценции в целом зависит от числа пар цветков в парциальных соцветиях: при увеличении числа пар цветков в кистевидных тирсах с четырёх-пяти до десяти-одиннадцати особь цветёт в течение 31–32 вместо 17–18 дней.
тей и синфлоресценций в целом у обоеполых и женских особей O. vulgare . Тем не менее, размерные и количественные различия отмечаются. Так, флоральные единицы и синфлоресценции обоеполых особей, как правило, образуют большее число цветков, чем те же структуры у особей с пестичными цветками. Аналогичные данные были получены Е.И. Демьяновой [2012] при изучении особенностей проявления полового диморфизма у O. vulgare в строении и размерах генеративных побегов.
Выводы
-
1. Синфлоресценция O. vulgare полителиче-ская, флоральная единица – открытый колосовидный тирс, многочисленные разветвленные пара-кладии несут такие же флоральные единицы. Тирсы брактеозные, их боковые оси редуцированы до одного цветка.
-
2. Обоеполые цветки O. vulgare строго протан-дричны, функционируют в среднем 2–3 дня. Пестичные цветки цветут 1.5–2 дня.
-
3. Раскрывание цветков во флоральных единицах происходит в акропетальной последовательно-
- сти. Зацветание флоральных единиц в пределах главного побега и паракладиев, а также самих па-ракладиев в составе синфлоресценции происходит базипетально.
-
4. Несмотря на хорошо выраженную внутри-цветковую протандрию, наличие не строгой межцветковой протандрии приводит к возможности гейтоногамного завязывания семян у обоеполых особей O. vulgare.
2011. № 2 (8). С. 49–53.
Список литературы Особенности биологии цветения Origanum vulgare L. (Lamiaceae) в Московской области
- Байкова Е.В. Род шалфей: морфология, эволюция, перспективы интродукции. Новосибирск: Наука, 2006. 248 с.
- Буданцев А.Л. Морфологические преобразования соцветий у видов рода Nepeta (Lamiaceae) // Ботанический журнал. 1992. Т. 77, № 7. С. 79-84.
- Годин В.Н. Половой полиморфизм видов растений подкласса Lamiidae в Сибири. Обзор литературы // Растительный мир Азиатской России. 2011. № 2 (8). С. 49-53.
- Годин В.Н., Демьянова Е.И. О распространении гинодиэции у цветковых растений // Ботанический журнал. 2013. Т. 98, № 12. C. 1465-1487.
- Годин В.Н., Евдокимова Т.А. Морфология обоеполых и пестичных цветков Origanum vulgare // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2017. № 1 (17). С. 3-13.
- Джумаев Х.К., Мухамеджанова Р.М. Биология цветения и плодоношения Salvia sclarea L. и Origanum tyttanthum Gontsch. (Узбекская ССР) // Растительные ресурсы. 1990. Вып. 4. С. 519-524.
- Демьянова Е.И. Половой полиморфизм цветковых растений: автореф. дис.. д-ра биол. наук. М., 1990. 35 с.
- Демьянова Е.И. О половом полиморфизме душицы обыкновенной (предварительное сообщение) // Вестник Пермского университета. Биология. 2012. Вып. 3. С. 8-10.
- Зимина Л.Б. Селекция душицы обыкновенной в Московской области: автореф. дис.. канд. биол. наук. М., 2002. 23 с.
- Зимина Л.Б., Конон Н.Т. Биология цветения Origanum vulgare, выращиваемого в Москве // Растительные ресурсы. 1999. Вып. 4. С. 25-31.
- Кузнецова Т.В. Морфология соцветий: современное состояние // Итоги науки и техники. Сер. Ботаника. М., 1991. С. 51-174.
- Кузнецова Т.В., Пряхина Н.И., Яковлев Г.П. Соцветия: морфологическая классификация. СПб., 1992. 127 с.
- Левина Р.Е. Репродуктивная биология семенных растений. М.: Наука, 1981. 96 с.
- Нухимовский Е.Л., Черкасов О.А. Морфология Origanum vulgare L. в естественных местообитаниях и при выращивании в Московской области // Растительные ресурсы. 1987. Т. 23, вып. 3. С. 345-356.
- Пономарев А.Н. Изучение цветения и опыления растений // Полевая геоботаника. М.; Л.: Изд-во АН СССР, 1960. Т. 2. С. 9-19
- Шарыгина И.С. Некоторые вопросы биологии и биохимии Origanum vulgare L. и Hyssopus officinalis L. // Ботанический журнал. 1959. Т. 44, № 8. С. 1124-1128.
- Knuth P. Handbuch der Blutenbiologie. Leipzig: Verlag von Wilchelm Engelmann, 1899. Bd. II, T. 2. 705 S.
- Robinsohn I. Die Färbungsreaktion der Narbe, Stigmatochromie, als morpho-biologische Blütenuntersuchungsmethode // Sitzungsberichte der Akademie der Wissenschaften mathematischnaturwissenschaftliche Klasse. 1924. Vol. 133. P. 181-211
- Sell Y. Physiological and phylogenetic significance of the direction of flowering in inflorescence complexes // Flora. 1980. Vol. 169, № 4. P. 282-294.
- Troll W. Die Infloreszenzen. Jena: Fisher Verlag, 1964. Bd. 1. 615 S.
