Особенности дыхания и фотосинтеза листьев у разновозрастных деревьев тополя, березы повислой и сосны обыкновенной

Автор: Евлаков П.М., Амелин А.В., Жужукин К.В., Рыжкова В.С., Евтушенко Н.А.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агрономия
Статья в выпуске: 3, 2026 года.
Бесплатный доступ
Цель исследований - выделение перспективных пород и сортов деревьев для последующего использования в реализации лесоклиматических проектов. Исследования проведены с использованием системного подхода и современных приборов учета показателей активности дыхания и фотосинтеза. Объекты исследования - генотипы древесных пород разного возраста: у тополя (Populus L.) - 4- и 10-летние, у березы повислой (Betula L.) - 4- и 46-летние, у сосны обыкновенной (Pinus L.) - 4- и 50-летние, которые были выращены на экспериментальном калибровочном полигоне учебно-опытного лесхоза ВГЛТУ. Для измерения скорости поглощения и выделения углекислого газа листом in vivo использовали газовый анализатор CI-340 (SID Bio-Science, США). Замеры интенсивности фотосинтеза проводили в режиме онлайн на интактных листьях деревьев в утреннее время с 08:00 до 11:00 ч, когда насыщенность клеток водой высокая, а интенсивность солнечного света и температура воздуха близки к оптимальным. Учет активности темнового дыхания осуществляли с 23:00 и до 01:00 ч. Наиболее активным темновым дыханием отличались листья быстрорастущего тополя, который превосходил по значению данного показателя березу повислую в среднем на 21,0 %, а сосну обыкновенную на 26,3 %. Видовые различия по активности фотосинтеза были идентичны. У 10-летнего тополя (сорт ʻЭ.с.-38ʼ) значения интенсивности фотосинтеза и темнового дыхания были на 9,6 и 15,8 %, у 46-летней березы повислой (сорт ʻУг.-1ʼ) на 32,6 и 13,3 %, а у 50-летней сосны обыкновенной на 10,9 и 14,0 % меньше по сравнению с деревьями 4-летнего возраста. В создании лесоклиматических проектов по улучшению экологии среды обитания и производства наиболее перспективным из изученных пород деревьев является быстрорастущий тополь.
Дыхание, фотосинтез, устьичная проводимость, видовые и генотипические особенности, тополь, береза повислая, сосна обыкновенная, экология среды обитания
Короткий адрес: https://sciup.org/140314954
IDR: 140314954 | УДК: 581.132.2 | DOI: 10.36718/1819-4036-2026-3-27-37
Features of respiration and photosynthesis in leaves of different-aged trees of poplar, silver birch, and Scots pine
The objective of research is to identify promising tree species and cultivars for subsequent use in forest climate projects. The study was conducted using a systems approach and modern instruments for measuring respiration and photosynthesis activity. The objects of the study were tree genotypes of different ages: 4- and 10-year-old poplar (Populus L.), 4- and 46-year-old silver birch (Betula L.), and 4- and 50-year-old Scots pine (Pinus L.), which were grown on the experimental calibration site of the VSTU educational and experimental forestry enterprise. A CI-340 gas analyzer (SID Bio-Science, USA) was used to measure the rate of carbon dioxide absorption and emission by leaves in vivo. Online measurements of photosynthesis rates were taken on intact tree leaves in the morning from 8:00 AM to 11:00 AM, when cell water saturation is high and sunlight intensity and air temperature are close to optimal. Dark respiration activity was recorded from 11:00 PM to 1:00 AM. The leaves of fast-growing poplar exhibited the most active dark respiration, exceeding those of silver birch by an average of 21.0% and Scots pine by 26.3%. Species-specific differences in photosynthetic activity were identical. In 10-year-old poplar (variety 'E.s.-38'), the photosynthesis intensity and dark respiration rates were 9.6 and 15.8 % lower, respectively; in 46-year-old silver birch (variety Ug.-1), they were 32.6 and 13.3 % lower, respectively; and in 50-year-old Scots pine, they were 10.9 and 14.0% lower, respectively, compared to 4-year-old trees. Fast-growing poplar is the most promising of the studied tree species for the development of forest climate projects to improve the ecology of habitats and production.
Текст научной статьи Особенности дыхания и фотосинтеза листьев у разновозрастных деревьев тополя, березы повислой и сосны обыкновенной
Funding: the study was conducted within the framework of the State Assignment of the Ministry of Science and Higher Education of the Russian Federation No. 1025032100180-3-1.6.11 Development of a Scientifically Substantiated Technology for Accelerated Cultivation of Standardized Planting Material of Deciduous and Coniferous Tree Species by In vitro Clonal Micropropagation Methods, as well as In vivo Seed Propagation for the Creation of Carbon-Storing Plantations in the Context of Global Climate and Anthropogenic Change in the Forest-Steppe Zone of European Russia (FZUR-2026-0005).
Введение. Изучению взаимосвязи фотосинтеза и дыхания у древесных растений посвящено множество отечественных [1–5] и зарубежных [6–10] исследований, показавших, что данные процессы являются ключевыми в обеспечении глобального углеродного баланса и образования биомассы на планете. Они тесно сопряжены между собой, хотя имеют разное физиологическое назначение. Фотосинтез направлен на преобразование электромагнитной энергии квантов света в энергию химических связей в виде АТФ и НАДФН, которая расходуется в частности на ассиимляцию СО2 из воздуха и образование органического вещества с выделе- нием кислорода [11]. Тогда как в процессе дыхания, наоборот, происходит окисление образующихся углеводов молекулярным кислородом с высвобождением в атмосферу большого количества молекул СО2 и свободной энергии [1]. Ежегодно около половины CO2, ассимилируемого растениями через фотосинтез, возвращается в атмосферу посредством дыхания растений [7], а в стрессовых условиях может достигать 80 % [5, 8].
При этом на свету большую часть работы в клетке выполняет фотосинтез, а на долю дыхания остается существенно меньше, что неизбежно ведет к ее замедлению, предполагаемо из-за конкуренции [12]. В результате дыхание на свету оказывается примерно в 2–3 раза выше, чем дыхание в темноте [13].
Но, какими бы не были биохимические изменения темнового дыхания на свету, вновь образованные при фотосинтезе углеводы сразу включаются в дыхательный метаболизм фотосинтезирующих тканей на свету, не ожидая темнового периода суток [13]. Поэтому даже небольшое повышение интенсивности данных процессов может оказать значительное влияние на годовой прирост биомассы и глобальный углеродный цикл природы [14–17].
Учитывая вышеизложенное, в 2024 г. нами были организованы и проведены специальные экспериментальные исследования по указанной тематике.
Цель исследования – установить характер взаимодействия дыхания и фотосинтеза у раз- личных по возрасту древесных пород и выделить из них перспективные виды и генотипы для последующего использования в реализации лесоклиматических проектов.
Объекты и методы . Исследования проведены в Воронежском ГЛТУ с использованием современных приборов учета показателей активности дыхания и фотосинтеза. Объектами изучения являлись селекционно ценные генотипы быстрорастущих древесных пород тополя ( Populus L., межсекционный гибрид – сорт ʻ Э.с.-38 ʼ ) и березы повислой ( Betula L., сорт ʻ Углянческая 1 ʼ (Уг.-1), а также сосны обыкновенной ( Pinus L.).
Изучаемые деревья произрастали на опытном участке площадью 0,3 га на территории учебно-опытного лесхоза ВГЛТУ по заранее разработанной схеме (рис. 1).
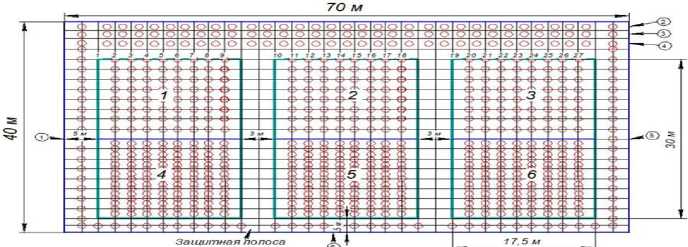
Рис. 1. Схема экспериментального карбонового полигона на территории учебно-опытного лесхоза ВГЛТУ (1–27 – номер ряда; 1–6 – номер секции; 0 – посадочные места;
(1) и (6) – ряды защитной полосы)
Layout of the experimental carbon polygon on the territory of the Training and Experimental Forest Enterprise of Voronezh State University of Forestry and Technologies (Voronezh State University of Forestry and Technologies) (designations: 1–27 – row number; 1–6 – section number; 0 -– planting spots;
(1) and (6) – protective belt rows).
Для измерения скорости поглощения и выделения углекислого газа листом использовали портативный инфракрасный газовый анализатор CI-340 (CID Bioscience, США), который оборудован специальной листовой камерой, сконструированной по типу открытой системы: окошко камеры имеет размер 25 × 25 мм, площадь 6,25 см2 (рис. 2).
Лист, помещенный в камеру, находился в таких же условиях освещения, в каких он был вне камеры, что обеспечивается прозрачностью окошка листовой камеры. Температура листа контролировалась путем изменения температурного режима воздушной среды внутри камеры.
Замеры интенсивности фотосинтеза (ИФ или Pg) и устьичной проводимости (УП) проводили в режиме онлайн на интактных листьях деревьев 12, 13 августа (в конце фазы интенсивного роста древесных растений) в утреннее время с 8:00 до 11:00 ч, когда насыщенность клеток водой высокая, а интенсивность солнечного света и температура воздуха близки к оптимальному. Учет активности темнового дыхания (Rd) осуществляли с 23:00 и до 01:00 ч. Повторность анализа для каждой точки учета была 3–5-кратной. Расчет нетто фотосинтеза осуществляли при помощи встроенного в прибор программного обеспечения по формуле [18].

Рис. 2. Измерение параметров газообмена и транспирации сосны обыкновенной с помощью инфракрасного газового анализатора CI-340 Measurement of gas exchange and transpiration parameters in Scots pine using the CI-340 portable infrared gas analyzer
Экономический коэффициент фотосинтеза листьев (Ер) рассчитывали отношением суммы значений фотосинтеза и темнового дыхания к значению темнового дыхания (Ер = Рg + Rd/Rd).
Метеоусловия вегетации опытных деревьев в весенне-летний период в целом характеризовались умеренным увлажнением, обеспеченностью теплом и светом, т. е. были относительно благоприятными для роста и развития. Динамика погодных условий, формировавшихся на исследуемой территории, показана в таблице 1.
Статистическую обработку экспериментальных данных проводили с использованием современных компьютерных программ.
Таблица 1
Характеристика метеоусловий во время проведения опытов в 2024 г. по данным Метеостанции P0016: Воронежская область, Левобережное лесничество, кордон Кожевенный Characterization of weather conditions during the experiments in 2024 according to Meteorological
Station P0016: Voronezh Region, Levoberezhnoye Forestry, Kordón Kozhevennyy
|
Месяц |
Декада |
Температура воздуха, °С |
Суммар. ФАР, Вт/м2 |
Средняя влажность воздуха, % |
Сумма осадков, мм |
||
|
60 см |
70 см |
80 см |
|||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
|
Апрель |
I |
12,3 |
1195 |
62 |
0,00 |
9,2 |
8,3 |
|
II |
14,0 |
1279 |
55 |
6,0 |
7,6 |
6,8 |
|
|
III |
14,0 |
1280 |
66 |
6,4 |
6,7 |
6,0 |
|
|
Среднее |
13,4 |
1251 |
61 |
12,4 |
7,8 |
7,0 |
|
|
Май |
I |
9,9 |
1625 |
62 |
20,2 |
5,6 |
5,8 |
|
II |
10,4 |
1420 |
68 |
6,4 |
5,0 |
4,0 |
|
|
III |
18,3 |
2186 |
53 |
1,6 |
4,3 |
3,2 |
|
|
Среднее |
12,9 |
1744 |
61 |
28,2 |
5,0 |
4,3 |
|
|
Июнь |
I |
21,6 |
1983 |
65 |
3,6 |
4,0 |
3,0 |
|
II |
21,5 |
1552 |
78 |
69,4 |
4,0 |
3,0 |
|
|
III |
19,5 |
1979 |
72 |
7,2 |
5,2 |
3,0 |
|
|
Среднее |
20,9 |
1838 |
72 |
80,2 |
4,4 |
3,0 |
|
|
Июль |
I |
24,7 |
2090 |
65 |
14,8 |
4,5 |
3,0 |
|
II |
24,9 |
2096 |
65 |
13,2 |
4,0 |
3,0 |
|
|
III |
20,0 |
1583 |
68 |
10,2 |
4,0 |
3,0 |
|
|
Среднее |
23,2 |
1983 |
66 |
37,6 |
4,3 |
3,0 |
|
Окончание табл. 1
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
|
Август |
I |
19,3 |
1629 |
75 |
23,6 |
4,0 |
3,0 |
|
II |
19,4 |
1711 |
65 |
0,4 |
4,0 |
3,0 |
|
|
III |
22,7 |
1425 |
55 |
0,0 |
4,0 |
3,0 |
|
|
Среднее |
20,5 |
1588 |
65 |
24,0 |
4,0 |
3,0 |
|
|
Дата учета |
12.08 |
16,4 |
1707 |
70 |
0,20 |
4,0 |
3,0 |
|
13.08 |
16,2 |
1395 |
73 |
0,00 |
4,0 |
3,0 |
Максимальный приход ФАР начинал проявляться в конце мая (III декада) и продолжался до начала III декады июля. Наиболее жарким был июль, а влажным – июнь. Во время проведения учетов (12 и 13 августа) отмечались невысокая температура воздуха (16,4 и 16,2 °С) и приход ФАР при относительной влажности воздуха 70–73 % (см. табл. 1).
Результаты и их обсуждение. В условиях естественной темноты произрастания растений (с 11:00 и до 01:00 ч), интенсивность дыхания листьев у молодых (4-летних) опытных деревьев варьировала от 1,40 до 1,91 мкмоль СО 2 /м2/с. Наиболее активным темновым дыханием отличались листья быстрорастущего тополя, который превосходил по значению данного показателя березу повислую в среднем на 21,0 %, а сосну обыкновенную – на 26,3 %.
Видовые различия по активности темнового дыхания во многом были схожи с экспериментальными данными по фотосинтезу, что подтверждает тесную взаимосвязь этих двух важнейших физиологических процессов в растениях [4, 5, 9].
У изученных нами 4-летних деревьев максимальная интенсивность фотосинтеза листьев во время активного роста варьировала от 18,9 до 33,3 мкмоль CO 2 /м2/с. Наибольшая ее величина так же отмечалась у гибрида тополя, а наименьшая – у сосны обыкновенной. Береза повислая (сорт ʻ Уг. 1 ʼ ) занимала промежуточное положение. Максимальные значения ассимиляции СО 2 у листьев сорта тополя были выше, чем у сорта березы повислой, в среднем на 13,5 %, а по сравнению с сосной обыкновенной – на 43,2 %.
При этом интенсивность темнового дыхания листьев у изученных пород деревьев была ниже активности их фотосинтеза в среднем в 16,7 раза. Отношение интенсивности темнового дыхания к интенсивности фотосинтеза (Rd/Pg) на плато световой кривой составляло в среднем 0,06, или 6,1 %, в т. ч. у тополя – 5,7 %; у березы повислой – 5,2 %, у сосны обыкновенной – 7,4 %. Экономический коэффициент фотосинтеза (Ер) варьировал у 4-летних деревьев от 14,3 до 20,2 и был наиболее высоким у быстрорастущих березы повислой и тополя, которые превосходили сосну обыкновенную в среднем в 1,4 раза (табл. 2).
Таблица 2
Интенсивность темнового дыхания (Rd) и фотосинтеза (Pg) и эффективность их взаимодействия у 4-летних пород деревьев (по данным 2024 г.)
Intensity of Dark Respiration (Rd) and Photosynthesis (Pg), and the Efficiency of Their Interaction in 4-Year-Old Tree Species (According to 2024 Data)
|
Растение |
(Rd), мкмоль СО 2 /м2/с |
Pg (ИФ), мкмоль СО 2 /м2/с |
Pg /Rd |
Rd/Pg |
Ер, (Рg + Rd) / Rd |
|
Тополь, сорт Э.с. 38 |
1,91 ± 0,24 |
33,3 ± 0,41 |
17,4 |
0,057 |
18,4 |
|
Береза повислая, сорт У.г.-1 |
1,50 ± 0,21 |
28,8 ± 0,71 |
19,2 |
0,052 |
20,2 |
|
Сосна обыкновенная |
1,42 ± 0.16 |
18,9 ± 0,54 |
13,3 |
0,075 |
14,3 |
В известной степени это согласуется и с литературными данными, свидетельствующими, что максимальные значения фотосинтеза в сезоне приурочены к периодам интенсивного апикального и радиального роста дерева [19]. Считается, что 60–70 % накопленных при фотосинтезе углеводов расходуется на образование углеродных скелетов, а 30–40 % идет на образование АТФ и восстановителей [12]. То есть на свету большую часть работы в клетке выполняет фотосинтез, а на долю дыхания остается существенно меньше, что неизбежно ведет к ее замедлению. Биологическое значение такого переключения заключается в том, чтобы в клетке не тратился на свету запас ранее накопленных ассимилятов. Благодаря этому устанавливается определенный баланс между расходом дыхательных субстратов на синтез АТФ и восстановительных эквивалентов и потребностью в углеродных скелетах как для построения и поддержания клеточных структур, так и для образования запасных веществ [12].
При достижении определенного возраста интенсивность фотосинтеза листьев деревьев на- чинала заметно снижаться: у 10-летнего тополя (сорт ʻЭ.с.-38ʼ) в среднем на 9,6 %, у 46-летней березы повислой (сорт ʻУг.-1ʼ) – на 32,6, а у 50летней сосны обыкновенной на 10,9 % по сравнению с деревьями 4-летнего возраста. При этом видовые различия остались прежними.
С увеличением возраста дерева снижалась активность и темнового дыхания (в среднем на 14,4 %), в т. ч. у 10-летнего тополя в среднем на 15,8 %, у 46-летней березы повислой – на 13,3, а у 50-летней сосны обыкновенной – на 14,0 % по отношению к 4-летним деревьям. Отношение активности дыхания к интенсивности фотосинтеза было относительно стабильным и составляло в среднем 0,063, или 6,3 %. Однако выявленные возрастные изменения позволили увеличить экономический коэффициент фотосинтеза листьев (Ер) у тополя и сосны обыкновенной в среднем на 6,9 и 5,3 %, а у березы повислой снизить его в среднем на 21,3 %, прежде всего по причине сильного падения активности фотосинтеза (в среднем на 32,6 %) и повышения транспирационной активности листьев более чем на 40 % (табл. 3).
Таблица 3
Интенсивность темнового дыхания (Rd) и фотосинтеза (Pg) и эффективность их взаимодействия у пород деревьев разного возраста (по данным 2024 г.)
Intensity of Dark Respiration (Rd) and Photosynthesis (Pg), and the Efficiency of Their Interaction in Different-Aged Tree Species (According to 2024 Data)
|
Растение |
Возраст, лет |
Rd, мкмоль/м2/с |
Pg (ИФ), мкмоль СО 2 /м2/с |
Pg /Rd |
Rd/Pg |
Ер, (Рg+ Rd) / Rd |
|
Тополь, сорт Э.с. 38 |
10 |
1,6 ± 0,31 |
30,1 ± 0,57 |
18,8 |
0,053 |
19,8 |
|
Береза повислая, сорт Уг.-1 |
46 |
1,3 ± 0,5 |
19,4 ± 0,26 |
14,9 |
0,067 |
15,9 |
|
Сосна обыкновенная |
50 |
1,2 ± 0,18 |
16,9 ± 0,13 |
14,0 |
0,071 |
15,1 |
Схожие экспериментальные данные приводят по многим параметрам и другие ученые, проводившие исследования с использованием аналогичных приборов. К примеру, в условиях Карелии интенсивность темнового дыхания составляла у молодых листьев Карельской березы 2,22 мкмоль СО 2 /м2/с, а у сформировавшихся – 1,76 мкмоль СО 2 /м2/с. У березы повислой значения показателя были чуть меньше: 2,14 и 1,47 мкмоль СО 2 /м2/с соответственно [19].
В опытах В. Б. Придача с соавт. (2022, 2023), изучавших газообмен древесных растений на экс- периментальном участке пригородной территории г. Грозного, интенсивность темнового дыхания в начале весны (май) и осени (сентябрь) соответственно составляла: у дуба черешчатого – 1,8 и 2,0; у клена полевого 1,4 и 0,2 мкмоль СО2/м2/с [20, 21].
Снижение активности фотосинтеза листьев с увеличением возраста дерева обычно связывают с ослаблением ростовых процессов у нефотосинтезирующих органов [1, 12, 19]. При этом соотношение активности дыхания и фотосинтеза (R/Pg) остается на определенном уровне ввиду сохра- нения углеродного баланса в ходе роста и развития растений и в большинстве случаев находится в пределах 0,35–0,80. Однако это соотношение не постоянно [5, 6, 9, 14]. Наименьшие значения регистрируются весной и ранним летом, во время основного периода роста, а наибольшие – в осенне-зимний период и свидетельствуют о больших расходах на рост [6].
Низкие значения отношения дыхания к фотосинтезу в наших экспериментах в основном связаны с тем, что в расчетах использовались данные только по темновому дыханию, которое в 2–3 раза ниже, чем дыхание на свету [3]. Если произвести расчеты с учетом данного факта (R = Rg · 3), то мы получим данные, которые будут близки к имеющимся в литературе сведениям: у опытных деревьев интенсивность фотосинтеза станет выше интенсивности дыхания в 4,7–6,3 раза, а отношение R/Pg будет находиться в пределах 0,16–0,21, или составит 16–21 % (табл. 4).
Таблица 4
Интенсивность дыхания в темноте и на свету и ее связь с активностью фотосинтеза у изученных пород деревьев разного возраста (по данным 2024 г.)
Intensity of Respiration in Darkness and in Light, and Its Relationship with Photosynthetic
Activity in Different-Aged Studied Tree Species (According to 2024 Data)
|
Растение |
Возраст, лет |
Темновое дыхание (Rd), мкмоль/м2/с |
Дыхание на свету (R = Rd · 3), мкмоль/м2/с |
Pg (ИФ), мкмоль, СО 2 /м2/с |
Pg /R |
R/Pg |
Ер,(Рg + R) / R |
|
Тополь, сорт Э.с. 38 |
10 |
1,61 ± 0,31 |
4,8 ± 0,31 |
30,1 ± 0,57 |
6,3 |
0,16 |
6,3 |
|
Береза повислая, сорт У.г.-1 |
46 |
1,30 ± 0,15 |
3,9 ± 0,15 |
19,4 ± 0,26 |
5,0 |
0,20 |
5,0 |
|
Сосна обыкновенная |
50 |
1,20 ± 0,18 |
3,6 ± 0,18 |
16,9 ± 0,13 |
4,7 |
0,21 |
4,7 |
По-видимому, нельзя исключить наличие и других причин существующих различий с литературными данными по R/Pg, связанных, к примеру, с особенностями природно-климатических условий места произрастания растений и характером температурного режима во время проведения учетов [10, 22–24], а также с временем учета и местом расположения в кроне анализируемых листьев.
На взаимоотношения дыхания и фотосинтеза определенное влияние может оказывать и устьичная проводимость листьев. Показано, что у деревьев разного класса роста фотосинтез при высокой инсоляции (освещенность выше 100 Вт/м2) в основном определяется величиной и динамикой изменения мезофильного сопротивления (rm), которое более чем в 5 раз превосходит устьичное (rs.). Но при восходе и заходе солнца основную роль играет устьичное сопротивление. Обнаружена положительная корреляция устьичного и мезофильного сопротив- лений на протяжении длительного летнего периода. Для величин фотосинтеза, превосходящих 12 мг СО2/(г · ч), связь мезофильного и устьичного сопротивления становится практически линейной – г = 0,97 [12, 19].
В наших опытах значения устьичной проводимости листьев у изученных 4-летних деревьев изменялись в диапазоне от (78 ± 6,4) до (146 ± 5,9) ммоль/(м2 · с), что свидетельствует о высокой видовой обусловленности показателя. Наибольшая его величина зафиксирована у тополя ((146 ± 5,9) ммоль/(м2 · с)) и была в 1,9 раза выше, чем у березы повислой, и в 1,3 раза выше, чем у сосны обыкновенной (рис. 3).
С увеличением возраста устьичная проводимость листьев у опытных деревьев повышалась с 113,3 до 155,5 ммоль/(м2 · с), или в среднем на 27,1 %, в т. ч. у 10-летнего тополя на 29,6 %, у 46-летней березы повислой – на 43,5, а у сосны обыкновенной – на 4,1 % (рис. 4).
146 ± 5,9

Сосна Береза повислая Тополь сорт Э.с.
обыкновенная сорт У.г. - 1 38
Рис. 3. Устьичная проводимость листьев (УП) у пород деревьев 4-летнего возраста разного возраста (по данным учетов 2004 г.) Leaf stomatal conductance (gs) in 4-year-old tree species of different ages (according to 2004 census data)
[ЗНАЧЕНИЕ] ± 4

207,5 ± 6,5

Сосна Береза повислая Тополь сорт Э.с.обыкновенная сорт У.г. - 1 38
Рис. 4. Устьичная проводимость листьев (УП) у пород деревьев разного возраста (тополь – 10 лет, береза повислая – 46 лет, сосна обыкновенная – 50 лет) (по данным учетов 2004 г.)
Leaf stomatal conductance (gs) in tree species of different ages
(poplar – 10 years, silver birch – 46 years, Scots pine – 50 years) (according to 2004 census data)
Таблица 5
Устьичная проводимость (УП), интенсивность фотосинтеза (ИФ), интенсивность транспирации (ИТ) и темновое дыхание (Rd) у пород деревьев разного возраста (по данным 2024 г.)
Stomatal conductance (gs), photosynthetic rate (A), transpiration rate (E), and dark respiration (Rd) in tree species of different ages (according to 2024 data
|
Растение |
Возраст, лет |
УП, ммоль CO 2 /м2/с |
ИФ max, мкмоль СО 2 /м2/с |
ИТ, ммоль Н 2 О/м2/с |
Rd, мкмоль CO 2 /м2/с |
|
Тополь, сорт Э.с. 38 |
4 |
146 ± 5,9 |
33,3 ± 0,41 |
3,3 ± 0,12 |
1,91 ± 0,24 |
|
10 |
207,5 ± 6,5 |
30,1 ± 0,57 |
4,0 ± 0,08 |
1,61 ± 0,31 |
|
|
Береза повислая, |
4 |
78 ± 6,4 |
28,8 ± 0,71 |
1,9 ± 0,15 |
1,50 ± 0,21 |
|
сорт У.г.-1 |
46 |
138 ± 5,0 |
19,4 ± 0,26 |
3,3 ± 0,11 |
1,30 ± 0,15 |
|
Сосна обыкновенная |
4 |
116 ± 2,2 |
18,9 ± 0,54 |
2,6 ± 0,10 |
1,42 ± 0,16 |
|
50 |
121 ± 4,8 |
16,9 ± 0,13 |
2,3 ± 0,07 |
1,20 ± 0,18 |
Возрастное увеличение устьичной проводимости листьев у тополя и березы повислой было обусловлено прежде всего повышением их транспирационной активности, поскольку интенсивность ассимиляции СО 2 снижалась и негативно сказывалась на активности темнового дыхания (табл. 5).
Заключение. Таким образом, на основе комплексного анализа характера взаимодействия дыхания, фотосинтеза и устьичной проводимости из изученных пород деревьев наиболее перспективным в реализации лесоклиматических проектов является прежде всего быстрорастущий тополь, который на протяжении длительного периода вегетации характеризуется более высокими интенсивностью фотосинтеза и дыхания, их соотношением и экономическим коэффициентом эффективности фотосинтеза.
Кроме того, формируя мощную крону, тополь проявляет хорошие шумозащитные [6] и антимикробные свойства.
Во многом схожими параметрами обладает и береза повислая, тогда как сосна обыкновенная по многим указанным параметрам значимо уступает им. В силу этого она менее перспективна для использования в нормализации загазованности среды обитания и производства и, как следствие, в сдерживании глобального потепления климата.
