Особенности формирования биоморф у некоторых травянистых многолетников

Автор: Османова Г.О., Шивцова И.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 1-1 т.13, 2011 года.
Бесплатный доступ
В результате сравнительного морфологического анализа растений модельных видов, произрастающих в неодинаковых экологических условиях, описаны разные типы биоморф. Отмечено, что тип биоморф у Fragaria vesca L. меняется не только в ходе онтогенеза, но и в течение вегетационного периода, а у Plantago lanceolata L. - определяется жизненной формой и особенностями экотопа.
Жизненная форма, биоморфы: моноцентрическая, неявнополицентрическая, явнополицентрическая, ацентрическая, переходная
Короткий адрес: https://sciup.org/148199610
IDR: 148199610 | УДК: 581.4+582.96+582.71
Characters of biomorphs forming of several herbaceous perennials
Different types of biomorphes for several perennials growing in different ecological conditions are described using comparative morphological analysis. It is noted that biomorph's type of Fragaria vesca L. can change not only during ontogeny but at vegetative period also. Type of biomorph of Plantago lanceolata L. is defines by life form and site characteristics.
Текст научной статьи Особенности формирования биоморф у некоторых травянистых многолетников
У стойчивость любой системы как единого целого зависит от возможности системы и составляющих ее элементов изменяться во времени и пространстве. В основе структуры и динамики ценопопуляций лежат структура и динамика особей, которые определяются жизненными формами [9, 10, 20], типами биоморф растений [11, 18] и их положением в пространстве.
Термины «биоморфа» и «жизненная форма» часто используют как синонимы. Однако в нашей работе мы сочли необходимым разделить эти понятия. Жизненную форму мы рассматриваем в рамках эколого-морфологической концепции Варминга-Серебрякова. Жизненная форма - это внешний облик растений (габитус), который возникает в онтогенезе в результате роста растений в определенных экологических условиях. Тип габитуса определяется строением побегов, положением в пространстве и длительностью его жизни, а также строением побеговых систем.
Ряд исследователей рассматривают биоморфу как более широкое понятие, чем жизненную форму, подчеркивая ее фитоценотическую сторону согласно теории фитогенного поля А.А. Уранова [14]. Биоморфу мы рассматриваем в понимании О.В. Смирновой с соавторами [11] как демографическую единицу с разным количеством центров закрепления. По характеру пространственного размещения структурных частей (побегов, корней, почек возобновления) и степени их автономности выделены следующие биоморфы: моноцен-трический, неявнополицентрический, явнополицентрический, ацентрический [11, 18].
Объектами исследования были выбраны многолетние травянистые растения с дициклически-ми монокарпическими побегами и системой придаточных корней, гемикриптофиты: подорожник ланцетолистный ( Plantago lanceolata L. сем. Plantaginaceae Juss.) - короткокорневищный факультативно стержнекорневой или корнеотпрысковый поликарпик и земляника лесная ( Fragaria vesca L. сем. Rosaceae L.) - короткокорневищный наземно-столонообразующий поликарпик.
Цель работы - проанализировать формирование биоморф у модельных видов в различных местообитаниях.
МАТЕРИАЛЫ И МЕТОДЫ
Материал собирали на территории Республики Марий Эл (РМЭ). Исследования P. lanceolata проводили на суходольных лугах, пойменном лугу пастбищного использования, склоне мелиорационного канала и экотонно-опушечном ценозе, которые различались по механическому составу почв (песчаные, супесчаные, суглинистые, легкие супесчаные, щебень соответственно). Таким образом, нами был построен ряд местообитаний по механическому составу почв.
Сбор материала по изучению F. vesca проводили на территории Государственного природного заповедника «Большая Кокшага» РМЭ в сообществах, отличающихся по характеру почв и по типу фитоценоза: сосняк-можжевелово-ландышево-зеленомошный - почва - бурая, лесная, связнопесчаная на слоистых средних и крупнозернистых аллювиально-деллювиальных песках; экотонные сообщества: липняк осиновый - в ельнике-черничнике с примесью лиственных деревьев, почва - бурая, лесная, связно-песчаная грунтово-глееватая на слоистых песках; ежоворазнотравный луг - на опушке смешанного леса с почвой - дерново-среднеподзолистой, связнопесчаной, грунтово-глееватой на двучленных песчано-суглинистых породах; рудеральное сообщество на зарастающей насыпи железнодорожного полотна , с почвой слабо сформированной, где среди крупного известнякового щебня произрастает при полной фоновой освещенности F. vesca .
В работе использовали сравнительно-морфологический и популяционноонтогенетический методы [8, 15]. Жизненные формы модельных видов описаны по экологоморфологической классификации И.Г. Серебрякова [10], а типы биоморф - по экологодемографической классификации О.В. Смирновой с соавторами [11]. В качестве счетной единицы рассматривалась особь семенного и вегетативного происхождения.
Модельные виды, как в природе, так и в посадках могут быть однорозеточными и многорозеточными. Под однорозеточными мы понимаем однопобеговые растения с вегетативными ортотропными розеточными или генеративными ортотропными полурозеточными побегами. Многорозеточные особи представлены системой побегов или комплексом побегов замещения [1], состоящие из побега n-го порядка, за который мы принимали скелетный моноподиальный побег с собственной корневой системой [2] и всех боковых побегов разных порядков.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
Для более полной биоморфологической характеристики растений разных видов необходимо учитывать следующие признаки: структуру подземных органов, характер ветвления надземных и подземных побегов, наличие специализированных органов размножения, особенности пространственной структуры особи и степень вегетативной подвижности, способность к вегетативному размножению. Типы биоморф у объектов исследова- ния представлены разными жизненными формами и способны изменяться не только в различных местообитаниях (P. lanceolata), но и в течение вегетационного периода (F. vesca).
Высокая пластичность подземных органов у особей P. lanceolata в разных местообитаниях, в зависимости от механического состава почвы, приводит к крайним проявлениям морфологической поливариантности — вплоть до изменения жизненной формы [4].
Моноцентрический тип биоморф у P. lanceolata представлен:
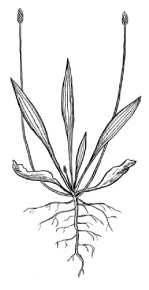
а) одно- и многорозеточными растениями стержнекорневой жизненной формы на песчаных и легких супесчаных почвах; б) одно- и многорозеточными короткокорневищно-кисте-корневыми особями — на суглинистой почве; в) стержнекор-невыми-короткокорневищными однорозеточными растениями — на щебне (рис. 1).
А
Б
Рис. 1. Моноцентрическая биоморфа у P. lanceolata:
А - стержнекорневая жизненная форма, Б - короткокорневищная жизненная форма
А

Б
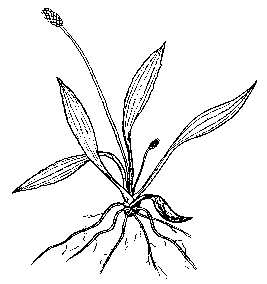
Рис. 2. Моноцентрический тип биоморф у F. vesca:
А - диаспора имматурного состояния; Б - особь субсенильного состояния
У F. vesca моноцентрическую биоморфу имеют диаспоры имматурного (im) состояния (рис. 2), которые развиваются на столонах (кистекорневой габитус), а также особи виргинильного (v), молодого генеративного (g1), средневозрастного (g2) и старого генеративного (g3), субсенильного (ss) и сенильного (s) состояний с короткокорневищной жизненной формой (рис. 2). В виде моноцентри-ческой биоморфы растения в виргинильном и генеративных онтогенетических состояниях суще- ствуют в начале вегетационного сезона, когда рост столонов только начинается, и в конце вегетационного периода, когда их появление прекращается. Однорозеточные и многорозеточные особи F. vesca субсенильного и сенильного онтогенетических состояний на протяжении всего вегетационного периода являются моноцентрическими.
Приземное расположение почек возобновления часто обусловлено деятельностью втягивающих корней растений [3, 10, 12, 13 и др.], как, например, у особей P. lanceolata. В пазухах листьев вегетативного розеточного побега закладывались почки, которые в дальнейшем могли стать подземными «скрытыми укороченными побегами» [6] или спящими почками, как их называют другие авторы [7, 10]. Повреждение верхушки побега растения n-го порядка (вытаптывание или скашивание), а также его отмирание стимулировало рост почек возобновления, а это, в свою очередь, приводило к формированию системы побегов или многорозеточности. Побеги замещения с собственной корневой системой можно рассматривать как единицы воздействия на среду.

А

Б
Рис. 3. Неявнополицентрическая биоморфа: А — P. lanceolata , Б — F. vesca
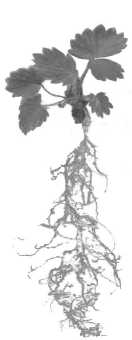
На экспериментальном участке МарГУ у особей P. lanceolata в случае гибели побега n-го порядка на живых подземных органах закладывались адвентивные почки. Вначале их развитие происходило под землей, а затем за счет удлинения междоузлий. Дочерние розеточные побеги выносились на поверхность почвы с образованием собственной корневой системы. Наличие нескольких близкорасположенных центров закрепления с перекрывающимися фитогенными полями [15] приводило к формированию неявнополицентрической биоморфы (рис. 3). Такой же тип биоморф нами был обнаружен у особей F. vesca. На главном побеге, залегающем глубоко в почве, из спящих почек формировались диаспоры, которые при помощи удлиненных междоузлий или «ложных» [19] столонов выносились в верхние слои почвы для поддержания жизнедеятельности особи. Когда на таком дочернем розеточном побеге начиналась формироваться собственная корневая система, а столоны у такого растения [5] не образовывались, это приводило к появлению несколько сближенных центров закрепления, и особь имела неявнополицентрическую биоморфу (рис. 3Б). Явнополицентрическая биоморфа у F. vesca формируется следующим образом: на плагиотропных побегах особей этого вида образуются диаспоры, в момент появления собственной корневой системы они закрепляются, сохраняя связь с материнской особью (рис. 4). Растения этой биоморфы существовали недолго, менее одного вегетационного сезона. Поэтому, придерживаясь взглядов А.П. Хохрякова [16], такой габитус можно считать фенобиоморфой, поскольку он регулярен, постоянен и закономерен краткосрочно. В конце лета наступала морфологическая дезинтеграция [11]: столоны отмирали и особь вновь становилась моно-

Рис. 4. Растение F. vesca явнополицентрической биоморфы
или неянополицентри-ческой.
Выделение переходного типа биоморф связано с неоднозначностью отнесения растений к тому или иному типу биоморф по ранее существующим классификациям [11, 18].
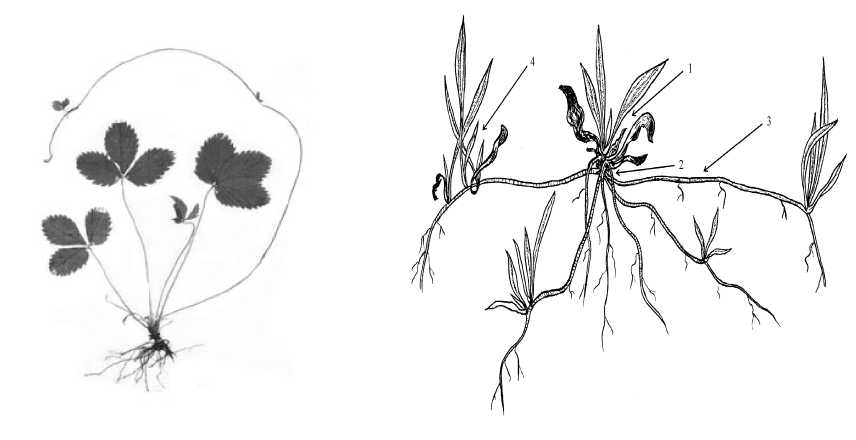
Впервые такой тип биоморфы был описан у F. vesca [17]. На определенном участке территории существовали особи F. vesca различных онтогенетических состояний и биоморф: моно- и неявнополицентрические. При переходе особей этих биоморф в явнополицентрические растения F. vesca существовали в виде системы, состоящей из особей со столонами и диаспорами на них. Особь с неукоренившимися диаспорами пока нельзя считать полицентрической. В итоге переходный тип у F. vesca сочетает черты моно- или неявнополицентрических особей с неукоренившимися диаспорами на столонах (рис. 5А).
Такой же тип биоморфы наблюдали и у особей P. lanceolata , произрастающих на склонах с нестабильным субстратом. Взрослые растения
P. lanceolata семенного происхождения имели выраженный центр сосредоточения биомассы, т.е. они принадлежат к моноцентрической би-оморфе. На многочисленных придаточных корнях, занимающих в пространстве векторное положение, по всей их длине неравномерно закладывались адвентивные почки, которые развивались в дочерние розеточные побеги. Следует отметить, что молодые диаспоры были представлены тронувшейся в рост почкой без собственной корневой системы, потому такая многорозеточная особь уже не является моноцентрической, но еще и не стала полицентрической. Несмотря на то, что корневая система еще не сформирована, мы счи таем, что надземная биомасса с почками возобновления оказывает влияние на среду. Поэтому такой тип биоморфы вполне справедливо назвать переходным (рис. 5 Б). При вегетативном размножении (механический разрыв коммуникаций или их перегнивание) судьба отделившихся рамет может складываться по-разному. Неукоренившие-ся раметы с небольшим участком придаточного корня могут погибнуть или образовать собственную корневую систему и далее функционировать как моноцентрические растения, а в случае сохранения связи диаспор с материнской особью и формирования собственной корневой системы — явнополицентрические.
А
Б
Рис. 5. Переходный тип биоморф у особей P. lanceolata (А) и F. vesca (Б): 1 - материнский розеточный побег; 2 - короткое корневище; 3 — придаточный корень;
4 - дочерний розеточный побег
Сходный тип биоморфы наблюдается у корнеотпрысковых растений, например: щавелька малого ( Rumex acetosella L.), осота огородного ( Sonchus oleraceus L.), льнянки обыкновенной ( Linaria vulgaris Mill.) и др. Растение может иметь переходный тип биоморфы короткий отрезок времени как, например, у наземно-столонообразующих или в определенном онтогенетическом состоянии - у длиннокорневищных и корнеотпрысковых растений.
Продолжительность пребывания растений в той или иной биоморфе у разных видов будет индивидуальна.
ЗАКЛЮЧЕНИЕ
Анализ материала выявил три типа биоморф у F. vesca (моноцентрический, неявнополицентрический и явнополицентрический) и два типа у P. lanceolata (моно- и неявнополицентрический). Изучение особенности размещения корней, побегов и почек возобновления и их укоренения в пределах особей на модельных видах подтвердили необходимость выделения переходного типа биоморф у наземно-столонообразующих и корнеотпрысковых растений. Следовательно, переход ный тип биоморф представлен исходным центром закрепления (моно- или неявнополицентрическим) с неукоренившимися диаспорами.
Список литературы Особенности формирования биоморф у некоторых травянистых многолетников
- Бологова В.Л. Функциональные и динамические аспекты макроморфологической структуры растительного организма на примернее ежи сборной: Автореф. дис. … канд. биол. наук. М., 1989. 21 с.
- Гатцук Л.Е. Элементы структуры жизненных форм геммаксилярных растений и биоморфологический анализ копеечника кустарникового (Hedysarum fruticosum Pall.). Автореф. дис. … канд. биол. наук. М. 1970. 21 с.
- Голубев В.Н. К онтогенезу корневищ кистекорневых растений//Бот. журн. 1956. Т. 41, № 2. С. 248-253.
- Жукова Л.А., Османова Г.О. Морфологическая пластичность подземных органов Plantago lanceolata L. (Plantaginaceae L.)//Бот. журн. 1999. Т. 86, № 12. С. 80-86.
- Коржинский С.И. Гетерогенез и эволюция (предварительное сообщение)//Изв. Имп. акад. наук. 1899. Т. X, № 3. С. 255-268.
- Любименко В.Н. О спящих почках//Тр. С.-Петербург, о-ва естествоиспытат. Отд. Ботан. 1900. Т. 30, Вып. 3. С. 195-259.
- Лященко Н.И. О спящих почках кустарников//Бот. журн. 1958. Т. 43, № 7. С. 1039-1047.
- Работнов Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах//Тр. БИН АН СССР. Сер. 3, Геоботаника. М; Л, 1950. Вып. 6. С. 7-204.
- Серебряков И.Г. Морфология вегетативных органов высших растений. М.: Наука, 1952. 391 с.
- Серебряков И.Г. Экологическая морфология растений. М: Высш. шк., 1962. 378 с.
- Смирнова О.В., Заугольнова Л.Б., Торопова Н.А. и др. Критерии выделения возрастных состояний и особенности хода онтогенеза у растений различных биоморф//Ценопопуляции растений. М.: Наука, 1976. 215 с.
- Таршис Г.И. Подземные органы многолетних травянистых растений. Уч. пособие. Из-во. Свердл. гос. пед. ин-та, 1975. С. 132.
- Таршис Л.Г. Структурное разнообразие подземных органов высших растений. Екатеринбург: УрО РАН, 2003. 196 с.
- Уранов А.А. Фитогенное поле//Проблемы современной ботаники. М.; Л. 1965. Т. 2. С. 251-254.
- Уранов А.А. Возрастной состав фитоценопопуляций как функции времени и энергетических волновых процессов//Биол. науки. 1975. № 2. С. 17-29.
- Хохряков А.П. Активная, а не пассивная жизненная форма//Природа. 1994. № 6. С. 36-41.
- Шивцова И.В. Эколого-морфологические особенности особей и организация популяций Fragaria vesca L.: Автореф. дис. … канд. биол. наук. Сыктывкар, 2008. 23 с.
- Шорина Н.И. Строение зарослей папоротника-орляка в связи с его морфологией//Жизненные формы: структура, спектры и эволюция. М.: Наука, 1981. С. 213-232.
- Hackel E. Die natűrlichen Pflanzenfamilien/A. Engler, K. Prantl. Gramineae. 1887. T. II. Abt 2.
- Warming E. Über perenne gewachse//Bot. Centralblatt. 1884. Bd. 18, № 19. S. 16-22.
