Особенности формирования мезофауны в 40-летних лесных культурах на старопахотной серой почве

Автор: Егунова М.Н., Безкоровайная И.Н.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки: Почвоведение
Статья в выпуске: 6, 2015 года.
Бесплатный доступ
В статье рассматривается эдификаторная роль основных лесообразующих пород Сибири в формировании комплекса почвенных беспозвоночных в процессе формирования подстилочного слоя и изменения гидротермических условий верхнего горизонта почвы.
Плотность, мезофауна, почвенные беспозвоночные, лесная культура, подстилка, анализ, формирование комплекса
Короткий адрес: https://sciup.org/14084343
IDR: 14084343 | УДК: 631.468
The peculiarities of mesofauna formation in 40-year-old forest cultures on the old-arable gray soil
The edificator role of the main Siberian forest-forming species in the formation of the soil in vertebrate complex within the process of the underlayer formation and changes in the hydrothermal conditions of the upper soil horizon are considered in the article.
Текст научной статьи Особенности формирования мезофауны в 40-летних лесных культурах на старопахотной серой почве
Введение . Экологами и почвоведами давно признано влияние растительности на формирование свойств почв [5, 6, 12, 13].
Несмотря на это, понимание механизмов взаимодействия в системе растительность↔почва до сих пор остается неполным. Причиной этого является тот факт, что принципы распространения растительности отражают ландшафтный уровень вариабельности почв, обусловленный другими устойчивыми факторами почвообразования (возраст, климат, рельеф, почвообразующие породы), которые мешают вычленить влияние растений на развитие почвы. Немногочисленные многолетние эксперименты с лесными культурами, когда разные древесные виды высаживаются в близких климатических и почвенных условиях, дают возможность минимизировать смешанное влияние факторов и вычленить особенности влияния растительности на почву [11–13].
Для изучения внутренних связей в системе растительность↔почва в 1968 г. в условиях южной тайги (56ос.ш. 92ов.д.) под руководством проф. Н.В. Орловского заложен эксперимент с основными лесообразующими породами Сибири на старопахотных серых почвах [10]. Эксперимент с лесными культурами на выровненном агрофоне является идеальным объектом для изучения динамики формирования лесных насаждений разного породного состава. Он дает возможность выявить роль растительности в организации и функционировании отдельных компонентов лесного сообщества.
Растительность оказывает влияние на почву через гетеротрофные процессы, в том числе через функциональную активность почвенных животных. В связи с этим анализ формирования ком- плексов беспозвоночных при изучении взаимодействия между растительностью и почвой имеет важное значение.
Цель исследования. Определить особенности формирования комплексов почвенных беспозвоночных группы «мезофауна» в 40-летних лесных культурах, созданных в одинаковых почвенноклиматических условиях.
Объекты и методы исследования. Исследования проводились на многолетнем эксперименте в 40-летних лесных культурах основных лесообразующих пород Сибири – сосне сибирской (Pinus sibirica) , ели (Picea obovata) , лиственнице (Larix sibirica) , сосне (Pinus silvestris) , березе (Betula fruticosa) и осине (Populus tremula) . Экспериментальный участок расположен на обширной террасе р. Качи, левобережного притока Енисея, и занимает 1,7 га на серой почве. Культуры высажены 2– 3-летними саженцами в 1971–1972 гг. на участок, который после раскорчевки вторичного березняка лесостепной зоны периодически использовался под сельскохозяйственные угодья [10].
На эксперименте периодически проводятся лесоводственные мониторинговые исследования процесса формирования лесных сообществ разного типа. К 40-летнему возрасту культур под ними сформировался равномерно распределенный по поверхности органогенный горизонт с четко выраженной стратиграфией из подгоризонтов L, F и Н, а гомогенный пахотный слой серой почвы (PY-горизонт) дифференцировался (O-AY1-AY2) [11].
В качестве фонового участка в работе использована разнотравно-злаковая залежь (возраст 5 лет), отражающая состояние сообщества до посадки культур. Залежь представлена злаковой растительностью – пырей ( Elytrigia repens L.) , мятлик ( Poa annua L.) и разнотравьем – тысячелистник ( Achillea millefolium L.) , мышиный горошек ( Vicia cracca L.) , ромашка непaхучая ( Matricaria perforata (L.) Sch. Bip.) , клевер белый ( Trifolium repens L.) , гулявник лезеля ( Sisymbrium loeselii L.) .
Сосняк разнотравно-осочковый (90 лет) и березняк орляково-разнотравно-осочковый (60 лет) отражают конечную стадию формирования лесных биогеоценозов разного типа.
Все контрольные участки примыкают к экспериментальному участку с лесными культурами и находятся в тех же почвенно-климатических условиях.
Для учета почвенных беспозвоночных группы «мезофауна» использовались прямые методы, общепринятые в почвенно-зоологических исследованиях: послойный отбор проб с последующей разборкой на колонке почвенных сит [4]. К мезофауне отнесены дождевые черви, энхитреиды (учитывались только особи, извлекаемые при ручной разборке проб), личинки и имаго членистоногих. В каждом типе насаждений пробы (размер 25х25 см) отбирались в 5-кратной повторности по двум слоям: подстилка и минеральный слой 0–15 см. Полевые исследования проводились в 2011–2013 гг.
Результаты исследования и их обсуждение. Запасы подстилки в 40-летних лесных культурах близки естественным лесным сообществам (рис. 1). В культурах хвойных пород запасы подстилки составляют 2348–3870 г/м2 [3, 8], а под березой и осиной они в 2–3 раза ниже, что отражает высокую степень биологической трансформации растительного вещества в лиственных сообществах.
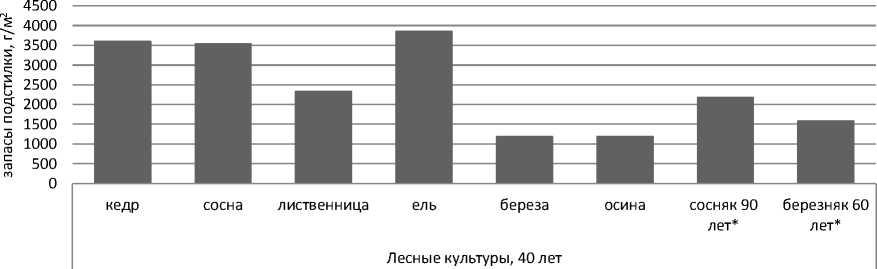
Рис. 1. Запасы подстилки в 40-летних лесных культурах и естественных лесных экосистемах*: сосняк разнотравно-осочковый (90 лет) и березняк орляково-разнотравно-осочковый (60 лет)
Анализ гидротермических условий под формирующимися лесными культурами показал, что различия по влагозапасам между культурами в почвенном слое в среднем за вегетационный период незначительны [1]. Наибольшие влагозапасы характерны для лиственных насаждений (рис. 2). Выявленные различия по температуре почвы между 30-летними культурами обусловлены контрастностью фитоценотических характеристик исследуемых биотопов (густота и плотность, возраст, сомкнутость, запасы надземной фитомассы, напочвенный покров и лесная подстилка), поскольку температурный режим почв под насаждениями является результирующим эффектом тепловых потоков в почву и из нее.
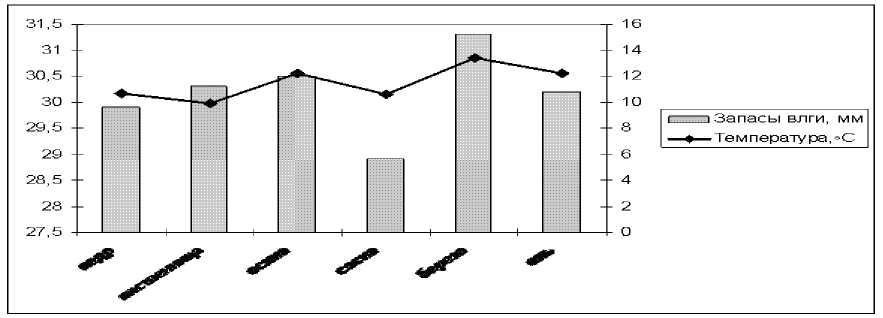
Рис. 2. Запасы влаги и температура минерального слоя 0–10 см под лесными культурами (по данным Безкоровайной, Яшихина, 2003), мм
Анализ комплексов почвенных мезофауны под 40-летними культурами и на контрольных участках показал, что наибольшая плотность крупных беспозвоночных отмечена под сосной – 359 экз/м2, а наименьшая под березой – 47,1 экз/м2 (рис. 3). Различия в формирующихся под древесными породами комплексах педобионтов связаны с подстилочным горизонтом. Выявлена достаточно тесная корреляция между запасами подстилки и плотностью беспозвоночных в подстилочном слое (R – 0,56). Основная плотность беспозвоночных (50–80 %) сосредоточена в подстилке, что отличает искусственные от естественных биогеоценозов, где большой процент педобионтов приурочен к верхнему минеральному слою почвы и превышает таковую под культурами в 5–7 раз. Такой тип вертикального размещения характерен для серых почв, где формирование верхнего аккумулятивного горизонта происходит через вовлечение педобионтами растительных остатков в минеральную часть почвы и их разложение непосредственно в ней [7].
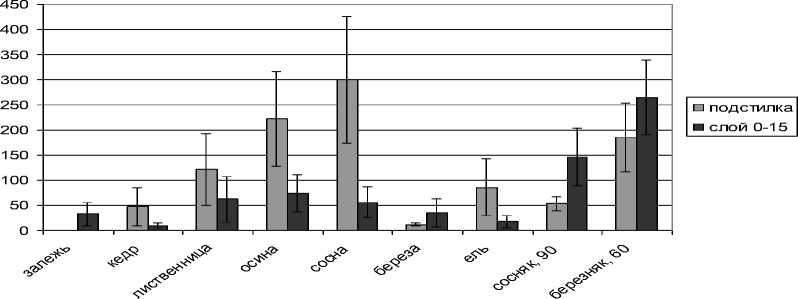
Рис. 3. Плотность почвенных беспозвоночных в 40-летних культурах, экз/м2
В минеральном слое почвы под культурами плотность беспозвоночных близка к залежи и колеблется от 9 до 74 экз/м2. Полученные данные по вертикальной стратификации крупных беспозвоночных показали, что изменение их плотности под культурами происходит за счет формирования подстилки и, соответственно, подстилочного комплекса беспозвоночных.
При анализе состава таксономических групп почвенной мезофауны под 40-летними культурами выявлено доминирование дождевых червей (Lumbricidae) практически под всеми 40-летними культурами. Исключение составляют культуры лиственницы и осины, где более 40 % приходится на альтернативную группу почвенных сапрофагов энхитреид (Enchytraedae). Для естественных лесных сообществ выявлено абсолютное доминирование энхитреид (80 %) (табл.1).
Таблица 1
Плотность мезофауны основных таксонов под культурами и на контрольных участках, экз/м2
|
Группа бес- |
Кедр |
Лиственница |
Осина |
Сосна |
Береза |
Ель |
Залежь |
Сосняк, 90 |
Березняк, 60 |
|
позвоночных |
Лесные культуры |
Контроль |
|||||||
|
Lumbricidae |
25,7 |
56,4 |
82 |
200, 6 |
38,0 |
61, 8 |
10,8 |
15,5 |
35, 6 |
|
Lithobiidae |
8,2 |
35, 6 |
36, 8 |
29,5 |
2,6 |
0 |
1,6 |
8, 7 |
26, 7 |
|
Diptera |
10,4 |
20, 7 |
8, 9 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Nematoda |
0 |
0 |
0 |
0 |
0 |
0 |
13,7 |
0 |
0 |
|
Enchytraeidae |
8,9 |
62,2 |
123 |
119, 6 |
1, 6 |
8, 9 |
0 |
157, 9 |
335,0 |
|
Aranei |
0 |
0 |
13, 3 |
0 |
0,7 |
1, 6 |
0 |
8, 7 |
0 |
|
Julida |
0 |
0 |
10, 9 |
0 |
0 |
28,5 |
1, 6 |
8, 7 |
26,6 |
|
Gastropoda |
0 |
8, 9 |
10, 9 |
0 |
1,1 |
0 |
0 |
0 |
0 |
|
Coleoptera |
3 |
0 |
10, 9 |
8, 9 |
3,1 |
3,0 |
4, 9 |
0 |
26,6 |
Почвообразовательный потенциал беспозвоночных тесно связан с их трофической специализацией [2, 9]. Накопление растительных остатков на поверхности почвы приводит к значительным изменениям в трофической структуре педокомплексов крупных беспозвоночных. До посадки культур в комплексе мезофауны, по данным В.К. Дмитриенко (1984), доминировали зоофаги – 79 %, на фитофаги приходилось 17, на сапрофаги – 4% [10]. Спустя 40 лет доминирующей группой под всеми культурами, как и в естественных лесных биогеоценозах, являются сапрофаги (80–95%), что свидетельствует о формировании типичных лесных комплексов мезофауны (рис. 4). Под культурами сапрофаги представлены дождевыми червями, кивсяками, брюхоногими, личинками двукрылых. На зоофаги (многоножки костянки и пауки) приходится 6–14 %, миксофитофаги (нематоды, личинки жесткокрылых) – от 9 до 30 %. На залежи миксофитофаги являются доминирующей группой (57 %), а сапрофаги составляют не более 38 %.
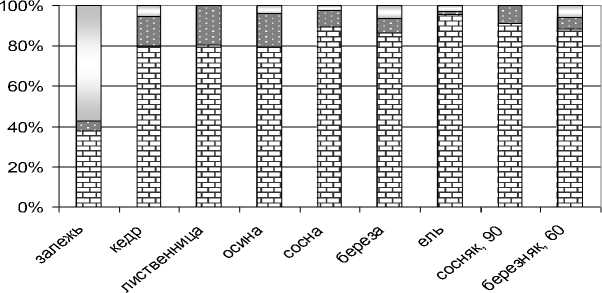
о миксофитофаги о хищники а сапрофаги
Рис. 4. Соотношение трофических групп почвенных беспозвоночных на 40-летних лесных культурах, %
Факторный анализ выделил три группы сообществ, различающихся между собой по групповому составу комплексов почвенной мезофауны: залежь, лесные культуры и естественные лесные биогеоценозы; это свидетельствует о том, что комплексы почвенных беспозвоночных в 40-летних культурах находятся еще на стадии своего формирования, занимая промежуточное положение между залежью и естественными лесными сообществами (рис. 5). Кроме того, для лесных культур четко выявлены различия в структуре комплексов беспозвоночных между хвойными и лиственными древесными породами. Примыкание в эксперименте к хвойным породам секции березы объясняет появление под её пологом подроста ели и сосны. Факторный анализ, направленный на выделение содержательной части факторов, выявил, что основной вклад в различие комплексов под лесными культурами разных древесных пород вносят группы дождевых червей (46,1%) и энхитреид (41,4 %) (табл. 2).
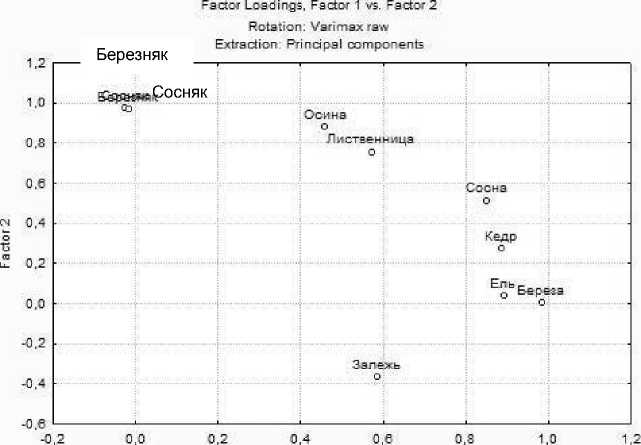
Рис. 5. Факторный анализ групп крупных почвенных беспозвоночных
Таблица 2
|
Тип растительности |
Factor 1 Lumbricidae |
Factor 2 Enchytraedae |
|
Контроль |
||
|
Залежь |
0,586 |
-0,366 |
|
Сосняк |
-0,026 |
0,975 |
|
Березняк |
-0,018 |
0,971 |
|
Лесные культуры |
||
|
Кедр |
0,885 |
0,274 |
|
Лиственница |
0,571 |
0,759 |
|
Осина |
0,459 |
0,884 |
|
Сосна |
0,851 |
0,516 |
|
Береза |
0,984 |
0,004 |
|
Ель |
0,897 |
0,040 |
|
Собственное значение |
4,152 |
3,729 |
|
Информационный вклад, % |
46,134 |
41,398 |
Достоверность различий комплексов почвенных беспозвоночных между исследуемыми участками подтверждена двухфакторным дисперсионным анализом. Показаны достоверные различия между культурами и разнотравно-злаковой залежью как по численности, так и по структуре комплексов мезофауны (Р <0,04), между культурами и естественными лесными биогеоценозами только по структуре (Р <0,03).
A. Szuiecki (1983), анализируя формирование почвенной мезофаны в лесной посадке на вырубках, пришел к выводу, что ее регенерация после вырубок включает в себя три периода развития насаждений: создание лесных культур, стадия смыкания крон молодняков и 20–30-летние жердняки [14]. В конце последнего периода лесная фауна, как правило, стабилизируется и остается в таком состоянии с небольшими изменениями в последующие средневозрастные, приспевающие, спелые и перестойные периоды развития насаждения. Исходя из различий по качественному и количественному составу почвенных беспозвоночных в культурах и естественных лесных биогеоценозах, можно предположить, что комплексы почвенных беспозвоночных в 40-летних культурах частично сохраняют в себе аграрные черты серой почвы, выведенной из-под леса, и еще не достигли стадии стабилизации. Скорее всего, при создании лесных культур на старопахотной почве, которая длительное время была вовлечена в сельскохозяйственное пользование, формирование педокомплек-сов беспозвоночных происходит путем первичной сукцессии, в то время как в почвах, где беспозвоночные временно исключаются из привычной экологической обстановки (пожары, рубки и т.п.), это происходит путем вторичного естественного возобновления сообществ.
Заключение. Формирование комплекса почвенных беспозвоночных формаций древесных пород происходит через опосредованное влияние фитоценотических особенностей каждого биотопа (породного состава, сомкнутости крон, запасов подстилки и гидротермических условий почвы). Запасы влаги в минеральном слое, наряду с таким фактором, как концентрация органического вещества, обуславливают вертикальное размещение почвенных животных.
Преобладание сапрофагов находится в прямой зависимости от сформированности подстилочного слоя. Где почва не имеет выраженной подстилки, сапрофаги уступают по числу и биомассе почвенным животным с другим типом питания. В свою очередь, увеличение доли сапрофагов указывает на приближение состава педокомплексов мезофауны культур к естественным лесным сообществам.
По структуре и плотности почвенных беспозвоночных 40-летние лесные культуры занимают промежуточное положение между начальной стадией восстановительного процесса и естественными лесными биогеоценозами.
Выявлена эдификаторная роль хвойных и лиственных видов древесных пород в формировании структуры комплексов беспозвоночных через доминирование двух основных групп беспозвоночных, перерабатывающих органическое вещество почвы, – дождевых червей и энхитреид.
Список литературы Особенности формирования мезофауны в 40-летних лесных культурах на старопахотной серой почве
- Безкоровайная И.Н., Яшихин Г.И. Влияние гидротермических условий почвы на комплексы беспозвоночных в хвойных и лиственных культурах//Экология. -2003. -№ 1. -С. 56-62.
- Бызов Б.А. Зоомикробные взаимодействия в почве. -М.: ГЕОС, 2005. -212 с.
- Ведрова Э.Ф., Решетникова Т.В. Масса подстилки и интенсивность ее разложения в 40летних культурах основных лесообразующих пород Сибири//Лесоведение. -2014. -№ 1. -С. 42-50.
- Гиляров М.С. Зоологический метод диагностики почв. -М.: Наука, 1965. -287 с.
- Зонн С.В. Влияние леса на почву. -М.: Наука, 1954. -189 с.
- Карпачевский Л.О. Структура почвенного покрова и разнообразие лесных фитоценозов//Почвоведение. -1996. -№ 6. -С. 722-27.
- Курчева Г.Ф. Роль почвенных животных в разложении и гумификации растительных остатков. -М.: Наука, 1971. -156 с.
- Решетникова Т.В. Лесные подстилки как депо биогенных элементов.//Вестник КрасГАУ. -2011. -№ 12. -С. 74-81.
- Стриганова Б.Р. Питание почвенных сапрофагов. -М.: Наука, 1980. -243 с.
- Моделирование развития искусственных лесных биогеоценозов/Л.С. Шугалей, М.Г. Семечкина, Г.И. Яшихин . -Новосибирск: Наука, 1984. -152 с.
- Шугалей Л.С. Влияние лесных культур на свойства плантажированной почвы//Почвоведение. -2002. -№ 3. -С. 345-354.
- Binkley D., Giardina C. Why do trees species affect soils? The warp and woof of tree-soil interactions//Biogeochemistry. -1998. -№ 42. -P. 89-106.
- Linking litter calcium, earthworms, and soil properties: a common garden test with 14 temperate tree species/J. Oleksyn, J. Modrzynski, P. Mrozinski //Ecology Letters 8. -2005. -Р. 811-818.
- Szuiecki A. Comparison of macrofauna of afforested field with that of forest soil//The process of forest soil macrofauna formation after afforestation of farmland. -Warsaw Agricultural University Press, 1983. -P. 116-122.
