Особенности формирования растительного покрова и пространственная структура макрозообентоса в условиях нарушения гидрологического режима водоема (на примере Березовcкого залива Воткинского пруда)
")
Автор: Капитонова О.А., Каргапольцев И.А., Мельников Д.Г.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 1-1 т.13, 2011 года.
Бесплатный доступ
Представлены результаты изучения водной и прибрежно-водной растительности Березовского залива Воткинского пруда (Удмуртская Республика). Показано, что в условиях нарушения гидрологического режима водоема в результате строительства дамбы происходит его обмеление, эвтрофирование, ускоренное зарастание. Вслед за изменением характера растительного покрова залива изменяется пространственная структура экотопически связанных с ней сообществ донных макробеспозвоночных. Выявленные изменения структуры растительности и макрозообентоса индицируют ухудшение экологического состояния экосистемы изученного водного объекта.
Макрофиты, водная и прибрежно-водная растительность, макрозообентос, гидробионты, воткинский пруд, удмуртская республика
Короткий адрес: https://sciup.org/148205510
IDR: 148205510 | УДК: 574.5(470.51)(045)
Features of formation of vegetative cover and spatial structure of macrozoobenthos in the conditions of infringement of reservoir hydrological mode (on an example of Beryozovka gulf of Votkinsk pond)
Results of study of aquatic vegetation of gulf Beryozovka of Votkinsk pond (the Udmurt Republic) are presented. It is shown that dike building in the gulf mouth has broken the reservoir hydrological mode that leads to its shallowing. The gulf is revealed to be quickly overgrown with aquatic plants. After having changed the gulf vegetation character the spatial structure of the communities of bottom macroinvertebrates connected with it alters. The revealed changes of the vegetation and the macrozoobenthos structure indicate deterioration of ecological state of the aquatic ecosystem.
Текст научной статьи Особенности формирования растительного покрова и пространственная структура макрозообентоса в условиях нарушения гидрологического режима водоема (на примере Березовcкого залива Воткинского пруда)
У силивающиеся антропогенные воздействия на водоемы и водотоки приводят к различного рода изменениям в структурно-функциональных характеристиках экосистем, в ряде случаев необратимым. Поскольку невозможно представить, что в обозримом будущем созидательная активность человечества резко сократится или прекратится вовсе, возникает необходимость поиска способов оптимальной эксплуатации водных экосистем, которые обеспечивали бы их неистощимое развитие. Это возможно только при умении прогнозировать направления изменений в этих экосистемах. Разработка методов и способов прогнозирования представляет собой особую задачу, решение которой возможно с учетом представлений о сложных биотических процессах, протекающих в экосистемах, их качественных и количественных изменениях на фоне влияния разнообразных факторов окружающей среды [1]. Для этого знания о том, как устроена экосистема, недостаточны. Необходимо иметь четкие представления о взаимоотношениях между отдельными компонентами экосистемы, понимать механизмы, обеспечивающие их внутренние взаимосвязи, уметь выражать их количественно.
Известно, что при зарастании мелководий водоёмов макрофитами видовое разнообразие пелагических гидробионтов увеличивается за счет фи-тофильной фауны [3, 5]. Показано также [4, 7], что в различных сообществах водных растений могут присутствовать разные виды и комплексы гидробионтов. Фитофильная фауна в совокупности с макрофитами образует единую комплексную систему, участвующую в процессах самоочищения водных экосистем и поддержания их
структурно-функциональных и продукционнодинамических свойств [10, 18]. Тем не менее, особенности распределения макрозообентоса в водоеме, связь его с водной и прибрежно-водной растительностью, а также изменение пространственной структуры вследствие антропогенного воздействия изучены к настоящему времени крайне слабо. Между тем подобные сведения могут быть полезны, как в теоретическом отношении при изучении экологии донных макробеспозвоночных, особенно стенобионтных и редких таксонов, их распределения в пространстве, связях с растительностью водоемов и водотоков, так и в практическом плане при поиске оптимальных путей эксплуатации водных объектов, а также разработке способов биологической очистки загрязненных поверхностных водных источников.
ЦЕЛЬ, МАТЕРИАЛЫ И МЕТОДЫ
Цель настоящей работы — изучение пространственной структуры макрофитной растительности и экотопически связанных с ней организмов макрозообентоса в условиях зарегулированного водного объекта. Исследования проводились в течение вегетационных сезонов 2002, 2005, 2006 и 2009 гг. с использованием стандартных методов гидробиологических [6, 13, 17] и гидроботаниче-ских [9, 15] исследований. Объектами исследований являлись водная и прибрежно-водная растительность, а также донные макробеспозвоночные Березовского залива Воткинского пруда (г. Воткинск, Удмуртская Республика).
Воткинский пруд создан в 1759 г. на р. Вотка для нужд Воткинского железоделательного завода. Площадь водного зеркала составляет 18,8 км2, пруд вытянут по долине р. Вотки на 13 км, максимальная ширина — 2 км, максимальная глубина — 15 м, полный объем водной массы — 85 млн. м3. Вода характеризуется как гидрокарбонатно-кальциево-магниево-натриевая, общая минерализация варьируется в пределах 157-361 мг/л [20]. Березовский залив пруда представляет собой устьевой участок р. Березовки — левобережного притока Воткинского пруда — отгороженный в 80-х годах XX в. от основной части пруда насыпной дамбой, которая перегородила залив, фактически превратив его в обособленный водоем. Протяженность залива составляет 600 м, максимальная ширина — около 150 м (в нижней части у дамбы), средняя глубина — 1,2 м, максимальная — 1,9 м (в нижней русловой части). С момента строительства дамбы по настоящее время наблюдается ускоренное зарастание и обмеление залива, что рассматривается как следствие нарушения его гидрологического режима. Ежегодные темпы заиления составляют 154 м3/год, или 2,0-2,5 см/год. Максимальная мощность донных отложений в пределах залива составляет 5,5 м, преобладают отложения мощностью 3,5-4,5 м [16].
По материалам геоботанических описаний составлена картосхема зарастания исследованного залива (рис. 1). Для выяснения особенностей пространственной структуры макрозообентоса качественные и количественные данные гидробиологических проб сопоставлялись с характером зонального распределения макрофитной растительности, при этом учитывалось проективное покрытие растений (ППР), которое ранжировалось по следующей схеме: ранг 1 придавался сообществам с ППР менее 10%, ранг 2 — с ППР 1030%, ранг 3 — с ППР 30-50%, ранг 4 — с ППР 5070%, ранг 5 — с ППР 70-90% и ранг 6 — с ППР 90-100%. Видовой состав донных макробеспозвоночных, сопряженных с определенными растительными формациями, сравнивали с использованием коэффициента сходства Съеренсена-Чекановского (Ksc). Дополнительно рассчитаны индекс Шеннона (H) и показатель выравненности по Пиелу (Е).
Исследования макрозообентоса проводились с июня по сентябрь, в период вегетации водной растительности. В Березовском заливе было заложено 5 трансект, на каждой трансекте было установлено 6-7 станций, на которых ежемесячно проводились сборы донных макробеспозвоночных. Сборы проб бентоса на мягких грунтах проводили трубчатым дночерпателем с площадью захвата 0,01 м2. Каждая проба включала содержимое 4 дночерпателей. Макрозообентос выбирали сразу, помещали в герметичную посуду и фиксировали 4%-ным раствором формальдегида [6, 13]. Всего в ходе проведенных исследований было отобрано 135 количественных проб гидробионтов, выявлено обитание в пределах изученной акватории 162 видов и таксонов более высокого ранга донных макробеспозвоночных из 71 семейства, выделено 53 ассоциации из 30 формаций водной, прибрежно-водной и береговой растительности на основе подходов и принципов доминантной системы классификации растительности.
Математическая обработка гидробиологического материала проводилась с применением стандартных методов и приемов биометрии [11] в табличном редакторе Microsoft ® EXCEL 2003, однофакторный дисперсионный и корреляцион ный анализы проводили с использованием пакета программ STATlSTICA 6.0 ® for Windows.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Растительность Березовского залива. Целенаправленное изучение растительного покрова Березовского залива Воткинского пруда начато нами в 2002 г., результаты флористических исследований опубликованы в 2003 г.[8]. Система высшей водной и прибрежно-водной растительности залива, составленная с учетом геоботанических описаний за весь период наблюдений, представлена следующими синтаксонами.
-
I. ТИП РАСТИТЕЛЬНОСТИ. ВОДНАЯ РАСТИТЕЛЬНОСТЬ - AQUIPHYTOSA
-
А. Группа классов. Настоящая водная (гидрофит-ная) растительность - Aquiphytosa genuina.
-
I. Класс формаций. Настоящая водная (гидрофит-ная) растительность - Aquiphytosa genuina.
-
1. Группа формаций гидрофитов, свободно плавающих в толще воды — Aquiherbosa genuina demersa natans.
-
1. Формация ряски трехдольной - Lemneta trisulcae (Асс.: 1) Lemnetum trisulcae).
-
2. Формация роголистника темно-зеленого - Cera-tophylleta demersi (Асс.: 2) Ceratophylletum demersi).
-
3. Формация пузырчатки обыкновенной - Utricu-larieta vulgaris (Асс.: 3) Utricularietum vulgaris).
2. Группа формаций погруженных укореняющихся гидрофитов — Aquiherbosa genuina submersa radicans.
-
4. Формация рдеста блестящего - Potameta lucentis (Асс.: 4) Potametum lucentis; 5) Potametum perfoliati-lucentis; 6) Hydrophytoso-Potametum lucentis).
-
5. Формация рдеста пронзеннолистного - Potameta perfoliati (Асс.: 7) Potametum perfoliati).
-
6. Формация рдеста сплюснутого - Potameta com-pressi (Асс.: 8) Potametum compressi).
-
7. Формация мелколистных рдестов - Potameta pu-silli (Асс.: 9) Potametum berchtoldii; 10) Potametum friesii; 11) Potametum pusilli; 12) Potametum trichoides).
-
8. Формация урути мутовчатой - Myriophylleta ver-ticillati (Асс.: 13) Hydrophytoso-Myriophylletum verticil-lati).
-
9. Формация шелковников - Batrachieta (Асс.: 14) Batrachietum circinati; 15) Hydrophytoso-Batrachietum circinati).
-
10. Формация болотника обоеполого - Callitricheta hermaphroditicae (Асс.: 16) Callitrichetum hermaphrodi-ticae).
-
11. Формация элодеи канадской - Elodeeta canden-sis (Асс.: 17) Elodeetum candensis).
3. Группа формаций укореняющихся гидрофитов с плавающими на воде листьями — Aquiherbosa genuina radicans foliis natantibus.
-
12. Формация кубышки малой - Nuphareta pumila (Асс.: 18) Nupharetum pumila).
-
13. Формация кубышки Спеннера - Nuphareta spenneriana (Асс.: 19) Nupharetum spenneriana).
-
14. Формация кувшинки северной - Nymphaeeta borealis (Асс.: 20) Nymphaeetum borealis; 21) Lemno-Nymphaeetum borealis).
4.
Группа формаций свободно плавающих на поверхности воды гидрофитов — Aquiherbosa genuina natans.
-
15. Формация ряски маленькой с многокоренником - Lemno minori-Spirodeleta (Асс.: 22) Lemno minori-Spirodeletum; 23) Hydrophytoso-Lemno minori-Spirodeletum).
-
16. Формация водокраса лягушачьего - Hydro-charieta morsus-ranae (Асс.: 24) Lemno-Hydrocharietum morsus-ranae).
Б. Группа классов. Прибрежно-водная растительность — Aquiherbosa vadosa.
-
II. Класс формаций. Воздушно-водная (гелофитная) растительность — Aquiherbosa helophyta.
-
5. Группа формаций низкотравных гелофитов — Aquiherbosa helophyta humilis.
-
17. Формация сусака зонтичного — Butometa umbellate (Асс.: 25) Hydrophytoso-Butometum umbellate).
-
18. Формация стрелолиста обыкновенного — Sagit-tarieta sagittifoliae (Асс.: 26) Sagittarietum sagittifoliae; 27) Butomo-Sagittarietum sagittifoliae; 28) Hydrophy-toso-Sagittarietum sagittifoliae).
-
19. Формация ежеголовника всплывшего — Sparga-nieta emersi (Асс.: 29) Sparganietum emersi).
-
20. Формация ежеголовника мелкоплодного — Spar-ganieta microcarpi (Асс.: 30) Hydrophytoso-Sparganietum microcarpi).
-
21. Формация хвоща приречного — Equiseteta fluvi-atilis (Асс.: 31) Equisetetum fluviatilis; 32) Heteroher-boso-Equisetetum fluviatilis).
6. Группа формаций высокотравных гелофитов — Aquiherbosa helophyta procera.
-
22. Формация камыша озерного — Scirpeta lacustris (Асс.: 33) Heteroherboso-Scirpetum lacustris).
-
23. Формация рогоза узколистного — Typheta angus-tifoliae ( Асс.: 34) Typhetum angustifoliae; 35) Lemno-Typhetum angustifoliae; 36) Heteroherboso-Typhetum angustifoliae).
-
24. Формация рогоза широколистного — Typheta latifoliae (Асс.: 37) Lemno-Typhetum latifoliae; 38) Het-eroherboso-Typhetum latifoliae).
-
25. Формация манника большого — Glycerieta maximae (Асс.: 39) Glycerietum maximae; 40) Ranuncu-loso lingua-Glycerietum maximae; 41) Lemno trisulcae-Glycerietum maximae; 42) Heteroherboso-Glycerietum maximae).
-
26. Формация тростника южного — Phragmiteta australis (Асс.: 43) Phragmitetum australis; 44) Equiseto fluviatilis-Phragmitetum australis; 45) Menyantheto trifo-liata-Phragmitetum australis; 46) Heteroherboso-Phragmitetum australis).
-
27. Формация осоки острой — Cariceta acutae (Асс.: 47) Caricetum acutae; 48) Equiseto fluviatilis-Caricetum acutae; 49) Scolochloeto festucacea-Caricetum acutae; 50) Menyantheto trifoliata-Caricetum acutae).
-
28. Формация вахты трехлистной — Menyantheta trifoliate (Асс.: 51) Menyanthetum trifoliate).
-
II. ТИП РАСТИТЕЛЬНОСТИ. БЕРЕГОВАЯ РАСТИТЕЛЬНОСТЬ - RIPAROPHYTOSA.
III. Класс формаций. Гигрогелофитная растительность — Aquiherbosa hygrohelophyta.
-
B. Группа классов. Древесно-кустарниковотравянистая растительность - Riparophytosa arbo-frutescenso-herbosus.
-
IV. Класс формаций. Травянистая береговая растительность — Riparophytosa herbosus.
-
29. Формация лютика ползучего — Ranunculeta repens (Асс.: 52) Heteroherboso-Ranunculetum repens).
-
30. Формация камыша лесного — Scirpeta sylvestris (Асс.: 53) Scirpetum sylvestris).
Таким образом, растительность Березовского залива Воткинского пруда представлена 53 ассоциациями, объединенными в 30 формаций из 2 типов растительности. Наибольшим разнообразием по числу синтаксонов отличается настоящая водная растительность, включающая 24 ассоциации (табл. 1). Учитывая небольшую площадь акватории залива, это свидетельствует о весьма благоприятных условиях для развития гидрофитов, чему способствует небольшая глубина залива, достаточно высокая степень защиты от волнобоя высокими укрепленными берегами и хорошо развитым поясом высокотравных гелофтов, практически по всему периметру окаймляющих водоем. Возведение дамбы в устье залива также способствует его зарастанию водными растениями, поскольку тем самым были созданы благоприятные для многих гидрофитов затишные условия.
Не намного по числу ассоциаций отстает от гидрофитной растительности воздушно-водная (гелофитная) растительность, однако существенно отличается от нее по числу формаций. Среди воздушно-водной растительности наибольшее синтаксономическое разнообразие отмечено для высокотравных гелофитов (5 формаций, 14 ассоциаций). Низкотравные гелофиты при таком же числе формаций включают лишь 8 ассоциаций. Такое соотношение синтаксонов может указывать на происходящие процессы заболачивания и обмеления, при которых пояс высокотравных гелофитов широким фронтом «наступает» на пояс погруженных и плавающих гидрофитов, вытесняя также и низкотравные гелофиты.
Таблица 1. Синтаксономический состав растительности Березовского залива Воткинского пруда
|
Класс формаций |
Число формаций |
% |
Число ассоциаций |
% |
|
Настоящая водная (гидрофитная) растительность |
16 |
53,3 |
24 |
45,3 |
|
Воздушно-водная (гелофитная) растительность |
10 |
33,3 |
22 |
41,5 |
|
Гигрогелофитная растительность |
2 |
6,7 |
5 |
9,4 |
|
Травянистая береговая растительность |
2 |
6,7 |
2 |
3,8 |
|
Всего: |
30 |
100,0 |
53 |
100,0 |
Гигрогелофитная растительность не отличается большим разнообразием. Ассоциации этого класса формаций встречаются преимущественно в верхней части залива, где активно идут процессы сплавинообразования и заболачивания. Травянистая береговая растительность отмечена на ограниченных участках акватории и слабо представлена в растительном покрове залива. Причиной этому можно считать упомянутый выше характер берегов, отличающихся значительной крутизной.
Наши наблюдения за изменением флоры и растительности залива свидетельствуют об ускорении за последние годы процессов его зарастания (рис. 1), обмеления и антропогенного эвтро-фирования, что рассматривается нами как следствие зарегулирования этого водного объекта, в результате чего гидрологический режим водоема оказался нарушен. Наблюдаемое распределение водных и прибрежно-водных растений на акватории рассматриваемого водоема связано преимущественно с эколого-биологическими особенностями растений-ценозообразователей (эдификато- ров), приуроченных к определенным глубинам, донным отложениям и степени проточности. Так, на небольших глубинах (0,5-0,9 м), наиболее распространенных в заливе, обычны сообщества болотника обоеполого (Callitriche hermaphroditica L.), рдестов Фриса (Potamogeton friesii Rupr.), сплюснутого (P. compressus L.), гребенчатого (P. pectina-tus L.) и блестящего (P. lucens L.), кубышки малой (Nuphar pumila (Timm) DC.). Глубины от 0,9 до 1,0 м заняты формациями стрелолиста обыкновенного (Sagittaria sagittifolia L.), который, благодаря гетерофилии, способен проникать и на большие глубины, а также урути мутовчатой (Myriophyllum verticillatum L.) и шелковника завитого (Batrachium circinatum (Sibth.) Spach). На средних глубинах (0,8-1,3 м) развиты сообщества рдеста блестящего (Potamogeton lucens), элодеи канадской (Elodea canadensis Michx.), роголистника темно-зеленого (Ceratophyllum demersum L.). На максимальных глубинах залива (1,5-1,7 м) образуют ценозы пузырчатка обыкновенная (Utricu-laria vulgaris L.), рдест пронзеннолистный (Potamogeton perfoliatus L.), кувшинка северная (Nym-phaea x borealis E. Camus.), а также кубышка Спеннера (Nuphar x spenneriana Gaudin). В центральной части водоема плотный ковер образуют сообщества рясковых (Lemna minor L., L. turionif-era Landolt, L. trisulca L., Spirodela polyrhiza (L.) Schleid.).
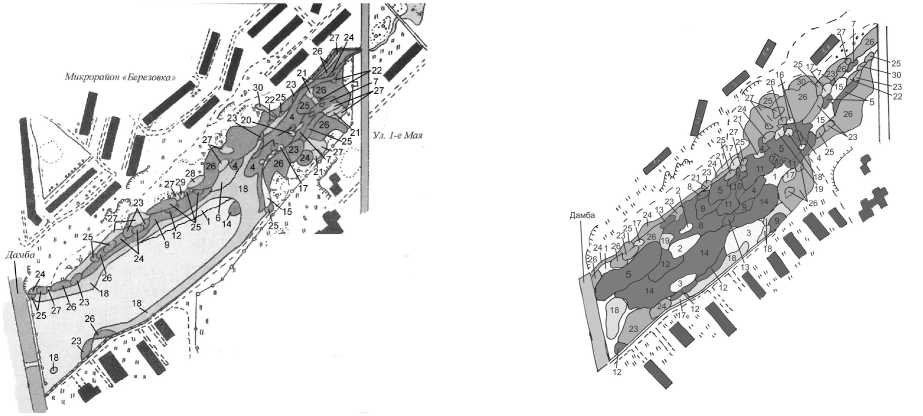
а
б
Рис. 1. Схема зарастания Березовского залива Воткинского пруда (а — 2002 г., б — 2009 г.):
1 - Lemneta trisulcae, 2 - Ceratophylleta demersi, 3 - Utricularieta vulgaris, 4 - Potameta lucentis, 5 - Potameta per-foliati, 6 - Potameta compressi, 7 - Potameta pusilli, 8 - Myriophylleta verticillati, 9 - Batrachieta, 10 - Callitricheta hermaphroditicae, 11 - Elodeeta canadensis, 12 - Nuphareta pumila, 13 - Nuphareta spennerianae, 14 - Nymphaeeta borealis, 15 - Lemno minori-Spirodeleta, 16 - Hydrocharieta morsus-ranae, 17 - Butometa umbellati, 18 - Sagittarieta sagittifoliae, 19 - Sparganieta emersi, 20 - Sparganieta microcarpi, 21 - Equiseteta fluviatilis, 22 - Scirpeta lacustris, 23 - Typheta angustifoliae, 24 - Typheta latifoliae, 25 - Glycerieta maximae, 26 - Phragmiteta australis, 27 - Cariceta acutae, 28 - Menyantheta trifoliata, 29 - Ranunculeta repens, 30 - Scirpeta sylvestris
На прибрежных мелководьях развиты сообщества гелофитов и гигрогелофитов. Здесь доминируют тростник обыкновенный ( Phragmites australis (Cav.) Trin. ex Steud.), рогозы широколистный ( Typha latifolia L.) и узколистный ( T. angustifolia L.), манник большой ( Glyceria maxima (C. Hartm.) Holmb.), стрелолист обыкновенный. Меньшие площади заняты ценозами хвоща приречного ( Equisetum fluviatile L.), ежеголовника всплывшего ( Sparganium emersum Rehm.), камыша озерного ( Scirpus lacustris L.), сусака зонтичного ( Butomus umbellatus L.), осоки острой ( Carex acuta L.), вахты трехлистной ( Menyanthes trifoliata L.).
Таким образом, основными доминантами мак-рофитной растительности Березовского залива являются широколистные и узколистные рдесты, кувшинковые, рясковые, элодея, шелковник, стрелолист, тростник южный, рогозы, манник большой. Общее проективное покрытие сообществ, образованных этими видами, составляет 70-100%. Их активное участие в формировании растительного покрова рассматриваемого водного объекта свидетельствует о продолжающихся процессах заболачивания прибрежий и обмеления центральной части акватории залива. На сегодняшний день площадь зарастания залива составляет около 95-97%, что на треть превышает этот показатель в 2002 г. [16].
Видовой состав и пространственная структура макрозообентоса. Распределение организмов макрозообентоса в пространстве определяется рядом биотических и абиотических факторов, среди которых к наиболее важным следует отнести пространственную структуру макрофитной растительности. Высокая мозаичность растительного покрова акватории Березовского залива обуславливает обитание в нем различных систематических и экологических групп донных макробеспозвоночных. Согласно результатам однофакторного дисперсионного анализа, при увеличении проективного покрытия водных и прибрежноводных растений в гидробиологических пробах падает доля личинок двукрылых по численности (р < 0,01) (рис. 2а). В то же время выявлено возрастание в пробах доли моллюсков (рис. 2б) и нимф стрекоз с увеличением проективного покрытия макрофитов (р < 0,01). Для брюхоногих моллюсков это связано в первую очередь с обильной кормовой базой, представленной как собственно макрофитами, так и их обрастателями — перифитоном. Увеличение доли стрекоз происходит за счет нимф подотряда равнокрылых, являющихся в основном фитофильными организмами.
За все время наблюдений наибольшее количество видов донных макробеспозвоночных было выявлено в формациях рдестов блестящего (102 вида) и пронзеннолистного (96), элодеи канад ской (72), стрелолиста (78), шелковника (67). По-видимому, помимо широкого распространения в заливе сообществ, образованных этими макрофитами, в них складываются наиболее благоприятные условия для жизни гидробионтов. Сходные с нашими результаты были получены В.Н. Столбу-новой [19], показавшей зависимость видового состава зоопланктона от характера зарастания мелководной зоны водоемов, в частности от степени развития рдестовых зарослей. Наименьшее количество видов донных макробеспозвоночных зафиксировано в формациях хвоща приречного, вахты, манника большого, осоки острой, рогозов узколистного и широколистного (от 18 до 26 видов).
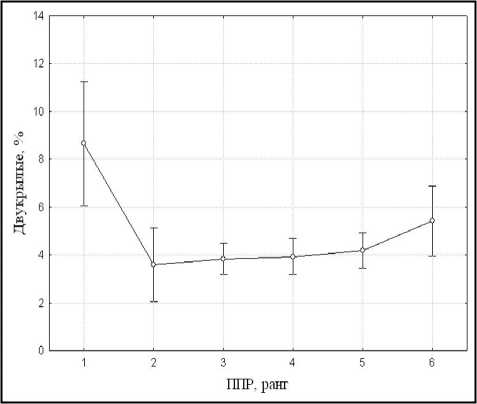
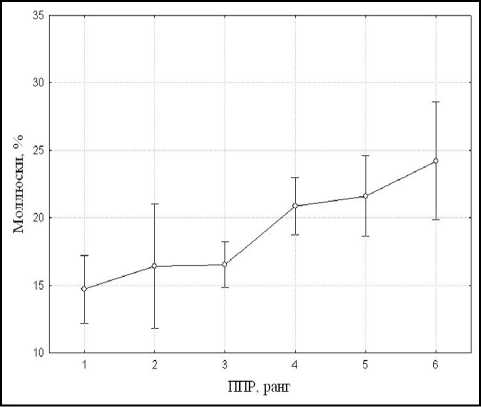
б
а
Рис. 2. Изменение средней доли личинок двукрылых (а) и брюхоногих моллюсков (б) в гидробиологических пробах при увеличении проективного покрытия макрофитов
В изученных растительных формациях видовое ядро макрозообентоса, представленное группой наиболее обильных видов, составляют стрекозы (Erythromma najas, E. viridulum, Coenagrion hastulatum, C. armatum, Epitheca bimaculata, Cordu-lia aenea), брюхоногие моллюски (Lymnaea lagotis, Anisus vortex, Euglesa sp.), пиявки (Helobdella stag-nalis, Glossiphonia complonata), комары-звонцы подсемейства Chironominae, малощетинковые черви (Limnodrilus hoffmeisteri, Nais simplex, Stylaria lacustris). Наибольшие средние значения числен ности и биомассы донных макробеспозвоночных наблюдались в формациях полностью погруженных в воду гидрофитов (рдестов пронзеннолистного и блестящего, элодеи канадской), а также в сообществах рясковых, наименьшие — в формациях гидрофитов с плавающими на поверхности воды листьями и низкотравных гелофитов. Эта же закономерность наблюдалась для индексов Шеннона и выравненности по Пиелу (табл. 2).
Таблица 2. Средние значения численности и биомассы макрозообентоса в основных растительных формациях Березовского залива Воткинского пруда
|
Растительные формации |
Численность, тыс. экз./м2 |
Биомасса, г/м2 |
H (бит/экз.) |
Е |
|
Хвоща приречного |
0,9 |
29,1 |
1,5 |
0,5 |
|
Осоки острой |
1,2 |
11,5 |
1,6 |
0,4 |
|
Рогоза широколистного |
16,8 |
156,4 |
2,4 |
0,7 |
|
Рогоза узколистного |
18,2 |
268,2 |
2,6 |
0,6 |
|
Тростника южного |
10,5 |
46,1 |
2,2 |
0,6 |
|
Манника большого |
12,1 |
76,4 |
1,9 |
0,6 |
|
Стрелолиста обыкновенного |
15,7 |
374,6 |
2,7 |
0,8 |
|
Сусака зонтичного |
6,3 |
98,1 |
2,1 |
0,5 |
|
Шелковника завитого |
27,2 |
307,8 |
2,6 |
0,8 |
|
Рдеста блестящего |
78,6 |
317,7 |
2,9 |
0,8 |
|
Рдеста пронзеннолистного |
84,2 |
329,2 |
2,6 |
0,6 |
|
Элодеи канадской |
76,7 |
296,8 |
2,5 |
0,8 |
|
Кувшинки северной |
5,4 |
132,7 |
1,9 |
0,8 |
|
Кубышки малой |
2,8 |
118,2 |
1,6 |
0,6 |
|
Кубышки Спеннера |
1,3 |
14,6 |
1,5 |
0,4 |
|
Ряски маленькой |
43,9 |
78,3 |
2,4 |
0,6 |
Полученные нами данные по пространственной структуре макрозообентоса в пределах исследованной акватории в целом согласуются с результатами исследований ряда авторов [2, 21], свидетельствующими о зависимости распределения фитофильной фауны от зонально-поясного характера произрастания макрофитной растительности. По-видимому, в густых зарослях прибрежно-водных растений (осоки, хвощи, камыши, рогозы, манник, тростник) создаются неблагоприятные условия (небольшая глубина, затенение, значительный прогрев воды) для жизнедеятельности гидробионтов, в связи с чем фауна водных беспозвоночных в таких зарослях существенно обедняется видами. Кроме того, известно, что некоторые виды воздушно-водных макрофитов, в частности, рогозы и тростники, обладают сильно выраженными аллелопатическими свойствами, ингибирующими развитие фито- и зоопланктона,
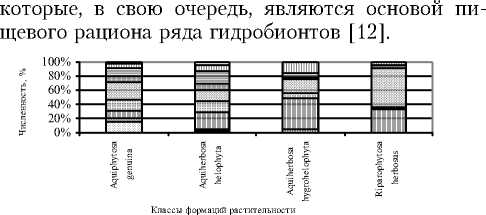
Odonata Heteroptera Diptera
Ephemeroptera Hirudinea другие
Рис. 3. Эколого-ценотические спектры макрозообентоса в исследованных классах растительных формаций Березовского залива Воткинского пруда
Результаты сравнения видового состава фауны донных макробеспозвоночных показали высокий уровень сходства изученных групп гидробионтов, функционально связанных с формациями Potameta lucentis, Potameta perfoliati, Potameta pu-silli, Elodeeta canadensis, Sagittarieta sagittifolii, Batrachieta (Ksc = 0,4-0,8), т.е. принадлежащих к одному классу растительности. Наименьшее сходство наблюдалось при сравнении фаунистических комплексов, биотопически связанных с различными классами и группами формаций (Ksc = 0,20,4). Выявлено, что сообщества макрозообентоса не являются специфичными для какой-либо растительной формации. Ассоциации и формации макрофитов отличаются между собой, главным образом, количественными показателями функционально связанных с ними макробеспозвоночных. Качественные отличия проявляются лишь при сравнении групп и классов формаций (рис. 3). На этих уровнях прослеживаются не только количественные, но и таксономические различия в структуре бентоса, что связано с влиянием на видовой состав донных беспозвоночных не только абиотических, но и биотических факторов. В этой связи комплексы донных беспозвоночных, топически и трофически зависимых от характера по- ясно-зонального распределения растительности в акватории водоема, следует рассматривать в качестве отдельных экологических групп макрозообентоса, различающихся не только таксономически, но и функционально, пространственно структурированных, играющих собственные роли в экосистеме водного объекта.
Вместе с тем в пределах сообществ ряда формаций водной и прибрежно-водной растительности некоторые виды донных макробеспозвоночных встречались очень редко и в небольшом количестве.
Так, в формациях рдеста пронзеннолистного встречены единичные экземпляры Ranatra linearis, Nepa cinerea, Dytiscus semisulcatus , в формации элодеи канадской — Hydraena sp. В ценозах стрелолиста обыкновенного выявлены единичные особи Donacia dentate , в сообществах рогозов широколистного и узколистного — Donacia semicu-prea , манника большого — Georissus crenulatus . Среди зарослей Sagittaria sagittifolia и Potamogeton lucens встречены нимфы Parapoynx stratiotata , в зарослях Nymphaea х borealis обнаружена Elophia nymphaeata , а в сообществах Nuphar pumila — Acentria ephemerella . Многие из отмеченных гидробионтов являются стенотрофными организмами [14], предпочитая селиться на определенных видах водных растений.
ЗАКЛЮЧЕНИЕ
Активизация процессов зарастания и связанное с ним обмеление и заболачивание акватории Березовского залива Воткинского пруда, вызванное нарушением гидрологического режима р. Березовки вследствие неправильного проектирования и строительства в ее устье дамбы, ведут к пространственному перераспределению организмов макрозообентоса. Функционально связанные посредством трофических и топических взаимоотношений с высшими водными и прибрежноводными растениями, фитофильные донные макробеспозвоночные образуют наиболее плотные сообщества в фитоценозах формаций настоящей водной растительности, в связи с чем их таксономическое разнообразие и обилие выше в тех частях водоема, которые подвергаются зарастанию гидрофитами, особенно погруженными видами. В пределах акватории Березовского залива — это его центральная часть и верховья. Напротив, прибрежные мелководья с доминированием сообществ высокотравных и низкотравных гелофитов, а также гигрогелофитов в меньшей степени заселяются организмами макрозообентоса, что, возможно, связано с выраженными аллелопатическими свойствами некоторых видов прибрежноводных растений, сильным затенением и дефицитом кислорода, вызванным гниением растительного опада. Последующее обмеление акватории залива, которое со временем приведет к расширению площадей прибрежно-водной и береговой растительности и, соответственно, сокращению сообществ погруженных растений, будет способствовать сокращению таксономического разнооб- разия и обилия донных макробеспозвоночных, среди которых имеются как широко распространенные таксоны, так и редкие стенотрофные виды, связанные с определенными видами макрофитов. По-видимому, следует также ожидать увеличения обилия организмов-переносчиков опасных заболеваний, что позволяет рассматривать проблему заболачивания рассматриваемого водоема с точки зрения санитарноэпидемиологической безопасности жителей г. Воткинска. Сохранение экологической и таксономической структуры макрозообентоса, выполняющего вкупе с водными и прибрежно-водными растениями важные средоформирующие функции, представляется возможным при оптимизации эксплуатации Березовского залива, предполагающую в первую очередь реконструкцию дамбы, что должно привести к увеличению проточности воды в заливе и замедлению его зарастания.
ÑÏÈÑÎÊ ËÈÒÅÐÀÒÓÐÛ
-
1. Алимов А.Ф. Элементы теории функционирования водных экосистем. СПб.: Наука, 2000. 147 с.
-
2. Баканов А.И., Столбунова В.Н, Довбня И.В., Жгарева НН, Законнов В.В. Животное население зарослей оз. Неро. Фауна растительных ассоциаций // Биол. внутр. вод. 2001, № 2. С. 43-52.
-
3. Баканов А.И., Столбунова В.Н., Жгарева Н.Н. Животное население озера Неро. Связи между компонентами экосистемы / Биол. внутр. вод. 2001. № 3. С. 48-55.
-
4. Зимбалевская ЛН. Экологические группировки фауны зарослей Днепра // Гидробиол. журн. 1966. Т. 2. № 5. С. 34-41.
-
5. Зимбалевская ЛН. Распределение фитофильных беспозвоночных и методы их количественного учета // Гидробиол. журн. 1972. Т. 8, № 2. С. 49-55.
-
6. Жадин В.И. Методика гидробиологического исследования: Учеб. пос. М.: Высш. шк., 1960. 189 с.
-
7. Жгарева Н.Н. Фауна зарослей / Экосистема малой реки в изменяющихся условиях среды. М.: Товарищество научных изданий КМК, 2007. С. 249-268.
-
8. Капитонова О.А., Мельников Д.Г. Флора Березовского залива Воткинского пруда (Удмуртская Республика) // Вести. Удм. ун-та. Сер. Биология. 2003. С. 21-32.
-
9. Катанская В.М. Высшая водная растительность континентальных водоемов. Л.: Наука, 1981. 187 с.
-
10. Кокин К.А. Экология высших водных растений. М.: МГУ, 1982. 160 с.
-
11. Лакин Г.Ф. Биометрия: Учеб. пособие для биол. спец. вузов. М.: Высш. шк., 1990. 352 с.
-
12. Мережко А.И., Кузьменко М.И, Величко И.М. Взаимоотношения различных видов водорослей и высших водных растений, обусловленные их метаболитами // Летучие биологически активные соединения биогенного происхождения. М., 1971. 152 с.
-
13. Методические рекомендации по сбору и обработке материалов при гидробиологических исследованиях на пресноводных водоемах. Зообентос и его продукция / Под ред. Ю.А. Барулина. Л.: ГОСНИОРХ, 1984. 51 с.
-
14. Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 5. Высшие насекомые / Под ред. С.Я. Цалолихина. Спб.: Наука, 2001. 836 с.
-
15. Папченков В.Г. Растительный покров водоемов и водотоков Среднего Поволжья, Ярославль: ЦМП МУБиНТ, 2001. 214 с.
-
16. Разработка программы мероприятий по оздоровлению Березовского залива Воткинского пруда: Научный отчет. Ижевск, 2002. 148 с.
-
17. Руководство по методам гидробиологического анализа поверхностных вод и донных отложений / Под ред. В.А. Абакумова. Л.: Гидрометеоиздат, 1983. 239 с.
-
18. Садчиков А.П., Кудряшов М.А. Экология прибрежноводной растительности. М.: Изд-во НИА-Природа, РЭФИА, 2004. 220 с.
-
19. Столбунова В.Н. Особенности зоопланктона мелководий верхневолжских водохранилищ и условия его существования // Зооценозы водоемов бассейна Верхней Волги в условиях антропогенного воздействия. СПб: Гидрометеоиз-дат, 1993. С. 20-38.
-
20. Удмуртская Республика: Энциклопедия. Ижевск: Изд-во «Удмуртия», 2008. 768 с.
-
21. Зорта-Сахарова К.Е. Фiтофiльна макрофауна водойм та водотокiв ониззя Дунаю як шдикатор пх еколопчного стану: Автореферат дис. ... канд. бiол. наук. Кипв, 2009. 24 с.
FEATURES OF FORMATION OF VEGETATIVE COVER AND SPATIAL STRUCTURE OF MACROZOOBENTHOS IN THE CONDITIONS OF INFRINGEMENT
OF RESERVOIR HYDROLOGICAL MODE
(ON AN EXAMPLE OF BERYOZOVKA GULF OF VOTKINSK POND)
Список литературы Особенности формирования растительного покрова и пространственная структура макрозообентоса в условиях нарушения гидрологического режима водоема (на примере Березовcкого залива Воткинского пруда)
- Алимов А.Ф. Элементы теории функционирования водных экосистем. СПб.: Наука, 2000. 147 с.
- Баканов А.И., Столбунова В.Н., Довбня И.В., Жгарева Н.Н., Законнов В.В. Животное население зарослей оз. Неро. Фауна растительных ассоциаций//Биол. внутр. вод. 2001, № 2. С. 43-52.
- Баканов А.И., Столбунова В.Н., Жгарева Н.Н. Животное население озера Неро. Связи между компонентами экосистемы/Биол. внутр. вод. 2001. № 3. С. 48-55.
- Зимбалевская Л.Н. Экологические группировки фауны зарослей Днепра//Гидробиол. журн. 1966. Т. 2. № 5. С. 34-41.
- Зимбалевская Л.Н. Распределение фитофильных беспозвоночных и методы их количественного учета//Гидробиол. журн. 1972. Т. 8, № 2. С. 49-55.
- Жадин В.И. Методика гидробиологического исследования: Учеб. пос. М.: Высш. шк., 1960. 189 с.
- Жгарева Н.Н. Фауна зарослей/Экосистема малой реки в изменяющихся условиях среды. М.: Товарищество научных изданий КМК, 2007. С. 249-268.
- Капитонова О.А., Мельников Д.Г. Флора Березовского залива Воткинского пруда (Удмуртская Республика)//Вестн. Удм. ун-та. Сер. Биология. 2003. С. 21-32.
- Катанская В.М. Высшая водная растительность континентальных водоемов. Л.: Наука, 1981. 187 с.
- Кокин К.А. Экология высших водных растений. М.: МГУ, 1982. 160 с.
- Лакин Г.Ф. Биометрия: Учеб. пособие для биол. спец. вузов. М.: Высш. шк., 1990. 352 с.
- Мережко А.И., Кузьменко М.И., Величко И.М. Взаимоотношения различных видов водорослей и высших водных растений, обусловленные их метаболитами//Летучие биологически активные соединения биогенного происхождения. М., 1971. 152 с.
- Методические рекомендации по сбору и обработке материалов при гидробиологических исследованиях на пресноводных водоемах. Зообентос и его продукция/Под ред. Ю.А. Барулина. Л.: ГОСНИОРХ, 1984. 51 с.
- Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 5. Высшие насекомые/Под ред. С.Я. Цалолихина. Спб.: Наука, 2001. 836 с.
- Папченков В.Г. Растительный покров водоемов и водотоков Среднего Поволжья, Ярославль: ЦМП МУБиНТ, 2001. 214 с.
- Разработка программы мероприятий по оздоровлению Березовского залива Воткинского пруда: Научный отчет. Ижевск, 2002. 148 с.
- Руководство по методам гидробиологического анализа поверхностных вод и донных отложений/Под ред. В.А. Абакумова. Л.: Гидрометеоиздат, 1983. 239 с.
- Садчиков А.П., Кудряшов М.А. Экология прибрежно-водной растительности. М.: Изд-во НИА-Природа, РЭФИА, 2004. 220 c.
- Столбунова В.Н. Особенности зоопланктона мелководий верхневолжских водохранилищ и условия его существования//Зооценозы водоемов бассейна Верхней Волги в условиях антропогенного воздействия. СПб: Гидрометеоиздат, 1993. С. 20-38.
- Удмуртская Республика: Энциклопедия. Ижевск: Изд-во «Удмуртия», 2008. 768 с.
- Зорiна-Сахарова К.Е. Фiтофiльна макрофауна водойм та водотокiв ониззя Дунаю як iндикатор ïх екологiчного стану: Автореферат дис.... канд. бiол. наук. Киïв, 2009. 24 с.
