Особенности гибридных зон хромосомных рас обыкновенной бурозубки, Sorex araneus L., с разным уровнем кариотипической дифференциации

Автор: Черепанова Елена Владимировна, Гайдученко Елена Сергеевна, Крищук Ирина Александровна, Борисовец Ирина Павловна, Борисов Юрий Михайлович
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Статья в выпуске: 4, 2014 года.
Бесплатный доступ
На севере Тверской обл. обнаружены узкие гибридные зоны трех мономорфных хромосомных рас (Москва, Зап. Двина и Селигер) обыкновенной бурозубки, Sorex araneus, различающихся между собой по 3-4 диагностическим метацентрикам. Локализация гибридных зон совпадает с краевыми образованиями вепсовской стадии оледенения. Узость гибридных зон (от 0,5 до 2,5 км) обусловлена ограниченной областью совместного обитания этих кариотипически дифференцированных рас, что, вероятно, связано с конкурентными отношениями между ними. Диагностические метацентрики расы Зап. Двина, мономорфной до Смоленской возвышенности, обнаружены в популяциях расы Борисов на территории Беларуси. В популяциях рас Борисов и Bialowieza на территории Беларуси выявлена клинальная изменчивость - снижение частоты диагностических метацентриков - в направлении междуречья Днепра и Припяти. Вблизи гибридной зоны между полиморфными популяциями с диагностическими хромосомами расы Bialowieza и расы Киев обнаружена высокая частота акроцентриков. Особенности полиморфных популяций в междуречье Днепра и Припяти объясняются существованием на этой территории в прошлом популяций бурозубок с полностью акроцентрическим кариотипом. Кариотип, в котором все диагностические хромосомы представлены акроцентриками и считающийся исходным в эволюции S. araneus, обнаружен во многих полиморфных популяциях данного региона.
Рефугиумы, робертсоновские (rb-) транслокации, хромосомные расы, акроцентрическая раса, полиморфизм, клинальная изменчивость
Короткий адрес: https://sciup.org/146116540
IDR: 146116540 | УДК: 576:315.599.323.4
Specific features of hybrid zones of chromosomal races of the common shrew, Sorex araneus L., at different levels of karyotypic divergence
Narrow hybrid zones of three monomorphic chromosomal races of the common shrew, Sorex araneus, differing from each other by 3 - 4 diagnostic Robertsonian translocations, were revealed in the north of Tver Region. Localization of these hybrid zones follows marginal formation of the Veps stage glacier. A small width of hybrid zones (0,5 - 2,5 km) is determined by a small area of co-habitation of these karyotypically diverged races, which is caused, probably, by concurrent relations between them. Diagnostic metacentrics of the West Dvina race, which is monomorphic from the Valdai to the Smolensk Heights, were noted in populations of the Borisov race on the territory of Belarus (specific feature of this race is the absence of metacentric ip trait for the West Dvina race). A cline variability of the diagnostic metacentric frequencies, decreasing towards the Dnieper and Pripyat junction, was noted in populations of the Borisov and Bialowieza races on the territory of Belarus. A high acrocentric frequency near the hybrid zone between populations with diagnostic metacentrics of the Bialowieza race and the Kiev race was noted. The specific features of the polymorphic populations at the Dnieper and Pripyat junction can be explained by the existence in the past on the mentioned area the common shrew populations with completely acrocentric karyotype. Karyotype with all the diagnostic chromosomes represented by acrocentrics, regarded as primary in the evolution of S. araneus, was revealed in a number of polymorphic populations in the discussed region
Текст научной статьи Особенности гибридных зон хромосомных рас обыкновенной бурозубки, Sorex araneus L., с разным уровнем кариотипической дифференциации
Введение. Стабильная структура популяций мелких млекопитающих при внутривидовом и внутрипопуляционном
Работа проведена при финансовой поддержке РФФИ (грант 14-04-90006Бел-я).
полиморфизме определяется множеством факторов, как генетических, так и экологических. Процессы формирования внутривидовой популяционной структуры удобно изучать на хромосомнополиморфных видах. Обыкновенная бурозубка, Sorex araneus L., характеризующаяся огромным размахом межпопуляционного и внутрипопуляционного хромосомного полиморфизма, является одним из удобных объектов популяционной генетики. В кариотипе S. araneus три пары аутосом (af, be, tu) и половые хромосомы (XX у самок, XYY у самцов) мономорфны, а хромосомы j и I могут присутствовать как в акроцентрической, так и в метацентрической (//) форме. В приделах ареала вида известно более 70 хромосомных рас, которые различаются по Робертсоновским транслокациям акроцентрических хромосом (Rb) и занимают определенные территории (Hausser , 1994; Wojcik et al., 2002, 2003; Щипанов и др., 2009).
Кариотип, в котором 10 пар аутосом представлены акроцентриками (g, h, i, k, m, n, o, p, q, r\ считается исходным в эволюции 5. araneus (Wojcik et al., 2002). Такой кариотип был выявлен только в эндемичных популяциях горных изолятов Балкан (хромосомная раса Pelister) и Альп (раса Cordon) (Wojcik et al., 2002). Фиксация метацентрических хромосом 5. araneus могла происходить в малочисленных изолированных популяциях, например, в рефугиумах ледниковых областей (Орлов и др., 2007; Orlov, Borisov, 2007; Orlov et al., 2007).
Хромосомные расы, населяющие Восточную Европу и Сибирь, мономорфны и различаются между собой по 3-5 фиксированным диагностическим метацентрикам (Орлов и др., 2004, 2008; Щипанов и др., 2009). Такие хромосомные расы образуют узкие (шириной 0,5-5 км) гибридные зоны с комплексными гетерозиготами, в мейозе которых возникают сложные фигуры конъюгирующих хромосом - цепи или кольца (поливаленты). Поэтому плодовитость гетерозигот понижена (Jadwiszczak, Banaszek, 2006; Бородин, Поляков, 2008). У хромосомных рас, населяющих Зап. и Центр. Европу, как правило, фиксированы только 1-2 метацентрика, а 2 - полиморфны. Соседние расы различаются, чаще всего, присутствием у одной расы метацентрика и акроцентрического варианта - у другой расы (Орлов и др., 2008; Wojcik et al., 2002; Zima et al., 2003). Между такими полиморфными популяциями образуются широкие гибридные зоны. В мейозе гибридов-гетерозигот образуются триваленты, не влияющие на плодовитость. Предполагается, что полиморфизм по числу метацентриков, наблюдаемый во многих популяциях хромосомных рас Зап. и Центр. Европы (Lukacova et al., 1994; Brunner et al., 2002; Zima et al., 2003; Fredga, 2007), сформировался, вероятно, в результате гибридизации популяций с разным числом метацентриков при восстановлении единого ареала в межледниковье и постледниковый период (Fredga, 2007; Орлов и др., 2008).
В связи с этим интерес представляет выявление особенностей узких и широких гибридных зон S. araneus Вост. Европы. Нами изучены узкие гибридные зоны между тремя мономорфными расами на Валдайской возвышенность (Тверская обл.) (Орлов и др., 2012а, б, 2013), а также полиморфные популяции трех рас и широкая гибридная зона между двумя из них на территории Беларуси (Орлов, Борисов, 2009; Borisov et al., 2010, 2014). В данном сообщении мы представляем результаты сравнительного анализа зон контакта и гибридизации, сформированных между мономорфными расами и между полиморфными популяциями указанных территорий.
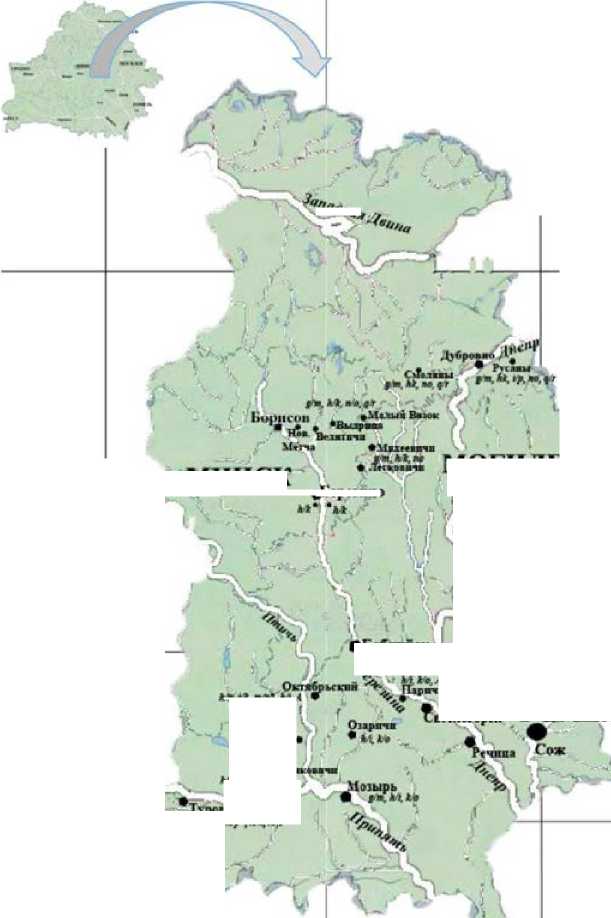
Методика. Проанализированы ранее опубликованные кариологические данные по 140 бурозубкам, отловленным в 2007-2010 гг. на Валдайской возвышенности, где контактируют хромосомные расы Зап. Двина, Москва и Селигер (Борисов и др., 2008, Борисов и др., 2009, Орлов и др., 2012, 2013) (табл. 1), и по 150 бурозубкам из полиморфных популяций с диагностическими хромосомами рас Борисов, Bialowieza и Киев, отловленным в бассейне Днепра и Припяти на территории Беларуси в 2008-2013 гг. (Орлов, Борисов, 2009, Borisov et al., 2010, 2014) (рисунок; табл. 2).
Таблица 1
Кариотипы бурозубок в гибридных зонах хромосомной расы
Зап. Двина с расой Москва и расой Селигер на Валдайской возвышенности (Тверская обл., РФ) (данные из статьи Орлов и др., 2013)
|
Гибридная зона |
и (всего) |
2NA |
Кариотипы (конфигурации хромосом в мейозе) |
|
Западная flBHHa(Wd) -Москва (Мо) |
(37) |
||
|
15 |
18 |
Wd: gm, hk, ip, no, qr |
|
|
9 |
18 |
Fl: gm, hk/kr/rq/qp/pi/ih |
|
|
13 |
18 |
Mo: gm, hi, kr, no, pq |
|
|
Западная Двина (Wd) -Селигер (S1) |
(ЮЗ) |
||
|
12 |
18 |
Wd: gm, hk, ip, no, qr |
|
|
13 |
19-20 |
Fl: 1) g/gm/mq/qr/rp/pi/ik/kh/hn/no (CXI) 2) g/gm/mq/qr/r + p/pi/ik/kh/hn/no/o (СУ+СУП) |
|
|
78 |
19-22 |
SI: g/o, hn, ik, m/q, p/r |
Хромосомные препараты приготовлены по стандартной методике из клеток костного мозга и селезенки. Идентификация хромосом проведена по рисунку G-окраски (обработка препаратов трипсином и 2 х SSC) согласно общепринятой номенклатуре хромосом S. araneus (Searle et al., 1991).
^ТИНСК
^ер^пно
Со*
L-K
27* )0'
31е№0
29-00’
Принять
рж. МИГ
МОГИЛЕВ
*«,*чЧ-»■.*». ^л
•Бибрумк
(«логово;
ГОМЕЛЬ
ss-oo1
54"00
53*00'
52‘00*
Рисунок. Распространение диагностических метацентриков хромосомных рас S. araneus на территории Беларуси в междуречье Днепра и Припяти: А - акроцентрический кариотип; F - гибриды; * - редкие метацентрики
Таблица 2
Пункты отлова бурозубок в междуречье Днепра и Припяти (Беларусь) и их кариотипы
|
№ |
Пункты; с. ш., в. д. |
N (всего) |
2NA |
Кариотипы |
|
1 |
2 |
3 |
4 |
5 |
|
Раса Западная Двина: полиморфные популяции |
||||
|
1 2 |
Русаны (Витебская обл., окр. Дубровно) |
3 |
18-20 |
g/m, hk, i/p, по, q/r |
|
Раса Борисов |
||||
|
2 2 |
Смоляны (Витебская обл.) |
2 |
20-21 |
g/m, hk, i, p, no, q/r |
|
3 3 |
Новая Метча (Минская обл., окр. г. Борисов) |
4 |
23 |
gm, hk, i, p, n/o, q, r |
|
43 |
Велятичи (Минская обл.) |
2 |
22-23 |
gm, hk, i, p, n/o, q/r |
|
53 |
Выдрица (Минская обл.) |
4 |
22-24 |
gm, hk, i, p, n/o, q/r |
|
63 |
Малый Вязок (Минская обл.) |
21 |
22-25 |
g/m, h/k, i, p, n/o, q/r |
|
7 3 |
Михевичи (Минская обл.) |
5 |
25-26 |
g/m, h/k, i, p, no, q, r |
|
8 3 |
Лесковичи (Минская обл.) |
2 |
25-26 |
g/m, hk, i, p, n, o, q, r |
|
9 3 |
Березин о (Минская обл., левый б. Березины) |
(17) |
||
|
13 |
25-27 |
g/m, h/k, i, p, n, o, q, r |
||
|
4 |
28 |
g, m, h, k, i, p, n, o, q, r |
||
|
103 |
Березин о (Минская обл., правый б. Березины) |
(Ю) |
||
|
9 |
26-27 |
g, m, h/k, i, p, n, o, q, r |
||
|
1 |
28 |
g, m, h, k, i, p, n, o, q, r |
||
|
11 1 |
Bobruisk (Могилевская обл.) |
2 |
27 |
g, h/i, k, m, n, o, p, q, r |
|
53° 02', 29° 15' |
||||
|
Полиморфные популяции с диагностическими хромосомами рас Bialowieza и Киев |
||||
|
12 4 |
Октябрьский (Гомельская обл., прав. б. р. Птичь |
(18) |
||
|
52°38', 28°52' |
1 |
24 |
g, h/n, ik, m/p, o, q, r |
|
|
13 |
24-27 |
g, h/n, i/k, m, o, p, q, r |
||
|
3 |
28 |
g, m, h, k, i, p, n, o, q, r |
||
|
1 |
26 |
g, hi, k, m, n, o, p, q, r |
||
|
16 3 |
Озаричи (Гомельская обл., р. Виша) |
(2) |
||
|
52°14', 29°13' |
1 |
26 |
g, hi, k, o, m, n, p, q, r |
|
|
1 |
26 |
g, h/n, i, k/o, m, p, q, r |
||
|
17 3 |
Лучицы (Гомельская обл., прав. б. р. Птичь) |
(5) |
||
|
52°27', 28°43' |
1 |
26 |
g/m, h/i, k, n, o, p, q, r |
|
|
3 |
28 |
g, h, i, k, m, n, o, p, q, r |
||
|
1 |
27 |
g, h/k, i, m, n, o, p, q, r |
||
|
18 3 |
Конковичи (Гомельская обл., лев. б. р. Припять) |
(7) |
||
|
52°09', 28°43' |
2 |
26 |
g, h/n, i/k, m, o, p, q, r |
|
|
4 |
28 |
g, h, i, k, m, n, o, p, q, r |
||
|
1 |
27 |
g, h/k, i, m, n, o, p, q, r |
||
|
19 3 |
Туров (Гомельская обл., прав. б. р. Припять) |
(22) |
||
|
№ |
Пункты; с. ш., в. д. |
N (всего) |
2NA |
Кариотипы |
|
52°04', 27°44' |
1 |
25 |
g/r, h/n, i/k, т, о, р, q, г |
|
|
1 |
25 |
g, h/n, i/k, т/р, о, q, г |
||
|
14 |
24-27 |
g, h/n, i/k, m, о, p, q, r |
||
|
3 |
28 |
g, h, i, k, m, n, o, p, q, r |
||
|
1 |
26 |
g, h, ik/ko, m, p, q, r |
||
|
2 |
27 |
g, h/k, i, m, n, o, p, q, r |
||
|
20 1 |
Туров (Гомельская обл., прав. б. р. Припять) |
(1) |
||
|
52°00', 27°56' |
1 |
27 |
g, h/k, i, m, n, o, p, q, r |
|
|
21 3 |
Мозырь (Гомельская обл., прав. б. р. Припять) |
(2) |
||
|
52°03', 29°15 |
1 |
25 |
g/m, h/i, k/o, n, p, q, r |
|
|
1 |
26 |
g, h/i, k/o, m, n, p, q, r |
Примечание. Гетерозиготность особи и полиморфизм в выборке показаны косой черточкой (/); ссылки: 1 - Mishta et al., 2000; 2 - Орлов, Борисов, 2009; 3 - Borisov et al., 2010; 4 - Borisov et al., 2014.
Результаты и обсуждение. Узкие гибридные зоны хромосомных рас S. araneus Москва, Зап. Двина и Селигер на Валдайской возвышенности. Две изученные хромосомные расы, Москва (диагностические метацентрики gm, hi, kr, по, pq) и Зап. Двина (gm, hk, ip, по, qr), характеризуются мономорфным полностью метацентрическим кариотипом (Орлов и др., 2012). Третья раса, Селигер (hn, ik, m/q, р/r), мономорфна по двум Rb-транслокациям, но полиморфна по транслокациям mq и рг. Кроме того, у этой расы изредка встречается транслокация go (Павлова и др., 2007; Bulatova et al., 2011). Расы Зап- Двина и Москва распространены строго парапатрически; узкая зона контакта и гибридизации между ними не превышает 0,5 км; частота гибридных особей составляет 24% (Орлов и др., 2012). Ширина гибридной зоны рас Зап. Двина и Селигер составляет около 1,5 км, а частота гибридных особей - 30 % (Орлов и др., 2013). В обеих гибридных зонах расы Зап. Двина частота гибридных особей значительно ниже, чем частота, ожидаемая по уравнению Харди-Вайнберга (Орлов и др., 2012, 2013). Гибридная зона рас Москва и Зап. Двина оказалась самой узкой из известных бимодальных зон. Так, ширина гибридной зоны рас Москва и Селигер - 2,5 км (Bulatova et al., 2007), a Drnholec х Bialowieza - 1,7 км (Szalaj et al., 1996).
Ширина гибридной зоны, как правило, коррелирует с кариологической дифференциацией между расами: чем больше у них метацентриков неполной гомологии (и, соответственно, сложнее конфигурации в первом делении мейоза, «цепочки» или «кольца»), тем уже гибридная зона. Например, у гибридов Зап. Двина - Москва в мейозе ожидается кольцо из шести метацентриков (RVI), а гибридов
Зап. Двина - Селигер - цепочка из И хромосом (CXI), или, у гетерозигот по Rb транслокации рг, две цепочки (CV + СУП) (табл. 1). При отсутствии метацентриков неполной гомологии ширина гибридной зоны достигает 100 км (Searle, Wojcik, 1998; Lukacova et al., 1994; Zima et al., 2003; Borisov et al., 2010). Однако, кариотипические различия между контактирующими расами - не единственный фактор, определяющий ширину гибридной зоны. Так, ширина гибридной зоны между расами Томск и Новосибирск (у гибридов-комплексных гетерозигот в мейозе формируются цепочки CIX) составляет 12,8 км (Polyakov et al., 2011).
Как уже было упомянуто, границы хромосомных рас Валдайской возвышенности проходят по границам последней стадии Валдайского оледенения (Борисов и др., 2008). Сформированы эти расы, однако, были еще до последнего оледенения, в процессе многократной смены рефугиальной (фиксация транслокаций в изолятах рефугиумов во время оледенений) и зональной фаз (распространение метацентриков при восстановлении ареала во время межледниковий) (Орлов и др., 2008). В результате многократной изоляции между хромосомными расами могли накопиться не только хромосомные, но и генные различия, которые влияют на экологические и/или этологические особенности расы. В случае рас Зап. Двина и Москва крайняя узость гибридной зоны может быть связана с очень маленькой территорией совместного обитания этих рас, что, в свою очередь, обусловлено внутривидовой конкуренцией рас (Орлов и др., 2013).
Не исключено также, что ширина гибридной зоны между кариологически дифференцированными расами занижена потому, что гибриды-беккроссы цитологически идентичны либо представителям исходной расы, либо гибридам F1, и при анализе хромосом их обнаружить невозможно. Лишь в случаях, когда одна из рас полиморфна по Rb-транслокациям, возможно выявить гибриды от возвратных скрещиваний, например, гибриды рас Селигер и Москва, наследующих акроцентрик g от расы Селигер (Bulatova et al., 2011). Изредка встречающиеся особи расы Москва, гетерозиготные по Rb-транслокации gm (Орлов и др., 2012), могут быть беккроссами, наследующими акроцентрик g от расы Селигер.
При наличии у контактирующих рас метацентриков монобрахиальной гомологии формируются узкие встречно направленные клины частот метацентриков (Banaszek, 1994; Szalaj et al., 1996; Bulatova et al., 2011). Если различия между расами сводятся к тому, что хромосомы, формирующие метацентрик у одной расы, представлены акроцентриками у другой (например, gm и g, т), то формируются однонаправленные клины, ширина которых достигает сотни километров (Searle, Wojcik, 1998; Jones, Searle, 2003; Zima et al., 2003; Brunner et al., 2002; Borisov et al., 2010).
Клиналъная изменчивость частоты метацентриков хромосомных рас Зап. Двина, Борисов, Bialowieza и Киев в междуречье Днепра и Припяти и гибридная зона Bialowieza - Киев. Хромосомная раса Зап. Двина мономорфна практически на всем своем ареале (протяженность около 300 км) от Валдайской до Смоленской возвышенности. На Оршанской возвышенности обнаружены особи расы Зап. Двина, гетерозиготные по Rb-транслокациям ip и qr, а южнее окр. гг. Смоляны и Борисов хромосомы i и р встречаются только в акроцентрическом состоянии (Орлов, Борисов, 2009). Популяции, гомозиготные по акроцентрическим хромосомам i и р и полиморфные по транслокациям gm, hk и по были выделены в хромосомную расу Борисов (Орлов, Борисов, 2009). Частота метацентриков gm, hk и по постепенно снижается на протяжении 100 км. На южной границе расы Борисов преобладают гетерозиготы по единственной транслокации hk, а также встречаются особи, в кариотипе которых все диагностические хромосомы представлены акроцентриками (Borisov et al.,2010).
Было высказано предположение (Орлов, Борисов, 2009; Borisov et al.,2010), что ранее на территории междуречья Днепра и Припяти были распространены популяции с 10 парами диагностических акроцентриков, т.е. с кариотипом, аналогичным кариотипу рас Cordon и Pelister (Wojcik et al., 2002). В послеледниковье, при расселении из рефугиумов, популяции расы Зап. Двина вступили в контакт с популяциями, характеризующимися акроцентрическим кариотипом. В результате распространения метацентриков расы Зап. Двина по ареалу популяций с акроцентрическим кариотипом была сформирована полиморфная раса Борисов (Орлов, Борисов, 2009; Borisov et al., 2010). Особого внимания заслуживает тот факт, что частота простых гетерозигот по Rb-транслокации по в популяциях расы Борисов не отличается от частоты, ожидаемой по уравнению Харди-Вайнберга, в отличие от пониженной частоты комплексных гетерозигот в гибридных зонах между расой Зап. Двина и расами Москва и Селигер (Орлов и др., 2013). Соответствие частоты Rb-гетерозигот теоретически ожидаемой свидетельствует о том, что гибридизация между расами без метацентриков неполной гомологии (когда среди гибридов только простые гетерозиготы по Rb-транслокациям) не ограничена.
При дальнейших кариологических исследованиях 5. araneus на территории Беларуси было обнаружено, что популяции двух хромосомных рас Bialowieza и Киев, характеризуются большим полиморфизмом по сравнению с популяциями, обитающими на основной части их ареалов в Польше и на Украине (Borisov et al., 2014). От территории восточной Польши (Wojcik et al., 1996) до р. Птичь (Borisov et al., 2014) распространены бурозубки с диагностическими хромосомами расы Bialowieza (gr, hn, ik, т/р). Диагностические метацентрикиы gr и тр были зарегистрированы лишь в западных -210- пунктах; в 2 популяциях на левом берегу р. Птичь их частота не превышала 2 %; еще в двух пунктах наблюдались только метацентрики hn и ik, частота которых варьировала от 36 до 67 % и от 34 до 73 %, соответственно. Следует также отметить, что в четырех пунктах наблюдались кариотипы, в которых все диагностические хромосомы были представлены акроцентриками (Borisov et al., 2014) (табл. 2; рис. 1). '
Бурозубки с двумя диагностическими метацентриками расы Киев, hi и k/о, наблюдались восточнее р. Птичь на правом и левом берегу р. Березина, а на правом берегу р. Припять, в окр. г. Мозырь, были отмечены все три метацентрика этой расы, g/m, hi, к/о (табл. 2).
Популяции расы Bialowieza на основной части ее ареала в восточной Польше мономорфны по Rb-транслокациям gr, hn и ik (лишь в отдельных популяциях выявлен полиморфизм по транслокации тр, а акроцентрический вариант хромосом g и г крайне редок) (Wojcik et al., 1996). Клинальная изменчивость прослеживается от окр. гг. Гродно и Ганцевичи, где отловлены гетерозиготы по Rb-транслокациям, до правобережья р. Птичь, где метацентрики практически исчезают (Borisov et al., 2014). Раса Киев мономорфна по транслокации hi (Mishta et al., 2000); гетерозиготы обнаружены только на территории Беларуси (Borisov et al., 2014).
Клинальная изменчивость в популяциях рас Борисов, Bialowieza и, возможно, расы Киев, свидетельствуют в пользу гипотезы о существовании в прошлом на данной территории акроцентрической расы. Дополнительным аргументом являются также встречающиеся особи с 10 парами диагностических акроцентриков и особи с единственным Rb-метацентриком hi и hk, рассматриваемые ранее как представители расы Бобруйск и расы Туров (Mishta et al., 2000). Следует подчеркнуть, что особи с акроцентрическим кариотипом, считающимся исходным в эволюции 5. araneus (Wojcik et al., 2002), не были выявлены в западноевропейских популяциях с низкой частотой диагностических метацентриков (Lukacova et al., 1994; Brunner et al., 2002; Zima et al., 2003).
В окр. г. Октибрьский, где распространены бурозубки с диагностическими хромосомами расы Bialowieza), была обнаружена особь с диагностическим метацентриком hi расы Киев (Borisov et al., 2014). У одной из бурозубок, отловленной на правобережье р. Птичь (пос. Конковичи) (табл. 2, рис. 1), в кариотипе присутствовали два метаценрика расы Киев, gm и hi. В окр. г. Туров был зарегистрирован гибрид-комплексная гетерозигота с метацентриками обеих рас: ik/ko (табл. 2). По-видимому, зона контакта между популяциями с диагностическими хромосомами рас Киев и Bialowieza проходит приблизительно вдоль русла р. Птичь на протяжении 100 км (табл. 2,
№№ 12, 17, 18) и заходит на правый берег р. Припять (окр. г. Туров) (табл. 2, № 19, рис. 1).
На первый взгляд, низкая частота метацентриков и, соответственно, высокая частота акроцентриков вблизи зоны контакта рас Bialowieza и Киев напоминает явление «акроцентрического пика», т.е. повышение частоты простых гетерозигот с акроцентрическим вариантом полиморфных хромосом вследствие отбора против комплексных гетерозигот (Fedyk, 1985; Searle, 1986; Banaszek, 1994). Однако, «акроцентрический пик» наблюдается в гибридных зонах с выраженными встречно направленными клинами метацентриков двух рас. Однонаправленные клины частот метацентриков расы Борисов в бассейне р. Березина и расы Bialowieza с запада на восток до р. Птичь свидетельствуют о распространении диагностических метацентриков в популяциях с акроцентрическим кариотипом. Частоты гомо- и гетерозигот по транслокациям hn и ik в выборках с правобережья р. Птичь и из окр. г. Туров не отличаются от частот, ожидаемых согласно уравнению Харди-Вайнберга, следовательно, высокая частота акроцентриков в этих популяциях не связана с акроцентрическим пиком (Borisov et al., 2014).
Интересно, что в полиморфных популяциях расы Bialowieza в зоне контакта с расой Киев (Табл. 2, №№ 17-20; Borisov et al., 2014; Mishta et al., 2000) были выявлены кариотипы с одним диагностическим метацентриком hk в гетерозиготном состоянии, описанные как хромосомная раса Туров (Mishta et al., 2000). По-видимому, метацентрик hk, диагностический для рас Лепель, Зап. Двина и Борисов, распространенных на 100 км севернее (Bulatova et al., 2000; Borisov et al., 2010), смог проникнуть в эти полиморфные популяции из-за высокой частоты акроцентриков на этой территории.
Внимания заслуживает факт, что разные диагностические метацентрики распространились в популяциях с акроцентрическим кариотипом на неодинаковое расстояние. Так, в клине метацентрических хромосом расы Борисов метацентрик hk проникает на юг дальше, чем метацентрики по и qr, а в клине метацентриков расы Bialowieza - hn и ik - дальше на восток, чем gr и тр. Плечи метацентрических хромосом и гомологичные им акроцентрические хромосомы, идентичные по дифференциальной G-окраске, могут отличаться особенностями генного комплекса. Вероятно, в процессе независимой эволюции популяций (в изолятах) у них формируются устойчивые генные комплексы, сцепленные с метацентрическим или акроцентрическим вариантом хромосомы. Эти генные комплексы могут оказаться селективно нейтральными или обладать конкурентным преимуществом. Конкурентные преимущества Робертсоновских транслокаций хромосом могут зависеть от условий окружающей среды, в том числе биотопических. Например, было обнаружено, что в лесном -212- “ массиве Беловежская пуща частота метацентрика тр хромосомной расы Bialowieza несколько отличалась в популяциях, населяющих сухие и влажные биотопы (Wojcik et al., 1996). По мнению авторов, селективное преимущество генного комплекса метацентрика тр может зависеть от условий окружающей среды. Не исключено, что разное распространение диагностических метацентриков рас Борисов и Bialowieza определяется экологическими факторами.
Принимая во внимание неодинаковое распространение метацентриков одной расы, разнообразие метацентриков, унаследованных от различных рас, мигрировавших в междуречье Днепра и Припяти из разных рефугиумов в послеледниковье, а также отбор против комплексных гетерозигот, ослабленный вследствие низкой частоты метацентриков, можно ожидать, что гибридные зоны полиморфных популяций будут характеризоваться очень сложной структурой.
Выводы. Крайне узкие (от 2,5 до 0,5 км) гибридные зоны, сформированные между мономорфными хромосомными расами S. araneus, Зап. Двина, Москва и Селигер, различающимися между собой по 3-4 Робертсоновским (Rb) транслокациям, на Валдайской возвышенности, по-видимому, обусловлены ограниченной областью совместного обитания этих рас, что, вероятно, связано с конкурентными отношениями между этими кариотипически дифференцированными расами. Низкая частота гибридов-комплексных гетерозигот в узких гибридных зонах рас Зап. Двина, Москва и Селигер, по сравнению с частотой, ожидаемой согласно уравнению Харди-Вайнберга, можно объяснить такими факторами, как пониженная приспособленность и плодовитость комплексных гетерозигот со сложными конфигурациями (кольцами и цепочками) в мейозе и ассортативное скрещивание в зоне контакта. Полиморфные популяции рас Зап. Двина, Борисов, Bialowieza и Киев сформировались в результате гибридизации с акроцентрической расой, существующей на этой территории в прошлом. Как известно, при гибридизации метацентрических и акроцентрических популяций образуются только простые гетерозиготы и диагностические метацентрики разных хромосомных рас могут свободно распространяться в популяциях с акроцентрическими кариотипами на большие расстояния. Однонаправленные клины частоты диагностических метацентриков рас Борисов и Bialowieza, а также соответствие частот гомо- и гетерозигот по Робертсоновским транслокациям S. araneus уравнению Харди-Вайнберга в полиморфных популяциях этих рас свидетельствуют, что низкая частота акроцентриков в области контакта полиморфных популяций с диагностическим хромосомами Bialowieza и Киев являются скорее результатом распространения метацентриков по ареалу популяций с акроцентрическим кариотипом, чем «акроцентрического пика».
Авторы благодарны доц. каф. биологии Тверского госуниверситета к. б. н. А.А. Емельяновой, а также студентам-дипломникам: Тверского госуниверситета - О.Е. Егоровой и А.Е. Пентюхову, Витебского университета - АМ. Спрингер, Кубанского университета - А.А. Ковалевой и Мозырского университета - Е.М. Бочко и Д.Ю. Лесничему за помощь на всех этапах работы.
Narrow hybrid zones of three monomorphic chromosomal races of the common shrew, Sorex araneus, differing from each other by 3 - 4 diagnostic Robertsonian translocations, were revealed in the north of Tver Region. Localization of these hybrid zones follows marginal formation of the Veps stage glacier. A small width of hybrid zones (0,5 - 2,5 km) is determined by a small area of co-habitation of these karyotypically diverged races, which is caused , probably, by concurrent relations between them. Diagnostic metacentrics of the West Dvina race, which is monomorphic from the Valdai to the Smolensk Heights, were noted in populations of the Borisov race on the territory of Belarus (specific feature of this race is the absence of metacentric ip trait for the West Dvina race). A cline variability of the diagnostic metacentric frequencies, decreasing towards the Dnieper and Pripyat junction, was noted in populations of the Borisov and Bialowieza races on the territory of Belarus. A high acrocentric frequency near the hybrid zone between populations with diagnostic metacentrics of the Bialowieza race and the Kiev race was noted. The specific features of the polymorphic populations at the Dnieper and Pripyat junction can be explained by the existence in the past on the mentioned area the common shrew populations with completely acrocentric karyotype. Karyotype with all the diagnostic chromosomes represented by acrocentrics, regarded as primary in the evolution of S. araneus, was revealed in a number of polymorphic populations in the discussed region.
КРИЩУК Ирина Александровна - аспирант, ГНПО «Научнопрактический центр Национальной академии наук по биоресурсам», 220072, Республика Беларусь, Минск, ул. Академическая, д. 27, e-mail: ikry shchuk@yandex .by
БОРИСОВЕЦ, Ирина Павловна - студентка, У О Мозырский государственный педагогический университет им. И.П. Шамякина, 247760, Республика Беларусь, Мозырь, ул. Студенческая, 28
Черепанова Е.В. Особенности гибридных зон хромосомных рас обыкновенной, бурозубки, Sorex araneus L., с разным уровнем кариотипической дифференциации / Е.В. Черепанова, Е.С. Гайдученко, И.А. Крищук, И.П. Борисовец, Ю.М. Борисов// Вести. ТвГУ. Сер. Биология и экология. 2014. № 4. С. 202-218.
Список литературы Особенности гибридных зон хромосомных рас обыкновенной бурозубки, Sorex araneus L., с разным уровнем кариотипической дифференциации
- Борисов Ю.М., Козловский А.И., Балакирев А.Е., Демидова Т.Б., Ирхин С.Ю. и др. 2008. Контакты хромосомных рас обыкновенной бурозубки, Sorex araneus L. (Insectivora) на краевых образованиях вепсовской стадии Валдайского ледника//Сибирский экологический журн. № 5. С. 763-771.
- Бородин П.М., Поляков А.В. 2008. Гены, хромосомы и видообразование//Современные проблемы биологической эволюции. М.: Изд-во ГДМ. С. 136-148.
- Орлов В.Н., Борисов Ю.М. 2009. Филогенетические связи популяций обыкновенной бурозубки (Sorex araneus, Insectivora) Белоруссии по кариологическим данным//Зоол. журн. Т. 88. № 12. С. 1506-1514.
- Орлов В.Н., Борисов Ю.М., Черепанова Е.В., Григорьева О.О., Шестак А.Г., Сычева В.Б. 2012. Узкая гибридная зона хромосомных рас Москва и Западная Двина и особенности изоляции популяций обыкновенной бурозубки, Sorex araneus (Mammalia)//Генетика. Т. 48. № 1. С. 80-88.
- Орлов В.Н., Борисов Ю.М., Черепанова Е.В., Милишников А.Н. 2013. Ассортативное скрещивание в гибридных зонах хромосомной расы Западная Двина обыкновенной бурозубки, Sorex araneus (Mammalia)//Докл. АН. Т. 451. №. 1. С. 110-113.
- Орлов В.Н., Козловский А.И., Балакирев А.Е., Борисов Ю.М. 2008. Процессы фиксации метацентрических хромосом в популяциях обыкновенной бурорзубки, Sorex araneus L., Восточной Европы//Генетика. Т. 44. № 5. С. 581-593.
- Орлов В.Н., Козловский А.И., Балакирев А.Е., Борисов Ю.М. 2007. Эндемизм хромосомных рас обыкновенной бурозубки Sorex araneus L. и возможность сохранения рефугиумов в области покровного оледенения Поздневалдайской эпохи//Докл. АН. Т. 416. № 6. С. 1-4.
- Павлова С.В., Булатова Н.Ш., Щипанов А.Н. 2007. Цитогенетический контроль гибридной зоны двух хромосомных рас Sorex araneus перед сезоном размножения//Генетика. Т. 43. № 11. С. 1619-1626.
- Щипанов Н.А., Булатова Н.Ш., Павлова С.В., Щипанов А.Н. 2009. Обыкновенная бурозубка (Sorex araneus) -модельный вид экологоэволюционных исследований//Зоол. журн. Т. 88. № 6. С. 975-989.
- Banaszek A. 1994. The structure of the contact zone between the chromosomal races Druzno and Legucki Mlyn in the common shrew (Sorex araneus) in north-east Poland: Preliminary results//Folia Zool. 1994. V. 43. Suppl. 1. P. 11-19.
- Borisov Yu.M., Cherepanova E.V., Orlov V.N. 2010. A wide hybrid zone of chromosome races of the common shrew, Sorex araneus Linnaeus, 1758 (Mammalia), between the Dnieper and Berezina Rivers (Belarus)//Comparative Cytogenetics. V. 3. No. 2. P. 195-201.
- Borisov Yu.M., Kryshchuk I.A., Cherepanova E.V., Gaiduchenko E.S., Orlov V.N. 2014. Chromosomal polymorphism of populations of the common shrew, Sorex araneus L., in Belarus//Acta Theriol. V. 59. Suppl. 2. P. 243-249.
- Brunner H., Turni H., Kapischke H-J., Strubbe M., Vogel P. 2002. New Sorex araneus karyotypes from Germany and the postglacial recolonization of Central Europe//Acta Theriol. V. 47. No. 3. P. 277-293.
- Bulatova N., Searle J.B., Bystrakova N., Nadjafova R., Shchipanov N., Orlov V. 2000. The diversity of chromosome races in Sorex araneus from European Russia//Acta Theriol 45. Suppl. 1. P. 33-46.
- Bulatova N., Shchipanov N., Searle J.B. 2007. The Seliger -Moscow hybrid zone between chromosome races of common shrews -an initial description//Russian J. Theriol. V. 6. No. 1. P. 111-116.
- Bulatova N.S., Jones R.M., White T.A., Shchipanov N.A., Pavlova S.V., Searle J.B. 2011. Natural hybridization between extremely divergent chromosomal races of the common shrew (Sorex araneus, Soricidae, Soricomorpha): hybrid zone in European Russia//J Evolution. Biol. V. 24. P. 573-586.
- Fedyk S. 1985. Genetic differentiation of Polish populations of Sorex araneus L. II. Possibilities of genetic flow between chromosomal races//Bul. Pol. Acad. Sci. Biol. Sci. V. 34. Nos. 7-9. P. 161-171.
- Fredga K. 2007. Reconstruction of the postglacial colonization of Sorex araneus into Northern Scandinavia based on karyotype studies, and the subdivision of the Abisco race into three//Russ. J. Theriol. V. 6. No. 1. P. 85-96.
- Hausser J., Fedyk S., Fredga K. et al. 1994. Definition and nomenclature of the chromosome races of Sorex araneus//Folia Zool. V. 43 Suppl. 1. P. 1-9.
- Jadwiszczak K.A., Banaszek A. Fertility in the male common shrews, Sorex araneus, from the extremely narrow hybrid zone between chromosome races//Mamm. Biol. 2006. V. 71. P. 257-267.
- Jones R.M., Searle J.B. 2003. Mapping the course of the Oxford-Hermitage chromosomal hybrid zone in the common shrew Sorex araneus -a GIS approach//Mammalia. V. 68. Р. 193-200.
- Lukacova L., Pialec J., Zima J. 1994. A hybrid zones between the Ulm and Drnholec karyotypic races of Sorex araneus in Czech Republic//Folia Zool. V. 43. Suppl. 1. P. 37-42.
- Mishta A.V., Searle J.B., Wojcik J.M. 2000. Karyotypic variation of the common shrew Sorex araneus in Belarus, Estonia, Latvia, Lithuania and Ukraine//Acta Theriol. V. 45. Suppl. 1. P. 47-58.
- Orlov V.N., Borisov Yu.M. 2007. Chromosome races of the common shrew Sorex araneus L. (Mammalia: Insectivora) from the south part of Valdai Heights//Comparative Cytogenetics. V. 1. No. 2. P. 101-106.
- Orlov V.N., Kozlovsky A.I., Okulova N.M., Balakirev A.E. 2007. Postglacial recolonisation of European Russia by the common shrew Sorex araneus//Russian J. Theriology. V.6. No. 1. P. 97-104.
- Polyakov A.V., White Th., Jones R. et al. 2011. Natural hybridization between extremely divergent chromosomal races of the common shrew (Sorex araneus, Soricidae, Soricomorpha): hybrid zone in Siberia//J. Evol. Biol. V. 24. P. 1393-1402.
- Searle J.B. 1986. Factors responsible for a karyotypic polymorphism in the common shrew, Sorex araneus//Proc. Royal Soc. London. B. V. 229. P. 277-298.
- Searle J.B., Fedyk S., Fredga K., Hausser J., Volobouev V.T. 1991. Nomenclature for the chromosomes of the common shrew (Sorex araneus)//Mem. Soc. Vaud. Sci. Nat. V. 19. P. 13-22.
- Searle J.B., Wojcik J.M. 1998. Chromosomal evolution: The case of Sorex araneus/Eds J.M. Wojcik, M. Wolsan. Evolution of Shrews. Bialowieza: Mammal Research Institute, Polish Academy of Sciences. P. 219-268.
- Szalaj K.A., Fedyk S., Banaszek A., Chqtnicki W., Ratkiewicz M. 1996. A hybrid zone between two chromosome races of the common shrew, Sorex araneus, in eastern Poland. Preliminary results//Hereditas. V. 125. P. 169-176.
- Wojcik J.M., Borodin P.M., Fedyk S. et al. 2003. The list of the chromosome races of the common shrew Sorex araneus (updated 2002)//Mammalia. V. 67. P. 169-178.
- Wojcik J.M., RatkiewiczM., Searle J.B. 2002. Evolution of the common shrew Sorex araneus: chromosomal and molecular aspects. In: Gliwicz J. (ed) Theriology at the Turn of a New Century//Acta Theriol. V. 47. Suppl. 1. P. 139-167.
- Wojcik J.M., Wojcik A.M., Zalewska H. 1996. Chromosome and allozyme variation of the common shrew, Sorex araneus, in different habitats/Fredga K, Searle JB (eds)//Evolution in the Sorex araneus Group. Cytogenetic and Molecular aspects. Hereditas Offprint. V. 125. P. 183-189.
- Zima J., Slivkova L., Tomaskova L. 2003. New data on karyotypic variation in the common shrew, Sorex araneus, from the Czech Republic: an extension of the range of the Laska race//Mammalia. V. 68. No. 2. P. 209-215.
