Особенности горизонтальной структуры внутрилесных лугов Неруссо-Деснянского полесья

Автор: Горнов Алексей Владимирович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Структура и динамика растительных сообществ
Статья в выпуске: 1-5 т.14, 2012 года.
Бесплатный доступ
В статье рассмотрены особенности горизонтальной структуры внутрилесных лугов Неруссо-Деснянского полесья. Выявлены масштабы роющей деятельности кабанов на регулярно косимых лугах и некосимых лугах, заросших высокотравьем. Установлена площадь и структура фитогенной мозаичности, формируемая кустарниками. Показано, что наиболее крупные мозаики на лугах создаются деятельностью человека.
Внутрилесные луга, горизонтальная структура, мозаичность
Короткий адрес: https://sciup.org/148200790
IDR: 148200790 | УДК: 574.47
The specific properties of the horizontal structure of the forest meadows in the Nerussa-Desna Polesye
The specific properties of the horizontal structure of the forest meadows in the Nerussa-Desna Polesye are described in the article. The extent of the wild boar digging activity on the regularly moving meadows and not moving meadows, overgrown with tall grass, was revealed. The size and the structure of the phytogenic patchiness, formed by shrubs, were determined. It was shown, that the largest mosaics in the meadows are created by the human activity.
Текст научной статьи Особенности горизонтальной структуры внутрилесных лугов Неруссо-Деснянского полесья
Современные луга лесного пояса возникли благодаря хозяйственной деятельности человека: сенокошения, выпаса, распашки, намеренных палов и др. [12, 11, 2 и др.]. С появлением внутрилесных лугов сформировались природные механизмы поддержания их биологического разнообразия [15, 6, 19 и др.]. Для понимания таких механизмов в первую очередь необходимо изучать структуру сообществ. В связи с этим в работе поставлена цель – описать особенности формирования горизонтальной структуры внутрилесных лугов.
Материал собран на влажных внутрилесных лугах Неруссо-Деснянского полесья (НДП). Это юго-восточная часть Брянской области. В ботанико-географическом плане район принадлежит Полесской подпровинции Восточноевропейской широколиственной провинции [13]. Исследуемые луга в основном подвержены ручному сенокошению, которое сохраняет большинство природных мозаик.
В ходе исследований описаны особенности формирования горизонтальной неоднородности лугов и проведено картирование их мелко- и крупноконтурной мозаичности. Картирование выполнялось во второй половине лета. При выборе мест картирования исходили из того, что луг представляет собой систему разновозрастных и разноразмерных вложенных Элементов мозаики (микросайты) разного происхождения. Картирование мелкоконтурной мозаики выполняли с использованием веревок. Участок луга размечали веревками на квадраты по 1 кв. м. Далее на карту-схему наносились контуры элементов мозаики в масштабе 1 : 25. Для всех мозаик отмечены размеры, происхождение и, по возможности, давность возникновения, которая определялась по возрасту деревьев и кустарников, внедрившихся на элемент мозаики. Картирование крупноконтурной мозаики проводили при помощи GPS-навигатора. При этом закартированы три луга общей площадью 2,15 га. При картировании по контуру обходились крупные элементы мозаики луга (кабаньи порои и скопления кустарников) с включенным прибором GPS. Во время об-
обхода координаты местоположения регистрировались в памяти прибора через каждые два метра. Далее координаты всех точек обрабатывались в пакете программы MapInfo Professional, где по ним строились соответствующие карты-схемы.

Рис. 1. Фрагмент горизонтальной структуры регулярно косимого внутрилесного луга. Площадь – 54 кв. м. Порои кабанов: 1 – свежие, 2 – возрастом 1-2 месяца, 3 – возрастом до одного года, 4 – возрастом один и более лет; муравейники: 5 – Lasius niger , 6 – L. flavus ; мелкие выбросы и постройки: 7 – кротов и мышевидных грызунов, 8 – различных луговых муравьев.
На внутрилесных лугах НДП наиболее четко выражены зоогенные, фитогенные и антропогенные мозаики1. Рассмотрим их подробнее.
Зоогенная мозаика формируется в ходе деятельности животных, в основном роющих. Их порой, выбросы, поверхностные ходы и постройки создают горизонтальную неоднородность луговых фитоценозов (рис. 1). Наиболее крупные элементы зоогенной мозаики формируют кабаны ( Sus scrofa ). При этом они охотнее перекапывают регулярно косимые луга: здесь плотность пороев в 20 раз выше, чем на угодьях, которые не косят (рис. 2). Видимо, это связано с их кормностью. Мозаики меньших размеров (табл. 1) появляются благодаря кротам ( Talpa europaea ), мышевидным грызунам ( Muridae ) , муравьям ( Lasius flavus , L. niger и др.). Перечисленные животные также активнее на косимых лугах. Все зоогенные микросайты характери-
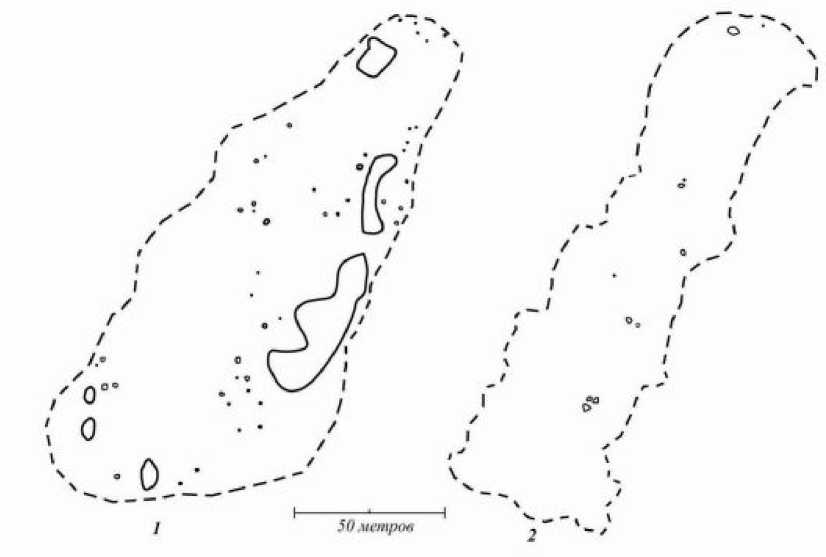
Рис. 2 . Расположение кабаньих пороев на внутрилесных лугах. 1 - регулярно косимый луг, 2 - некосимый луг, заросший высокотравьем. Черным пунктиром показаны границы лугов. Черными кружками обозначены порои кабанов площадью менее 0,5 кв. м, а контурами - более 0,5 кв. м
Таблица 1 Размеры некоторых элементов мозаик, встречающихся на лугах
|
Элементы мозаики |
Площадь, кв. м |
Зоогенные
|
Порои Sus scrofa |
0,01-20,00 |
|
Муравейники Lasius niger |
0,03-2,00 |
|
Выбросы Talpa europaea |
0,10-0,36 |
|
Выбросы Arvicola terrestris |
0,08-0,30 |
Фитогенные
|
Заросли кустарников |
1,00-200,00 |
|
Заросли Filipendula ulmaria |
1,00-10000,00 и более |
|
Кочки Carex appropinquata |
0,03-0,35 |
Антропогенные
|
Осушенные участки лугов |
10000000,0 и более |
|
Косимые участки лугов |
10000,00 и более |
|
Стоговины |
1,00-10,00 |
Фитогенная мозаика определяется вегетативным разрастанием особей растений. Например, при зуются особыми экологическими условиями. Например, пороям кабанов свойственны обнаженный субстрат, повышенная аэрация, влажность, температура и значительная микробиологическая активность [5, 7, 21 и др.]. Муравьи при строительстве жилищ перемешивают и взрыхляют почву. Разрыхленная почва муравейников также отличается повышенной температурой и улучшенным газообменом. Это усиливает интенсивность микробиологических процессов и увеличивает количество доступных элементов питания [8, 3, 18]. Особые экологические условия зоогенных микросайтов определяют их флористический и фаунистический состав, который отличается от окружающих участков луга [4].
отсутствии сенокошения на лугах могут разрастаться кустарники (Salix cinerea, S. rosmarinifolia, S. starkeana) и подрост деревьев (Betula pubescens, Pinus sylvestris, Salix pentandra). Это происходит в том случае, если кабаны перерывают луга и создают нарушения, необходимые для приживания молодого поколения древесных растений. При этом кустарники и деревья располагаются отдельными плотными скоплениями, размещение которых определяется прошлыми кабаньими пороями (рис. 3). Разросшиеся кустарники и подрост деревьев значительно влияют на окружающую территорию. Например, почва под древесными растениями обогащается азотом за счет опада [17]. Крупные элементы мозаики образуют монодоминантные заросли высокотравья: Filipendula ulmaria и Carex acuta. Они возникают на лугах при отсутствии сенокошения и роющей деятельности животных. Маршрутные наблюдения показали, что размеры таких мо- заик в настоящее время бывают разные (табл. 1). Другие виды растений (например, кочкообразующие осоки Carex appropinquata и C. сespitosa) формируют мозаики меньшего размера. По площади они аналогичны нарушениям мелких животных. Биологический смысл формирования кочки – вынос почек возобновления выше уровня затопления во время половодья. В связи с этим кочки представляют собой благоприятный субстрат для приживания растений, которые формируют здесь особые микрогруппировки [14, 20].
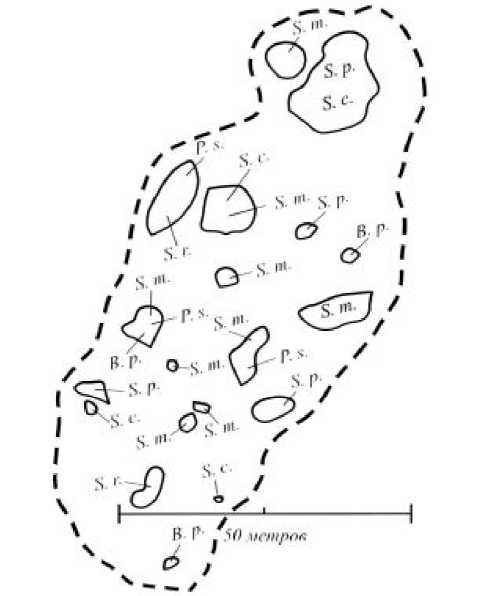
Рис. 3 . Расположение скоплений кустарников и подроста деревьев на внутрилесном лугу. Сенокошение отсутствует 4-10 лет. Черным пунктиром показана граница луга, а контурами - скопления кустарников и подроста деревьев: B. p . – Betula pubescens , P. s. – Pinus sylvestris, S. c. – Salix cinerea, S. m. – S. myrsinifolia, S. p. – S. pentandra, S. r. – S. rosmarinifolia
Антропогенная мозаика. В результате хозяйственной деятельности человека возникает два варианта мозаичности лугов: небольшие и крупные элементы мозаики. Небольшие элементы мозаики (стоговины, навозные кучи, сбоины от копыт и др.) чаще появляются при традиционном природополь-зовнаии. По экологической роли они сопоставимы с природной неоднородностью растительного покрова. Крупные элементы мозаики возникают в ходе промышленного использования угодий, при котором определяющее значение принадлежит урожайности и качеству сена [1, 16 и др.]. Если они не удовлетворяют имеющиеся нужды, то обычно проводят коренное улучшение лугов. Под коренным улучшением понимают систему мероприятий, направленных на повышение урожайности сельскохозяйственных угодий. К ним относят: осушение заболоченных лугов и пастбищ, очистку от кустарников и подроста деревьев, поверхностное рыхление (фрезерование и боронование), распашку, внесение удобрений [8, 9 и др.]. Перечисленные мероприятия значительно изменили структуру и экологические условия практически всех луговых сообществ. Например, фрезерование и боронование уничтожают осоковые кочки, муравейники, кротовины и другую горизонтальную неоднородность сообществ. При этом на всей их площади луга выравниваются многие почвенные характеристики: кислотность, содержание доступного азота, обменных кальция и магния [16]. При фрезеровании лугов изменяется водный режим почвы: сначала влажность корнеобитаемого слоя может увеличиться на 10-15%, но потом значительно сократиться [1]. Распашка лугов также уничтожает зоогенный микрорельеф, обнажает поверхность почвы, развивает ветровую и водную эрозию, дезагрегирует почвенный материал. Все это ведет к минерализации гумуса и поверхностной деградации почвы.
Заключение. Внутрилесные луга НДП состоят из элементов мозаичной структуры разного происхождения, размера и возраста. В ходе исследований выделены природные (фитогенные и зоогенные) и антропогенные элементы мозаики. Хозяйственная деятельность создает самые крупные элементы мозаики. Элементы мозаики меньших размеров формируют растения и животные. Зоогенные элементы мозаики обычно формируются роющими животными. Фитогенные элементы мозаики, как правило, образуются вегетативно разрастающимися видами растений. В результате деятельности человека природная мелкомасштабная мозаика может сменяться на чрезвычайно крупную антропогенную мозаику, которая сглаживает неоднородность экологических условий на лугах.
Работа выполнена при поддержке гранта РФФИ № 12-04-01448
Список литературы Особенности горизонтальной структуры внутрилесных лугов Неруссо-Деснянского полесья
- Вагина Т.А. Луга Барабы. Новосибирск, 1962. 196 с.
- Восточноевропейские леса: история в голоцене и современность. Кн. 1. М., 2004. 479 с.
- Голиченков М.В., Новосёлов А.Л., Марфенина О.Е., Добровольская Т.Г., Закалюкина Ю.В., Лапыгина Е.В., Замолодчиков Д.Г. Микробиологическая характеристика муравейников Lasius niger//Известия РАН. Серия биологическая. 2011. № 3. С. 334-339.
- Горнов А.В. Фитогенная и зоогенная мозаичность и флористическое разнообразие влажных лугов Нерусссо-Деснянского Полесья//Бюл. МОИП. Отд. биол. 2011. Т. 116. С. 64-69.
- Гусев А.А. Функциональная роль диких копытных животных в заповедных биогеоценозах//Роль крупных хищников и копытных в биоценозах заповедников. М., 1986. С. 94-105.
- Дымина Г.Д. Луга Дальнего Востока (Зейско-Буреинское Приамурье). Новосибирск, 1985. 193 с.
- Завьялова Л.Ф. Биогеоценотичекая роль кабана в Дарвинском заповеднике и его значение в соседних сельхозугодьях//Научные исследования в заповедниках и национальных парках России (федеральный отчет за 1992-1993 годы). М., 1997. С. 99-100.
- Зрянин В.А. Особенности луговых мирмекокомплексов на почвах с разным механическим составом//Зоологический журнал. 1998. Т. 77. № 7. С. 800-808.
- Качественная оценка, улучшение и рациональное использование природных кормовых угодий Калужской области (рекомендации). Калуга, 1970. 123 с.
- Ковалев Н. Улучшение лугов//В помощь крестьянину. Серия III «Техника и мелиорация». Вып. IV. Курск, 1927. 31 c.
- Оценка и сохранение биоразнообразия лесного покрова в заповедниках Европейской России. М., 2000. 196 с.
- Работнов Т.А. Луговедение. М., 1974. 384 с.
- Растительность европейской части СССР. Л., 1980. 431 с.
- Сарычева Е.П. Микросукцессионные процессы в растительном покрове микроместообитаний черноольховых лесов заповедника «Брянский лес»//Труды международной конференции по фитоценологии и систематике высших растений, посвященной 100-летию со дня рождения А.А. Уранова. М., 2001. С. 159-151.
- Ярошенко П.Д. К изучению горизонтального расчленения растительного покрова//Бот. журн. 1958. Т. 43. № 3. С. 381-387.
- Ярошенко П.Д. Сенокосы и пастбища Приморского края. М.-Л., 1962. 183 с.
- Ястребов А.Б. Влияние ив на травянистый покров луга//Бот. журн. 1989. Т. 74. № 4. С. 476-486.
- Dauber J., Wolters V. Microbial activity and functional diversity in the mounds of three different ant species//Soil Biology & Biochemistry. 2000. Vol. 32. P. 93-99.
- Hopkins B. Pattern in the plant community//Journal of Ecology. 1957. Vol. 45. P. 451-463.
- Werner J.K., Zedler B.J. How sedge meadow soils, microtopography and vegetation respond to sedimentation//Wetlands. 2002. Vol. 22. № 3. P. 451-466.
- Wirthner S. The role of wild boar (Sus scrofa L.) rooting in forest ecosystems in Switzerland. A dissertation for the degree of doctor in sciens. Zurich, 2011. 103 p.
