Особенности экологии гнездования куликов в Байкальской Сибири

Автор: Бадмаева Евгения Николаевна
Журнал: Природа Внутренней Азии @nature-inner-asia
Рубрика: Байкальский регион, Россия
Статья в выпуске: 3 (4), 2017 года.
Бесплатный доступ
Видовой состав гнездящихся куликов в Байкальской Сибири составляет 21 вид. Рассматриваются особенности гнездовой экологии: начало яйцекладки, размеры кладок, длительность инкубации, гнездовые биотопы и места расположения гнезд, приуроченность поселений куликов к другим видам птиц. Особые экологические условия гнездования куликов на водоемах Байкальской Сибири обусловливают формирование некоторых специфических адаптаций. К ним относят растянутые сроки размножения куликов, способность к возобновлению кладок в случаях их гибели, образование разных гнездовых поселений и высокую степень осторожности, агрессивность крупных и скрытность мелких видов куликов.
Ржанкообразные, кулики, байкальская сибирь, экология гнездования
Короткий адрес: https://sciup.org/148317989
IDR: 148317989 | УДК: 598.2:502.7(571.5) | DOI: 10.18101/2542-0623-2017-3-7-21
Peculiarities of the ecology of the waders in the Baikal Siberia
Specific Composition of waders nesting in Baikal Siberia includes 21 species. Features of nesting ecology were viewed - laying beginning, size of egg set, time of incubation, nesting biotopes and nest locations, vicinity wader’s colonies to another bird species. Special ecological conditions of waders nesting determine some special adaptations - extended reproduction time, ability for egg set recovery in case of their death, form different colonies, high level of caution, big waders are aggressivenessand small are secretive.
Текст научной статьи Особенности экологии гнездования куликов в Байкальской Сибири
Кулики — неотъемлемый компонент экосистем внутриконтинентальных озер аридных зон Северной Азии и одна из наиболее разнообразных групп птиц средних и высоких широт. Кулики принимают заметное участие в формировании орнитофауны приводных экосистем Байкальской Сибири.
Первые материалы по фауне куликов Байкальского региона изложены в трудах первых исследователей Сибири в конце XIX и начале XX веков. Известны труды И. Г. Георги [Georgi, 1775], Г. Радде [Radde, 1863], Б. И. Дыбовского и В. А. Годлевского [1870], В. К. Тачановского [1877]. В начале XX века сведения о куликах встречаются в трудах С. С. Турова [1923], К. А. Воробьева [1927], А. В. Третьякова [1934] и Б. К. Штегмана [Stegmann, 1929, 1936]. Далее материалы по биологии и экологии размножения куликов встречаются в работах О. К. Гусева [1960], Т. Н. Гагиной [1958; 1960; 1961; 1988 и др.] и В. А. Толчина [1977; 1979; 1983]. Вторая половина прошлого столетия по настоящее время связана с основными трудами и текущими исследованиями по биологии и экологии куликов сотрудников НИИ биологии Иркутского госуниверситета в разное время — В. Е. Журавлева, Ю. И. Мельникова, В. А. Подковырова, С. В. Пыжьянова, Н. Н. Сафронова, И. И. Тупицына, И. В. Фефелова и др. Сведения о куликах степных озер в данном регионе представлены преимущественно в работах эколого-фаунистического содержания [Моллесон, 1896; Козлова, 1930; Измайлов, Боровицкая, 1973] и статьях, опубликованных за последние десятилетия
[Доржиев, 1997; 2002; 2011; Доржиев и др., 1991; 1995; 1999; 2000; 2003;
2009; Малеев, 2001; 2003]. Текущие исследования по фауне и экологии куликов связаны с трудами сотрудников Бурятского госуниверситета — Ц. З. Доржиева, Э. Н. Елаева, Е. Н. Бадмаевой; сотрудниками Баргузинского заповедника — А. А. Ананина [2010] и др.
Автором целенаправленное изучение биологии и экологии куликов начато с 2002 года. За это время получен материал по срокам прилета, пролета, гнездовой биологии, успеха размножения, кормового поведения околоводных птиц на местах миграционных остановок и вопросов их охраны.
Цель настоящей работы — обобщить имеющиеся данные по гнездовой экологии куликов Байкальской Сибири.
Материал и методика
Статья основана на источниках, опубликованных в многочисленных научных изданиях, а также на собственных наблюдениях в период с 1999 года и по настоящее время в разных районах Байкальской Сибири. Основные данные по экологии гнездования куликов собраны по степным озерам и рекам в Оронгойской, Боргойской и Баргузинской котловинах. Рассмотрены сроки начала кладок, продолжительность насиживания, гнездовые стации и расположение гнезд, размеры кладок. Названия птиц даны классификации, принятой в работе Е. А. Коблика и В. Ю. Архипова [2014].
Результаты и обсуждение
Гнездящаяся фауна куликов Байкальской Сибири представлена 21 видом [Доржиев, Бадмаева, 2016; Мельников, 2011; Бадмаева, 2006; Фефелов и др., 2001], что составляет 35,5% от всей фауны куликов региона. В систематическом отношении гнездящиеся кулики представлены двумя семействами: Шилоклю-вковые Recurvirostridaе: ходулочник и шилоклювка (2 вида) и семейство Ржанковые Charadriidae: чибис, малый зуек, хрустан, вальдшнеп, горный дупель, азиатский бекас, лесной дупель, бекас, азиатский бекасовидный веретенник, большой веретенник, большой кроншнеп, травник, поручейник, большой улит, черныш, фифи, перевозчик, длиннопалый песочник, турухтан (19 видов) . Гнездование всех указанных куликов достоверно (описаны кладки и наблюдались выводки). Исключение составляет травник, поскольку его гнезд, кладок и выводков никто не находил. Сведения по биологии некоторых редких и охраняемых видов куликов в регионе отрывочны и требуют дополнительных исследований.
Рассмотрим основные характеристики по гнездовой биологии куликов.
-
1. Ходулочник Himantopus himantopus (Linnaeus, 1758) в регионе носит статус перелетного нерегулярно гнездящегося вида. Точных сведений по гнездованию не имеется. Кладок и птенцов не находили, хотя часто отмечается в гнездовой период по соленым и горько-соленым озерам Боргойской котловины, по открытым илистым берегам, часто в колонии шилоклювок.
-
2. Шилоклювка Recurvirostra avosetta Linnaeus, 1758. Сроки начала яйцекладки отмечены со II декады мая. Насиживание продолжается по II декаду июня. Вылупление птенцов наблюдали со II декады июня по I декаду июля [Бадмаева, 2004]. Гнездовыми стациями являются солончаковые грязи и отмели со скудной 8
-
3. Чибис Vanellus vanellu s (Linnaeus, 1758). Сроки яйцекладки со II по III декаду мая. Вылупление птенцов наблюдали во II–III декадах июня. Гнездовыми стациями являются сырые луга по поймам рек, травянистые болота и берега озер. Избегает мест с развитой кустарниковой и древесной прибрежной растительностью. Охотно гнездится в антропогенных ландшафтах [Измайлов, Боровицкая, 1973; Фефелов и др., 2001; Бадмаева, 2016]. Гнезда располагают открыто, по топким разнотравным лугам, примыкающим к водоемам; берегам соленых озер, всегда среди невысокой травы; пашням и залежам. Кладки состоят из 1–4 яиц (ср. 3,2). Длительность инкубации 24–26 дней [Толчин, 1984; Фефелов и др., 2001; Бадмаева, 2016].
-
4. Малый зуек Charadrius dubius Scopoli, 1786. Сроки начала кладок отмечены в период с III декады мая. Насиживание продолжается по III декаду июня. Птенцы вылупляются в период с I по II декаду июля. Гнездовыми стациями являются песчано-галечниковые берега и отмели рек, проток, пресных и соленых озер, ручьев. Гнезда всегда располагаются открыто на песке или галечнике вдоль береговой линии с редкой растительностью. Расстояние до уреза воды варьирует от 10 до 12 м, размер кладки — 1–4 яйца (ср. 3,4). Длительность инкубации 25– 26 дней [Бадмаева, 2003; 2016].
-
5. Хрустан Eudromias morinellus (Linnaeus, 1758). Известны лишь единичные находки кладок. Кладки находили в период с III декады мая по I декаду июня [Ананин, 2013]; на юге Якутии [Воробьев, 1963]. Пуховичков наблюдали в период с III декаду июня по начало июля [Ананин, 2013]; на юге Якутии пуховичков наблюдали во II декаду июля [Воробьев, 1963]. Гнездовыми стациями являются возвышенные сухие и каменистые тундры, в горах — гольцы со щебнистолишайниковой и дриадовой тундрой, с куртинами кедрового стланика и березки круглолистной, на хр. Хамар-Дабан — на высоте 1800–2300 м над ур. м. Гнездо представляет собой ямку на земле. Располагается на верхних частях возвышенностей и увалов, занятых щебенчатой тундрой. Среди мелких камешков, где лоток был выложен лишайниками [Успенский и др., 1962]. Размер кладок по югу Якутии — 1–3 яйца [Воробьев, 1963]. По длительности инкубации яиц в регионе сведений нет.
-
6. Вальдшнеп Scolopax rusticola Linnaeus, 1758. Начало откладки яиц со II по III декады июня. Птенцы вылупляются со II декады июля. Гнездовые стации в лиственных лесах с буреломом и кустарниками, в горах или в пойменных лесах с небольшими болотистыми участками. Гнездо располагает под негустой травой или под кустом, в мелкой ямке, лоток гнезда выстилает прошлогодней травой и листьями [Ананин, 2013].
-
7. Горный дупель Gallinago solitaria (Hodgson, 1831). По кладкам точных сведений не имеется. Пуховичков отмечали со II декады июля [Ананин, 2013]. Гнездится на субальпийских луговинах на верхней границе леса с зарослями карликовых ив, берез и обязательным присутствием водотоков на каменистом ложе (ключей, подтаивающих снежников) [Дурнев, 2009; Васильченко, 1982]. Размер кладки — 4 яйца. По длительности инкубации яиц в регионе сведений нет [Ана-нин, 2013].
-
8. Азиатский бекас Gallinago stenura (Bonaparte, 1830). В нашем регионе описанных находок кладок не имеется. По данным Воробьева [1963], по югу Якутии начало яйцекладки отмечено в период с I по II декаду июня. Вылупление — в III декаде июня. Гнездовыми стациями являются лиственничная тайга, горные леса, на сырых луговинах в альпийской зоне гор. Лесные болота, сырые гари и вырубки. Гнездо располагается на открытом месте, кочке, под прикрытием. Размер кладки — 4 яйца. По длительности инкубации яиц в регионе сведений нет [Воробьев, 1963].
-
9. Лесной дупель Gallinago megala Swinhoe, 1861. Пуховичков отмечали в I декаде июля. В качестве гнездовых стаций выбирает старые гари, разреженные березняки и осинники, в дельте р. Селенга — острова, поросшие ивняками и другими кустарниками, на севере — к лиственничникам, кедровникам с луговыми полянами и кустарником по влажным кочкарникам среди леса. Гнезда представляют собой ямки на сухих местах [Ананин, 2013]. По длительности инкубации яиц в регионе сведений нет.
-
10. Бекас Gallinago gallinago (Linnaeus, 1758). Начало кладок отмечено в III декаде мая по II декаду июня. По срокам вылупления и пуховичкам точных сведений нет. Гнездовые стации представлены сырыми лугами с кочковатыми болотцами, кустами ивняков в долинах рек, степные болотца, травянистые берега луговых и степных озер. Гнезда располагает на кочках по влажным осоковомоховым лугам, на заросших осокой берегах рек и озер [Фефелов и др., 2001]. По размерам кладок и длительности инкубации в регионе информации нет.
-
11. Азиатский бекасовидный веретенник Limnodromus semipalmatus (Blyth, 1848). Начало кладок отмечено с III декады мая по II декаду июня [Мельников, 2010; Фефелов и др., 2001]. Вылупление отмечено со II декады июня [Мельников, 2010]. Места гнездовий выбирает на грязевых отмелях и мелководьях, поросших осокой, хвощом или водяной сосенкой. Также низкие болотистые луга с топкой почвой, илистые мелководья, по плоским травянистым берегам озер. Избегают густых и сплошных зарослей, предпочитая разреженные участки [Мельников, 2010]. Гнезда располагает в очень сыром месте, часто среди воды, на гривке, кочке, в траве или почти открыто [Фефелов и др., 2001]. Гнезда на гривах разнотравных лугов, возвышениях микрорельефа подтопленных лугов или открытых грязей на кочках [Мельников, 2010]. Размер кладок составляет 1–4 яиц (ср. 1,8). Встречаются повторные (компенсационные) кладки. Длительность инкубации — 20–26 дней. [Мельников, 2010].
-
12. Большой веретенник Limosa limosa (Linnaeus, 1758). Сроки откладки яиц и насиживание в течение всего мая. Вылупление идет в течение всего июня. Гнездовые стации — это открытые травянистые сырые луга и болота без высокой растительности, реже на сухих лугах. Гнезда располагает на наиболее сухих участках или небольших кочках на влажных закочкареных лугах, пастбищах [Фефелов и др., 2001]. В Боргойской котловине нами было найдено 2 гнезда большого веретенника с кладками их двух яиц на кочковатом сыром лугу вблизи подземного источника в окр. степного соленого озера В. Белое (долина р. Джи-да). Кладка из 1–4 (3) яиц [Фефелов и др., 2001]. Нередко в непосредственной близости от колоний чайковых (сизых чаек и речных крачек).
-
13. Большой кроншнеп Numenius arquata (Linnaeus, 1758). Начало кладки приходится на I декаду мая и полностью формируется к I декаде июня [Фефелов и др., 2001]. Вылупление идет в III декаде июня [Измайлов, Боровицкая, 1973]. В качестве гнездовых стаций выбирает сухие остепненные участки островов дельты, не избегает влажных заливных лугов, предпочитая при этом наиболее сухие места [Фефелов и др., 2001]; сырые и заболоченные луга в окрестностях степных озер [Измайлов, Боровицкая, 1973]. Гнездо располагает на сухом участке среди невысокой травы [Фефелов и др., 2001]. Кладка из 4 яиц [Измайлов, Боровицкая, 1973; Фефелов и др., 2001]. Длительность инкубации известна — 28– 30 дней. Часто гнездовые участки вблизи колоний сизых или озерных чаек.
-
14. Травник Tringa totanus (Linnaeus, 1758) носит статус летующего и, возможно, гнездящегося вида. Точных сведений (гнезд и кладок, выводков) по размножению в регионе нет. Часто встречается по мелководьям степных соленых озер (наши данные) и реже в дельте р. Селенга [Швецов, Швецова, 1967; Толчин, 1977; Мельников, 1998].
-
15. Поручейник Tringa stagnatilis (Bechstein, 1803). Начало кладок приходится на II и III декады мая. Массовое вылупление идет во II декаде июня [Фефелов и др., 2001]. Гнездовые стации представлены сырыми лугами и травянистыми берегами озер, окраинами болот, низкими берегами степных рек. Гнезда устраивает на сухих участках среди густой травы, дающих укрытие; на кочках, на сплавинных островах среди водных зарослей. В кладке 4 яйца и длительность насиживания составляет 22–24 дня [Фефелов и др., 2001].
-
16. Большой улит Tringa nebularia (Gunnerus, 1767). Сведения по биологии этого вида очень скудны. Точных данных по находкам гнезд неизвестно. Известно, что нелетающих птенцов отлавливал Ю. И. Мельников в средней части дельты Селенги в I декаде июля [1998]. В кладках по 4 яйца и длительность инкубации составляет 24–25 дней.
-
17. Черныш Tringa ochropus Linnaeus, 1758. Точных сведений по биологии размножения этого вида в регионе также нет. Встречается часто в гнездовой период, но гнезд обнаружить еще не удавалось [Толчин, 1983; Юмов, Бадмаева и др., 1999]. Биотопами являются лесные болота и озерки, лесные речки с заболоченными берегами [Измайлов, Боровицкая, 1973]. В кладках по 4 яйца и длительность инкубации составляет 20–23 дня.
-
18. Фифи Tringa glareola Linnaeus, 1758. Приступает к откладке яиц в III декаде мая и насиживает до I декады июня. Гнездовые стации представлены открытыми пространствами дельты рек [Фефелов и др., 2001]; заболоченными лугами у озер, открытыми осоковыми болотцами, часто с кустиками ив [Измайлов, Боровицкая, 1973]; травянистые и моховые болота, стоячие водоемы, сырые поляны, не очень густые смешанные леса. Гнездо всегда на земле под прикрытием куста, трав, дерева, часто вдали от воды. В кладках по 4 яйца и длительность инкубации составляет 20–24 дня.
-
19. Перевозчик Actitis hypoleucos (Linnaeus, 1758). Кладки начинает формировать в III декаде мая и насиживает их до конца июня [Измайлов, Боровицкая, 1973; Фефелов и др., 2001]. В качестве гнездовых стаций выбирает небольшие реки с лесистыми берегами; по открытым травянистым берегам, голым отмелям; галечниковые и песчаные берега озер и рек, в том числе и таежных [Измайлов, Боровицкая, 1973]. Гнездо располагается на земле среди кустов, в лесу или относительно открыто, часто на значительном удалении от берега. В кладке 4 яйца, насиживает в течение 21–23 дней.
-
20. Длиннопалый песочник Calidris subminuta (Middendorff, 1851). Известны единичные находки гнезд: на заболоченном лугу у оз. Боргой от 25 июня 1966 г., с кладкой из 4 яиц [Измайлов, Боровицкая, 1973]; в устье реки Верхняя Ангара (Северный Байкал) от 12 июня 1973 года на моховом болоте между озером Светлым и протокой Акуликан с кладкой из 2 яиц [Толчин, Сонин, 1976]; Малое Море [Пыжьянов и др., 1979]. Гнездовой биотоп длиннопалого песочника на р. Верхняя Ангара представляет собой моховой покров с редкими куртинками осок и карликовой ивы на месте погибшего, после подъема уровня Байкала, чахлого лиственничника. Гнездо находилось на моховой кочке под кустиком осоки, дно лунки выстлано сухими листочками карликовой ивы.
-
21. Турухтан Phylomachus pugnax (Linnaeus, 1758). Начало кладок приходится на III декаду мая и насиживание продолжается до II декады июня [Толчин, 1983; Фефелов и др., 2001]. Гнездовые стации представлены мохово-осоковыми, часто сплавинными, участками калтусных болот, сухими прирусловыми валами проток и озер среди мелководных разливов, осоково-переувлажненными кочкар-никовыми лугами, часто чередующимися с ленточными лесными колками. Гнезда располагаются на сухих участках или кочках среди мелководных разливов, на моховых или осково-кочкарниковых болотах [Толчин, 1983].
растительностью, открытые прибрежные топкие мелководья степных соленых озер. Гнезда располагает вдоль береговой линии на песчаных или обсохших грязевых отмелях с редкой растительностью, часто сплошь покрытых выцветами солей. Удаленность гнезд от воды варьирует не более 100 м. Размеры кладок — 1–4 яйца (ср. 2,8). Редко встречаются 5–6 яиц (сдвоенные кладки). Длительность инкубации 24–25 дней. Селятся рыхлой колонией, устраивая от 2–3 до 32 гнезд. Расстояние между гнездами варьирует от 1 до нескольких метров. Иногда возможны одиночные гнездовья [Бадмаева, 2015].
По мнению П. С. Томковича [2014], «…в заметке A. A. Васильченко и В. В. Унжакова [1982] размеры яиц, приведенные для кладки азиатского бекасовидного веретенника из дельты реки Селенги, не укладываются в пределы изменчивости яиц этого вида. Величина кладки (3 яйца) также необычна для этого 10
веретенника, для которого характерна полная кладка из 2 яиц. Эти факты указывают на ошибочность видового определения птиц — обладателей указанного гнезда». Это можно объяснить «появлением кладок с одним яйцом, так же, как и с 3 и 4 яйцами, вызванных нарушениями в яйцекладке отдельных пар, связанных с гибелью неоконченных кладок. В таких случаях самки обычно подкладывают одно яйцо в чужое гнездо, но изредка они строят новое гнездо и откладывают в него одно яйцо» [Мельников, 2010, С. 148]. Нередко селится рядом с колониями чайковых (белокрылая крачка, малая, сизая и серебристая чайки), а также чибисом.
Общая продолжительность начала прилета птиц в регион охватывает середину — конец марта — конец мая. Кулики, в общем, прилетают позже других водно-болотных птиц и в более сжатые сроки, (вторая половина апреля — май, за исключением отдельных видов). В конце мая влажные и залитые водой участки начинают обсыхать и становятся пригодными для размещения гнезд, хотя травяной покров еще не установился. В это время большинство мелких водоемов 12
освобождается ото льда и прибрежные мелководья, прогреваясь, привлекают большое количество беспозвоночных. Для куликов очень важно наличие открытых водоемов с доступными кормовыми объектами. Поэтому появление большинства видов куликов и других ржанкообразных связано с освобождением ото льда мелководных и хорошо прогреваемых озер (конец апреля — первая половина мая), где очень быстро устанавливается хорошая кормовая база. Исключение составляют полифаги — чибис, чайки, речная крачка и некоторые другие, которые прилетают сравнительно рано (конец марта — первая половина апреля) [Доржиев, Бадмаева, 2017].
Гидрологический режим водоемов в этот период часто нестабилен и часть кладок подтапливается. Так, у водоплавающих птиц выявлена хорошо выраженная избирательность гнездовых местообитаний, проявляющаяся в преимущественном использовании ими только определенных стаций, в основе которой лежат экологические требования вида к качеству гнездовых угодий [Мельников, 2017].
Гнездовые стации куликов распространены крайне неравномерно и связаны с прибрежными биотопами, большей частью мелководными, заболоченными, илистыми грунтами. Излюбленными стациями являются дельты рек, протоки, побережья степных водоемов. Гнезда птицы устраивают на земле, часто под прикрытием трав или открыто.
Требовательность куликов к выбору мест гнездования всегда видоспецифична — встречаются виды как с узким комплексом требований, так и более пластичные в этом отношении. Размещение гнезд куликов по микрорельефу показывает их приуроченность к ровным (28%) или среднекочковатым стациям (66%). По характеру гнездования куликов делят на две экологическиe группы: устраивающие гнезда открыто на илистых и песчанно-галечниковых грунтах и более или менее укрыто: в траве, на кочковатых влажных субстратах . Так, у шило-клювки — ярко выраженного стенобионта, требовательность к условиям выражается в выборе мест устройства гнезд по солончаковым грязям и отмелям со скудной растительностью, открытым прибрежным топким мелководьям степных соленых и горько-соленых озер. Большой веретенник тяготеет к травянистым сырым лугам без высокой растительности, располагая гнездо на сухих участках или небольших кочках на влажных закочкаренных лугах, пастбищах. Малый зуек приурочен к песчано-галечниковым берегам и отмелям водоемов, располагая гнездо открыто на песке или галечнике вдоль береговой линии. Большинство улитов гнезда устраивают на сухих участках в плешинах прошлогодних трав, дающих укрытие; на кочках, на сплавинных островах среди водных зарослей и т. д. Чибисы — более пластичны, гнезда располагают открыто, по топким разнотравным лугам, примыкающим к водоемам; берегам соленых озер, всегда среди невысокой травы; пашням и залежам.
Массовая откладка яиц у куликов начинается от II декады мая и до конца месяца (рис. 1). Сроки начала яйцекладки достаточно продолжительны: с начала мая до I декады июля. У некоторых видов это связано с повторными (взамен утраченных) кладками, вследствие подтопления гнездовий, хищничества. Такая большая разница в сроках и продолжительности яйцекладки объясняется динамикой экологических условий региона, наступлением благоприятных периодов 13
для обитания разных видов в различное время. Естественно, определенную роль играют особенности биологии видов, включая продолжительность насиживания. Полные кладки большинства куликов традиционно состоят из 4 яиц. Большее количество яиц (5–6) часто связано со спаренными кладками, а кладки по 1 и 2 яйца часто бывают повторными. Массовое вылупление птенцов куликов начинается со II декады июня и продолжается до I июля; вылупление повторных кладок идет до середины июля. Следовательно, вылупившиеся из повторных кладок птенцы гораздо позже встают на крыло.
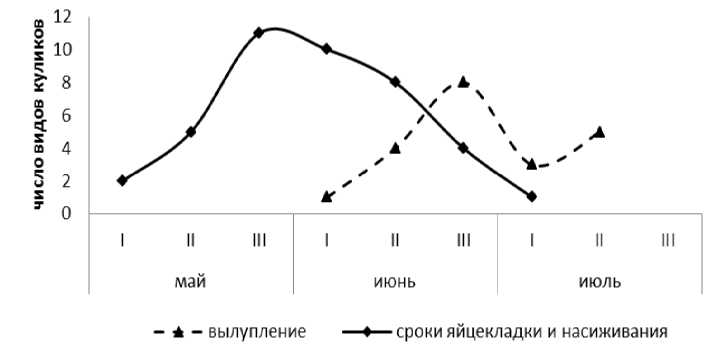
Рис. 1. Сроки откладки яиц, насиживания и вылупления птенцов у куликов в Байкальской Сибири
Реакция на приближение опасности у открытогнездящихся куликов (малый зуек, шилоклювка) проявляется в том, что они заранее покидают кладки (птенцов) и устраивают дальние демонстрации (позы, сигналы) на дальнем расстоянии. При приближении человека и др., объектов, несущих угрозу, птицы отлетают далеко от гнезда и садятся на открытых местах, часто проявляя имитацию насиживания. Все демонстрации сопровождаются тревожными голосовыми реакциями. Птенцы в это время, лоцируя сигналы родителей, затаиваются.
У куликов второй экологической группы защитные реакции иные. Птица может близко подпустить наблюдателя и выпорхнуть прямо из-под ног. Отвлекающие демонстрации часто исполняются прямо перед наблюдателем: птицы могут близко подходить или зависают в воздухе прямо над головой наблюдателя. Все это также сопровождается сигналами тревоги.
Особые экологические условия гнездования куликов на водоемах Байкальской Сибири обусловливают формирование некоторых специфических адаптаций. Можно выделить растянутые сроки размножения внутри популяций, способность к возобновлению кладок, в случаях их гибели, а также образование разных гнездовых поселений, высокая степень осторожности, агрессивность крупных и скрытность мелких видов куликов.
Список литературы Особенности экологии гнездования куликов в Байкальской Сибири
- Ананин А. А. Птицы Северного Прибайкалья: динамика и особенности формирования населения. Улан-Удэ: Изд-во Бурят. гос. ун-та, 2010. 296 с.
- Бадмаева Е. Н. Динамика видового состава и населения куликов (Charadriidae) в летний период в Юго-Западном Забайкалье // Вестник Бурятского университета. Сер. 2: Биология. 2004. Вып. 6. С. 182-184.
- Бадмаева Е. Н. К экологии гнездования шилоклювки и малого зуйка в Юго -Западном Забайкалье. Растения и животные в наземных экосистемах // Байкальский экологический вестник. 2003. Вып. 3. С. 91-97.
- Бадмаева Е. Н. Кулики степных озер Юго-Западного Забайкалья. Сибирская орнитология // Вестник Бурятского госуниверситета. Вып. 4. Спец. сер. 2006. С. 18-33.
- Бадмаева Е. Н. К экологии гнездования шилоклювки (Recurvirostra avosetta) в Юго-Западном Забайкалье // Сибирская зоологическая конференция: материалы всерос. конф., посвящ. 60-летию Института систематики и экологии животных СО РАН (15-22 сентября 2004 г.). Новосибирск: Изд-во ИСиЭЖ СО РАН, 2004. С. 219-220.
