Особенности экологии и морфологии толстуна Палласа (Deracantha onos Pall., 1772) в Юго-Западном Забайкалье
 в Юго-Западном Забайкалье")
Автор: Елаева Наталья Филипповна, Елаева Наталья Александровна
Журнал: Вестник Бурятского государственного университета. Философия @vestnik-bsu
Рубрика: Зоология
Статья в выпуске: 4, 2010 года.
Бесплатный доступ
Выявлены морфологические особенности кузнечика толстуна Палласа и проведен сравнительный анализ с особями Юго-Восточного Забайкалья. Описаны места обитания, встречаемость, характер расселения в Юго- Западном Забайкалье.
Кузнечик толстун палласа, масса тела, длина тела, переднеспинка, высота лба, голень, бедро, длина усиков
Короткий адрес: https://sciup.org/148179541
IDR: 148179541 | УДК: 565.728
The peculiarities of ecology and morphology of Derasantha onos Pall. in the South-East Zabaikalie
The morphological peculiarities of grasshopper Derasantha Onos Pall. are revealed, and the comparative analysis of specimen of the south-east and the south-west Zabaikalie was carried out. The localization, occurrence, settlement character in the studied region were described.
Текст научной статьи Особенности экологии и морфологии толстуна Палласа (Deracantha onos Pall., 1772) в Юго-Западном Забайкалье
Ареалы видов трибы Deracanthini мы приводим по М.Г. Сергееву [4]. Они ограничены в своем распространении Монголией и сопредельными территориями с аналогичными экологическими условиями. По литературным данным [1], подсемейство Deracanthinae можно рас- сматривать как древнейший элемент пустынностепной фауны, возможно связанной в своем происхождении с древнейшим материком Гондваны.
Морфологической особенностью кузнечика Deracantha onos является полная утрата крыльев и недоразвитые надкрылья, которые полностью заходят под крышу переднеспинки. Взрослое насекомое имеет темно-пеструю маскирующую окраску, которая хорошо сливается с разреженным тpaвocтoeм холодно-полынных степей. Это крупный кузнечик с массивным грузным телом, шаровидной головой. В степях ЮгоЗападного Забайкалья нами отмечены особи темной и светлой окраски. На переднеспинке и по бокам брюшка хорошо видны различные темные пятна.
Нами взяты основные морфометрические показатели (масса и длина тела, длина и ширина переднеспинки, высота лба, длина задней голени, длина заднего бедра и длина усиков), которые характеризуют весовые и линейные параметры представителей исследуемой популяции (табл. 1).
По массе тела самки превосходят самцов почти на 2 г, по длине – разница составляет около 15 мм. Ширина и длина переднеспинки у самцов и самок практически не отличаются. По другим морфологическим показателям самки превосходят самцов: высота лба у них больше, чем у самцов на 1 мм, длина задней голени – на
2 мм, длина заднего бедра – на 4 мм и длина усиков – на 2 мм. Заметно, что у представителей обоих полов соответственно длине тела меняются длина задних голеней, высота лба, длина зад- него бедра, длина усиков. То есть эти показатели имеют прямую корреляцию в отличие от показателей длины и ширины переднеспинки.
Таблица 1
Некоторые морфологические показатели толстуна Палласа (n = 150)
|
Показатели |
Взрослые особи |
|||
|
Самцы |
Самки |
|||
|
Lim |
M ± m |
Lim |
M ± m |
|
|
Масса тела, г |
2,9 – 5,2 |
4,61 ± 1,19 |
3,9-9,2 |
6,39 ± 2,06 |
|
Длина тела, мм |
39,8- 45,6 |
41,3 ± 0,31 |
5,2-6,5 |
56,6 ± 1,23 |
|
Длина переднеспинки, мм |
12,8-13,6 |
13,0 ± 0,14 |
13,9-14,8 |
14,3 ± 0,06 |
|
Ширина переднеспинки, мм |
7,8-9,5 |
9,0 ± 0,07 |
7,8-9,7 |
9,0 ± 0,06 |
|
Высота лба, мм |
10,9-11,7 |
11,5 ± 0,03 |
11,8-12,5 |
12,5 ± 0,09 |
|
Длина задней голени, мм |
14,8-16,0 |
15,5 ± 0,26 |
16,6-17,8 |
17,0 ± 0,35 |
|
Длина заднего бедра, мм |
15,1-16,8 |
15,7 ± 0,11 |
18,4-19,5 |
19,0 ± 0,19 |
|
Длина усиков, мм |
29,8-32,9 |
32,0 ± 0,4 |
33,3-34,8 |
34,5 ± 0,15 |
И.А. Дмитриева-Юргенсон приводит данные только по длине тела и массе толстуна Палласа из Юго-Восточного Забайкалья: самцы – длина тела 40-50 мм (средняя – 45), вес 4,8 – 10,0 г (средний – 6,5); самки – длина тела 57–67 мм (средняя – 62,5), вес 10,0-15,9 г (средний – 13,0) [2].
При сравнении наших данных (табл. 1) с таковыми И.А. Дмитриевой-Юргенсон видно, что особи из настоящих степей Восточного Забайкалья крупнее в весе и размерах тела. Сравнительно малые линейные и весовые показатели толстунов Палласа в нашем регионе могут объясняться более северным расположением района исследований и вытекающими отсюда суровыми абиотическими (климатическими, эдафическими и др.), биотическими (сравнительно слабое развитие степной растительности, влекущее плохую кормовую базу и защитные условия) экологическими факторами.
В связи с утратой летательных и прыгатель-ных способностей кузнечики обладают большими размерами и весом тела. Все конечности ходильные, но очень слабые, задние ноги легко отрываются, как и у всех прямокрылых. При вспугивании Deracantha onos старается уползти под ближайшие кустики, если он находился на растении, то падает или сползает вниз. По данным И.А. Дмитриевой-Юргенсон, «…на гладкой поверхности, лишенной растительности, кузнечики проходят за 10 секунд 1,5 метра, т.е. за 1 минуту могут пройти 9 м».
В наших экспериментах кузнечики бегали в 1,5–2 раза быстрее, правда, мы выставляли их на середину дороги (грунтовой, гудронированного шоссе). Они быстро, высоко приподняв свои толстые тела, убегали с открытого пространства в траву.
Кузнечик Deracantha onos относится к пассивным тамнобионтам . Этому способствует темно-пестрая маскирующая окраска, которая хорошо сливается с разреженным тpaвocтoeм холодно-полынных степей. В исследуемом регионе в зависимости от цвета субстрата местности встречаются особи как темной, так и светлой окраски. Массивное грузное тело с большой шаровидной головой, утрата способности летать и прыгать дополняют характеристику этой экоморфы. Насекомое со столь массивным туловищем не могло бы свободно передвигаться в густой растительности, поэтому данный вид наиболее многочисленнен на участках с угнетенной растительностью, со слабым проективным покрытием. Кроме того, стебли травянистых растений не выдерживают их массы тела. Кузнечики топически связаны со степными караганни-ками: листья караганы используются ими в пищу, а сами кусты – для укрытия и стрекотания. При ландшафтной классификации Deracantha onos относят к степным видам.
Излюбленными стациями данного вида являются сухие степи с разреженной растительностью и кустами караганы. За час сбора на участке настоящих степей можно набрать около 12 особей, их плотность при этом составляет 0,39 ос/м2. Здесь следует подчеркнуть, что для толстуна Палласа характерен мозаичный, а вернее групповой тип пространственной структуры, поэтому на небольшом участке можно встретить несколько особей подряд, а затем на довольно большом протяжении однотипного биотопа они не встречаются. Чаще попадаются самцы, что соответствует наблюдениям и других авторов, изучавших представителей семейства Bradypori-dae в разных регионах: Deracantha onos – в Восточном Забайкалье [2] и Bradyporus multitubercu-latus – в степях Предкавказья. По стрекотанию на линейном маршруте (100 м) можно насчитать до 20–25 особей.
Местами обитания этого вида являются хо-лодно-полынно-твердовато-осковые степи и частично ковыльно-дерновинно-злаковые степи, встречается он совместно с массовыми видами из семейства саранчовых – Pararcyptera microp-tera, Myrmeleotettix palpalis, Aeropedellus varie-gatus, Oedaleus decorus, Bryodema tuberculatum.
Нами кузнечик Deracantha onos также отмечен на участке ковыльных степей, что не характерно для представителей семейства Bradypori-dae . Здесь мы отмечаем его как малочисленный вид, за час сбора можно встретить только около 7 особей, причем все они самцы. Плотность при этом составляет 0,23 ос/м2. Нахождение его на этих участках можно объяснить близостью биотопов холодно-полынно-твердовато-осоковых и ковыльно-дерновинно-злаковых степей и свободным передвижением особей. Особенно активно передвигаются самцы в период размножения, в этот период можно наблюдать массовые передвижения кузнечиков по всем степным биотопам. Если на их пути встречается дорожное полотно, то они его преодолевают, и при этом огромное их количество погибает под колесами автомобилей.
Питание . По типу питания кузнечик относится к полифагам, но большее предпочтение отдает растительной пище. Как показали наши наблюдения, в природе и неволе спектр кормовых растений кузнечика Deracantha onos очень широкий. Обладая мощными челюстями, он включает в свой рацион более 20 видов растений. При содержании в садках кузнечикам были предложены овощные и плодовые культуры, которые также хорошо поедались. Отсутствие какой-либо пищи приводило к случаями каннибализма, при совместном содержании этих кузнечиков с саранчой наблюдались случаи поедания саранчи, даже если был в наличии растительный корм (свежие степные злаки).
Размножение . Взрослых насекомых в природе легко обнаружить по характерному стрекотанию. Стрекочут как самцы, так самки, что в целом не свойственно представителям кузнечиковых [3]. В общем же стрекотание самок данного вида, видимо, способствует встрече полов. В период размножения происходит скопление толстунов Палласа, мы отмечали группы до 6-7
особей, в которых обычно одна самка, а остальные самцы.
По Р.Д. Жантиеву и др., кузнечики семейства Bradyporidae издают призывные сигналы и звуки протеста. Для пения они обычно поднимаются на различные поверхности (камни, сурчины тарбаганов) или растения [3]. По данным наших наблюдений, кузнечики чаще взбираются на кусты караганы мелколистной, которые наилучшим образом подходят для столь грузных насекомых. Кроме того, в период размножения мы наблюдали, как самцы стрекотали на поверхности почвы, высоко приподнимаясь на конечностях, самки же передвигались, как бы ползая, волоча брюшко по земле, стрекотать они могли только остановившись. После оплодотворения самки иногда перестают издавать звуки, становятся очень осторожными, их трудно обнаружить в природе. Мы же наблюдали стрекотание оплодотворенных самок как в природе, так и при вольерном содержании.
По результатам наших наблюдений составлен календарь развития толстуна Палласа (табл. 2). Фаза яйца занимает большую часть цикла развития и продолжается с момента откладки – с августа до последней декады июня следующего года. Первые личинки начинали отмечаться нами в последней декаде июня, в Восточном Забайкалье стадия личинки начинается в июле [2]. Нахождение личинок в природе затруднено, так как они не стрекочут, стрекотание у кузнечика Deracantha onos начинается только на стадии имаго. Находясь на стадии личинки, кузнечики линяют 3-4 раза, при этом они взбираются на растения и повисают вниз головой, цепляясь задними ногами за стебель. Шкурка начинает лопаться со стороны брюшка, ближе к переднегруди. Весь процесс линьки продолжается около 30-40 минут. Сразу после линьки окраска кузнечика бледно-розовая, в течение часа она темнеет и приобретает характерную окраску. Личинки способны передвигаться сразу же после окончания линьки. Фаза личинки продолжается до конца июля. Имаго в массе встречаются с середины июля до середины сентября.
Массовое спаривание начинается в конце июля и продолжается почти весь август. Поведение Deracantha onos при размножении, по нашим наблюдениям, в Юго-Западном Забайкалье практически не отличается от такового в Восточном Забайкалье [7].
При вскрытии оплодотворенных самок можно обнаружить от 30 до 70 яиц, различимых простым глазом. Самки были вскрыты в разные сроки после оплодотворения. Судя по количеству яиц в яичнике, самки кузнечика отличаются большой потенциальной плодовитостью. Найденные нами в природе яйца располагались неглубоко, примерно в 3-4 см от поверхности почвы. Твердая и толстая скорлупа выполняет за- щитную функцию при низких зимних температурах. Размеры яйца в среднем не намного отличаются от популяции Восточного Забайкалья.
Календарь развития толстуна Палласа (1999-2005 гг.)
|
Количество генераций |
Май |
Июнь |
Июль |
Август |
Сентябрь |
Зимующая фаза |
|
1 2 3 |
1 2 3 |
1 2 3 |
1 2 3 |
1 2 3 |
1 2 3 |
|
|
Одна |
• • • |
• • ^^^^^^в |
^^^^^^™ ^^^^^^^ ^^^^^^^ + + |
+ + + • • |
+ + + • • • |
• • • |
Примечание: « • » - фаза яйца; « - » - активная фаза личинки; « + » - имаго
Таблица 2
Наблюдения показывают, что брачный период у Deracantha onos растянут на весь август и начало сентября. При содержании в садках мы также наблюдали высокую половую активность самцов, которая проявлялась в том, что в отсутствии самок, они пытались спариваться друг с другом.
Суточная активность кузнечика Deracantha onos изучалась в июле и августе, в период их наибольшей активности. Активность кузнечиков можно определить по звуку стрекотания. Первые признаки активности мы начинали отмечать через 4-5 часов после восхода солнца, т.е. к 9-10 часам утра. К этому времени они покидали ночные убежища и прогревали свое тело под лучами солнца. В качестве ночных убежищ кузнечики используют микроуглубления, чащу кустов караганы, норы степных грызунов. Мы неоднократно находили этих кузнечиков в полуразрушенных норах грызунов, И.А. Дмитриева-Юргенсон в своих исследованиях также отмечала вероятность этого явления [2]. Нам удалось понаблюдать за процессом отогрева от ночного холодового оцепенения: кузнечики с трудом поднимаются на растения или другие возвышенности, поворачивают свое тело, подставляя максимально большую его поверхность под воздействие солнечных лучей. В этот момент они малоподвижны, при прикосновении могут упасть на землю или медленно уползти в укрытие, там они замирают. Двигательная активность Deracantha onos в это время суток очень низкая, только после предварительного прогрева под утренними лучами солнца они начинают свое стрекотание.
В течение дня наблюдается два отчетливых пика активности, первый пик акустической и двигательной активности наблюдается в 12-13 часов, а второй, более высокий, – в 18-20 часов (рис. 1). На графике хорошо заметен спад активности в 14-16 часов, что объясняется высокой температурой воздуха. В период дневной жары толстуны Палласа прячутся в тени растений, в микронишах и других подобных местах.
В Юго-Восточном Забайкалье, по данным И.А. Дмитриевой-Юргенсон, регистрировалась акустическая активность как в солнечные, так и в пасмурные дни, а также и в дни с очень сильным ветром и в совсем безветренные. Однако за все время исследований не отмечалось активности (стрекотания) в пасмурную погоду и при сильном ветре.
Суточная активность толстуна Палласа
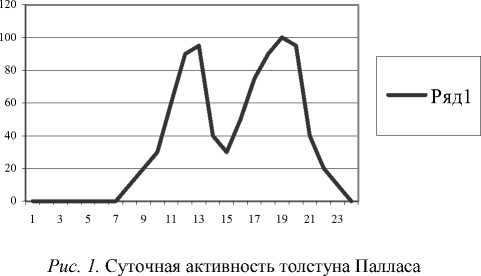
Вероятно, определяющим здесь является температурный градиент. Ночью в природе, впадая в холодовое оцепенение, кузнечики становятся неактивными, и мы не фиксируем их стрекотания. При содержании же их в садках, где нет суточных перепадов температуры, можно услышать их ночное стрекотание. В исследованиях, проведенных в Восточном Забайкалье И.А. Дмитриевой-Юргенсон, указывается, что «… в неволе ночью кузнечики остаются в том же неподвижном положении, на вершинках травы, близко у почвы, что и днем». Можно предположить, что И.А. Дмитриева-Юргенсон, содержала кузнечиков в открытых вольерах, где температурный фактор воздействовал также, как в природных условиях.
Таким образом, кузнечик Deracantha onos в степях Юго-Западного Забайкалья является обычным видом с мозаичным распространением, встречается только в настоящих степях с разреженным травянистым покровом. Реликто-вость толстуна Палласа обусловлена особенностями морфологии и экологии вида. Для него характерен неравномерный тип пространственной структуры, часто отмечаются группировки. Поэтому на небольших благоприятных для вида участках однородного местообитания отмечается плотность до 0,39 особей/м². По морфометрическим показателям представители исследуемой популяции мельче, чем кузнечики в Восточном Забайкалье. Это мы объясняем узким ареалом вида. Наличие темных и светлых по окраске форм легко объяснимо и обусловлено микро- биотопическими особенностями. Общеизвестен классический пример промышленного меланиз-ма у пядениц в Англии. Понятно, что крупные малоподвижные толстуны Палласа, обитающие на участках со слабым проективным покрытием, в случае отсутствия покровительственной окраски оказывались бы слишком легкой добычей для многих хищников. Наблюдаемые нами отчетливые распределения определенных морфоэкологических типов прямокрылых по соответствующим биотопам являются результатом непрерывно работающего эколого-эволюционного механизма борьбы за существование. Личиночный цикл развития обусловлен абиотическими факторами среды и считается особенностью изучаемых популяций. Выявленные способности к стрекотанию у оплодотворенных самок, вероятно, связаны с физиологией насекомых.
