Особенности экспрессии гена CCND1 (BCL1) циклина DL-регулятора клеточного цикла в различных структурах мозга при реализации долговременной памяти у взрослых крыс
 циклина DL-регулятора клеточного цикла в различных структурах мозга при реализации долговременной памяти у взрослых крыс")
Автор: Грудень Марина Алексеевна, Ратмиров Александр Максимович, Сторожева З.И.
Журнал: Академический журнал Западной Сибири @ajws
Рубрика: Эксперимент
Статья в выпуске: 3 (80) т.15, 2019 года.
Бесплатный доступ
Целью настоящего исследования явилось сравнительное изучение уровня транскрипционной активности гена CCND1 (BCL1), продукт которого циклин D1 принимает участие в регуляции Gi-фазы клеточного цикла в релевантных церебральных структурах при формировании долговременной пространственной памяти у половозрелых крыс самцов Wistar. Методы. В поведенческих экспериментах использовали 2-е группах животных, а именно, группу крыс, подвергнувшихся принудительному плаванию (активный контроль) и группу обученных животных, у которых вырабатывали пространственный навык нахождения скрытой платформы в водном лабиринте Морриса. В качестве контроля служили нативные животные того же возраста и пола, не проходившие экспериментальный протокол. В генетических экспериментах для изучения экспрессии гена CCND1 (BCL1) в церебральных структурах в группах нативных крыс, «активного контроля» и обученных животных применяли метод ПЦР в режиме реального времени. Результаты. По казано, что критерием установления прочной долговременной памяти являлся тот факт, что время достижения платформы в конце 4-го сеанса на 4-й день обучения составляло менее 10 секунд...
Пространственная память, клеточный цикл, гены, регуляция, гиппокамп, префронтальная кора, мозжечок, крысы
Короткий адрес: https://sciup.org/140248157
IDR: 140248157
Peculiarities of CCND1 (BCL1) gene expression of cell cycle regulator the cyclin D1 in various brain structures during long-term memory realization in adult rats
The main goal was a comparative study of CCND1 (BCL1) gene transcriptional activity, the product of which cycline D1 participates in the regulation of the G1 phase of the cell cycle in relevant cerebral structures during the formation of long-term spatial memory in male mature rats Wistar. Methods. In behavioral experiments, 2 groups of animals were used, namely, a group of rats subjected to forced swimming (active control) and a group of trained animals, who developed the spatial skill of finding a hidden platform in the Morris water maze.Native animals of the same age and sex, not passing the experimental protocol, served as controls. In genetic experiments, real-time PCR was used to study the expression of the CCND1 (BCL1) gene in cerebral structures in groups of native rats, “active control” and trained animals. Results. It цфы shown that the criterion for establishing solid long-term memory was the fact that the time to reach the platform at the end of the 4th session on the 4th day of training was less than 10 seconds...
Текст научной статьи Особенности экспрессии гена CCND1 (BCL1) циклина DL-регулятора клеточного цикла в различных структурах мозга при реализации долговременной памяти у взрослых крыс
Современные данные о явлениях в мозге, имеющих место при формировании долговременной памяти легли в основу представления о том, что развитие памяти является каскадным процессом, который вовлекает в себя последовательные стадии приобретения навыка, консолидации и реконсолидации памяти, извлечение памятного следа с его последующим угашением [1]. Изучение молекулярных событий, имеющих место на каждой стадии показало, что в функционировании механизмов памяти, например, пространственной памяти, участвуют многочисленные сигнальные молекулы, которые в ряде случаев могут включать или переключать молекулярные пути реализации определенных этапов ее формирования [2]. При этом, тонкие молекулярные настройки клеточного цикла у делящихся клеток при реализации долговременной пространственной памяти, а также роль регуляторов клеточного цикла, таких как циклина D1 [3] в процессе ее формирования, изучены недостаточно [4]. В связи с этим, необходимо подчеркнуть, что характерным свойством пролиферирующих клеток является их способность к делению и у животных клеток интервал между митозами (клеточный цикл, точнее митотический цикл) составляет примерно 10-24 ч. За это время клетка проходит четыре фазы жизненного цикла: G1-фазу начального роста, S-фазу удвоения молекул ДНК (репликации), G2-фазу роста и М-фазу клеточного деления. В совокупности фазы G1, G0, S и G2 носят название интерфазы, которая в клеточном цикле сменяется существенно более короткой фазой митоза (М). Наиболее детально изучена фаза клеточного деления, митоз (М-фаза). В G1-фазе, продолжительность которой может сильно варьировать, происходит синтез мРНК, белков и других компонентов клетки. Клетки, которые прошли дифференцировку и больше не делятся, постоянно находятся в фазе покоя G0. Белок цик-лин D1 является важным регулятором прогрессирования фазы G1 в S во многих различных типах клеток, в том числе и нервных. Было показано, что циклин D1 является нестабильным соединением с коротким периодом полураспада (~24 мин) и разлагается главным образом через протеасому 26S убиквитин-зависимым образом [5]. Вместе с тем, циклин D1 и его молекулярные партнеры по связыванию циклин-зависимой киназой 4 и 6 (CDK4 и CDK6) образуют активные комплексы, которые способствуют прогрессированию клеточного цикла путем фосфорилирования и инактивации бел-ков(RB)/ Известно, что циклин D1 также функционирует как модулятор транскрипции, регулируя активность нескольких факторов транскрипции и деацетилазы гистона (HDAC3) [5]. В совокупности рассмотренных данных, регуляция экспрессии его гена CCND1 (BCL1; BCL1/cyclin D1) имеет важное значение для поддержания необходимого его содержания в клетках. Особенный интерес может представлять регуляция его экспрессии в условиях различных физиологических экспериментов. Учитывая, что одним из эффективных подходов, наряду с фармакологическими манипуляциями [6] и парадигмами обучения, является использование генетического подхода, что способствовало выявлению определенных церебральных структур, молекулярных механизмов и белков, участвующих в формировании различных видов памяти, включая пространственную память [7].
В этой связи, функциональные исследования транскрипционной активности генов нервных клеток особенно информативны при определении региональных молекулярных особенностей протекания различных этапов формирования памяти. В результате исследований процесса транскрипции генов в условиях обучения животных в водном лабиринте Морриса в дополнение к изменениям в экспрессии ранних генов [8] была также обнаружена динамическая картина экспрессии поздних генов при формировании пространственной памяти [7].
Цель настоящей работы состояла в сравнительном исследовании уровня транскрипционной активности гена CCND1 (BCL1) в гиппокампе, префронтальной коре и мозжечке взрослых крыс Wistar при формировании гиппокамп-зависимой пространственной памяти.
Материалы и методы.
Исследование было проведено на 30-ти крысах самцах линии wistar в возрасте 3-х месяцев и массой 220,4±24,1 гр. (ФГБУН НЦБМТ ФМБА питом- ник «Столбовая», Россия). Все манипуляции с животными были проведены с соблюдением требований, изложенных в директиве Европейского парламента и Совета европейского союза (2010/63/ EU от 22 сентября 2010 г.), а также в соответствии с правилами, утвержденными комиссией по биоэтике ФГБНУ «Научно-исследовательский институт нормальной физиологии им. П.К. Анохина». Животные содержались в стандартных условиях вивария при свободном доступе к пище и воде и 12-ти часовом световом режиме. Крысы были разделены на три э группы: группа 1 – нативные крысы (n=10), группа 2 – «активный контроль» (принудительно плавающие) крысы (n=10), группа 3 – обученные крысы в водном лабиринте (n=10). Эксперименты по обучению и тестированию пространственной памяти проводили на модели водного лабиринта Морриса (Columbus Instruments, USA) по протоколу, описанному ранее [9]. Экспериментальный протокол составлен таким образом, что время плавания в группе активного контроля соответствовало времени, проведенному в воде обучавшимся животным, т.е. каждому обучавшемуся животному по времени и паттерну плавания соответствовала одна “контрольная” особь. Через 24 ч по окончании поведенческих экспериментов всех животных декапитировали и извлекали на холоду (+4ºС) структуры мозга: гиппокамп, префронтальную кору и мозжечок, которые использовали для изучения экспрессии гена CCND1 (BCL1) методом ПЦР в режиме реального времени по описанному ранее протоколу, применяя в качестве референтного – ген β-актина для последующего расчёта относительно уровня экспрессии изучаемых генов по методу 2-ΔΔCt [10]. Экспрессия гена CCND1 (BCL1) 2 и 3–й группах была оценена относительно интактных животных из группы 1, где экспрессия гена CCND1 (BCL1)была принята за единицу.
Статистическую обработку полученных результатов проводили по алгоритмам программы “Statistica 7,0”. При сравнении нескольких независимых выборок применяли однофакторный непараметрический дисперсионный анализ по методу Крускал-ла-Уолиса (Н-критерий) с последующим post-hoc анализом по U-критерию Манна-Уитни. Данные представлены в виде M±m. Критическое значение уровня статистической значимости при проверке нулевых гипотез принималось равным 0,05.
Результаты и обсуждение
Для выполнения задачи данной работы по изучению экспрессии гена CCND1 (BCL1)в церебральных структурах крыс при формировании пространственной памяти на начальном этапе исследования были выполнены поведенческие эксперименты по выработке долговременной пространственной памяти в водном лабиринте Морриса. Было документировано, чт при обучении среднее время для достижения платформы у обученных крыс по сравнению с группой животных « активного контроля» во втором и последующих сеансах было ниже, чем в первом сеансе обучения p<0,01 и p <0,05 соответственно). Обнаружено, что до конца последнего сеанса на четвертый день обучения среднее время достижения платформы составляло менее 10 секунд, что свидетельствует о формировании стабильной долговременной пространственной памяти (рис.1). Анализ средних значений для 2-го, 3-го и 4го сеансов обучения в сравнении с первым сеансом помещения в водный бассейн в каждый конкретный день, выявил следующее значение различий в среднем времени достижения платформы: 1-й день - 2-е испытание = p<0,05, 3-й и 4-й испытания = p<0,01; 2-й день - 4-е испытание = p<0,05; 3-й день - 2-е испытание = p<0,05, 3-е и 4-е исследования = p<0,01; 4-й день - 2-й, 3-й и 4-й испытания = p<0,01 (рис. 1).
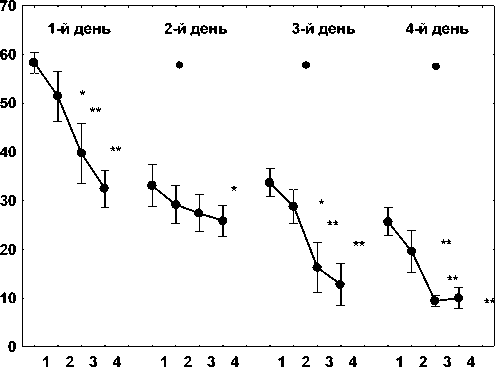
Примечание: По оси абсцисс - номер пробы, по оси ординат - время достижения платформы (с). * - p <0,05, ** - p <0,01 по сравнению с первой пробой соответствующего дня; • -статистически значимое (p<0,05) снижение среднего значения по всем пробам относительно аналогичного показателя предыдущего дня.
Рис. 1. Формирование долговременной простран ственной памяти у половозрелых крыс Wistar в водном лабиринте Морриса.
Сравнение и анализ группового времени достижения платформы животными в водном лабиринте Морриса показали следующие существенные отличия: во 2-й и 4-й дни время достижения платформы для первых испытаний значительно отличались от первых сеансов в дни 1-й и 3-й, соответственно (p<0,05). Документировано, что общие показатели для всех временных испытаний в дни 2, 3 и 4 значительно отличались от таковых в предыдущие дни, соответственно (p <0,05). Так как время достижения платформы в конце четвертого сеанса на четвертый день обучения составляло менее 10 секунд, это указывало на установление стабильной долговременной пространственной памяти у экспе- риментальных животных (рис. 1) в отличие от крыс из группы «активного контроля», подвергшихся принудительному плаванию в бассейне.
По окончанию поведенческих экспериментов в работе были проведены молекулярно-генетические исследования экспрессии гена CCND1 (BCL1) гиппокампе, префронтальной коре и мозжечке в головном мозге как контрольных («нативный» контроль и «принудительное плавание»), так и обученных животных существует региональная специфичность в экспрессии гена CCND1 (BCL1).
Проведенные в работе эксперименты показали, что у животных, которых подвергли физической нагрузке и стрессу новизны в водном лабиринте Морриса ( активный контроль) в гиппокампе наблюдали снижение активности гена CCND1 (BCL1) на 57% (P<0,05), при этом префронтальная кора манифестировала выраженной его активацией -3,2 раза (P<0,01), а в мозжечке экспрессия гена CCND1 (BCL1) оставалась на уровне интактных животных (рис. 2).
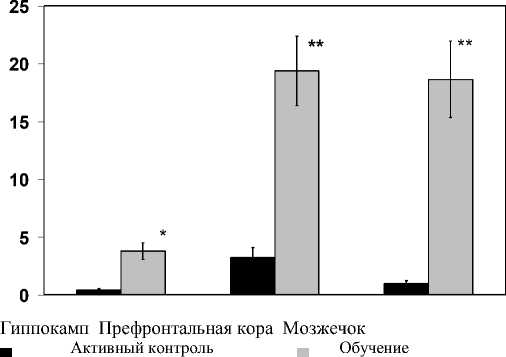
Примечание: По оси абсцисс - структуры мозга, по оси ординат -усл. ед. от нативного контроля. * - p <0,05 по сравнению с нативным контролем. ** - p <0,01 по сравнению с нативным контролем
Рис. 2. Уровень экспрессии гена CCND1 (BCL1) цик-лина D1 в гиппокампе, префронтальной коре и мозжечке у «активного контроля» и обученных пространственному навыку в водном лабиринте Морриса половозрелых крыс Wistar.
Эти данные указывают на то, что физическая нагрузка и стресс подавляют экспрессию гена цик-лина D в гиппокампе, что косвенно может влиять на количество пролиферирующих клеток в данной структуре. Кроме того, плавание и стресс не влияют в мозжечке, вовлеченного в обеспечение двигательной активности животных на активность гена CCND1 (BCL1) в условиях данного экспериментального протокола. Полученный факт, предположительно, может указывать на то, что скорость пролиферации нервных клеток в мозжечке остается неизменной и не зависит от физической нагруз- ки/стресса у животных при принудительном плавании в водном лабиринте. Обратная картина наблюдалась в префронтальной коре крыс из группы «активного контроля», где была выявлена значительное увеличение экспрессии гена CCND1 (BCL1) в ответ на принудительное плавание и стресс у крыс (рис. 2). Реакция на стресс - это не только первичный ответ, характерный для многих видов животных от саламандры до человека и вызывающий нарушения работы некоторых участков мозга. Недавние исследования выявили удивительный факт: в реакции на стресс также участвует и префронтальная область коры - зона мозга, находящаяся непосредственно за лобной костью и выполняющая функции центра управления высшими когнитивными способностями, в число которых входят концентрация внимания, планирование, принятие решений, понимание ситуации. Префронтальна кора -наиболее эволюционно молодая область мозга, и она особенно чувствительна даже к мимолетным тревогам и страхам, с которыми столкнулись животные при помещении в незнакомую среду в виде воды. Когда все идет хорошо, данная структура выступает в роли координатора, удерживающего в узде наши основные эмоции и импульсы. Но сильный и неконтролируемый стресс, как показали новые исследования, приводит к запуску каскада биохимических реакций, ослабляющих влияние префронтальной коры, в результате чего поведение начинают контролировать более эволюционно древние зоны мозга.
Особый интерес представляют полученные в работе генетические данные в условиях сформированной долговременной пространственной памяти, где паттерн экспрессионной активности гена CCND1 (BCL1)в релевантных церебральных структурах крыс изменяется по сравнению с обоими кон-тролями. Учитывая, что у животных формировалась гиппокамп - зависимая память, важны были результаты по активности гена циклина D в данной структуре. При изучении экспрессии гена CCND1 (BCL1) в образцах ткани гиппокампа экспериментальных животных было обнаружена повышенная активация гена циклина D в 3,8 раза (P<0,05) и 4,3 раз (P<0,05), по сравнению с интактным контролем или группой животных «активного контроля», соответственно. В префронтальной коре у обученных пространственному навыку животных наблюдали более выраженный по сравнению с гиппокампом рост активности гена CCND1 (BCL1) в 3,1 раза (P<0,05) и в 16,2 (P<0,05) по сравнению с нативными крысами и группой «активного контроля», соответственно (рис. 2). Удивительный факт, что в мозжечке обученных пространственному навыку животных и с реализованной на стадии тестирования пространственной памятью экспрессия гена циклина D возрастает в 18,6 раз (P<0,01) по сравнению с обоими контролями (рис. 2). Полученные факты свидетель- ствуют, что тонкие механизмы функционирования клеточного цикла нервных клеток оказались чувствительны к процессу становления пространственной памяти, а сам циклин D, вероятно, может быть вовлечен в их регуляцию через влияние на транскрипционные процессы других генов и наработки транскрипционных факторов, неоьходимых для запоминания.
На современном этапе развития нейробиологии дальнейшее изучение вклада генома в становление долговременной памяти продолжает оставаться актуальной задачей. Кроме того, понимание роли мРНК и синтеза белка в этом процессе, несомненно, облегчат дальнейшее изучение наименее исследованных процессов памяти как консолидация и реконсолидация [11] памяти, как в норме, так и при амнезиях [12]. Необходимо подчеркнуть, что функциональные свойства конкретных областей мозга определяются также в значительной степени генами, которые экспрессируются в нервных и глиальных клетках, локализованных в конкретных церебральных структурах. Очевидно, что субклеточные процессы индукции и экспрессии генов, особенно в гиппокампе, скорее всего, лежат в основе и навигационного обучения и формирования пространственной памяти. Тем не менее, транскрипционные процессы в гиппокампе, вероятно, могут варьировать в зависимости от активности генов в других областях мозга, таких как кора головного мозга и мозжечок, но они, вероятно, также могут изменяться при трансформации двигательной активности животных в водном лабиринте Морриса [13]. Ранее было продемонстрировано, что в процессе формирования пространственной памяти участвует гиппокамп, причем далее необходима последующая транслокация памятного следа в кору мозга для долговременной его консолидации [13]. В этот процесс также вовлечен мозжечок, хотя традиционный взгляд на основные функции мозжечка состоит из регулирования двигательной координации, баланса и моторной речи. В дополнение к координации двигательной активности мозжечок вовлечен в моторное обучение и более высокие познавательные функции, но схемы его участия в формировании когнитивных функций изучены недостаточно, хотя и имеются недавние свидетельства анатомической и функциональной связи, поддерживающие взаимодействие мозжечка и гиппокампа [14]. В этом контексте долговременная депрессия мозжечка может поддерживать общий сенсорный процесс адаптации, разделяемый как функциями моторного, так и пространственного обучения [14].
Таким образом, сравнительный анализ экспрессии гена регулятора клеточного цикла циклина D в гиппокампе, коре мозга и мозжечке при формировании долговременной пространственной памяти выявил специфические особенности активности гена CCND1 (BCL1) в изучаемых церебральных структурах. Показано усиление значительное активности гена CCND1 (BCL1) в изученных структурах у обученных животных по сравнению с интактными животными и крысами, подвергнувшихся «принудительному плаванию». Отмечено, что наиболее выражено повышение экспрессии гена циклина D в префронтальной коре и мозжечке, что, вероятно, отражает пролиферирующую способность нервных клеток в этих областях мозга. Ранее мы показали на генетическом уровне прямую связь между экспрессией генов, вовлеченных в регуляцию нейрогенеаза, апоптоза, NOTCH сигнальной трансдукции в отдельных областях мозга у нативных взрослых крыс при сформированной пространственной памяти [15]. Особенно интересно, что экспрессия гена CCND1 (BCL1), продуктом которого является цикли важный регулятор клеточного цикла может коррелировать с экспрессией ряда генов-регуляторов нейрогенеза и апоптоза, которая была наиболее выраженной в тех же структурах мозга животных при формировании пространственной памяти, которые были изучены в данной работе [16]. Полученные результаты могут быть полезными для понимания роли белков – регуляторов клеточного цикла на стадии консолидации пространственной памяти у взрослых животных.
Список литературы Особенности экспрессии гена CCND1 (BCL1) циклина DL-регулятора клеточного цикла в различных структурах мозга при реализации долговременной памяти у взрослых крыс
- Anderson J.R. Learning and memory; an integrated approach. 2nd Edition. John Wiley 2000. Р. 377.
- Abel T., Lattal K.M. Molecular mechanisms of memory acquisition, consolidation and retrieval // Current Opinion in Neurobiology. 2001. № 11. Р. 180-187.
- Lew D.J., Dulic V., Reed S.I. Isolation of three novel human cyclins by rescue of G1 cyclin (Cln) function in yeast // Cell. 1991. V. 66, № 6. Р. 1197-1206.
- Fang F., Lin W., Ling X. et al. The hippocampal cyclin D1 expression is involved in postoperative cognitive dysfunction after sevoflurane exposure in aged mice // Life Sci. 2016. № 160. Р. 34-40.
- Ma J. et al. Over-Expression of Cyclin D1 Promotes NSCs Proliferation and Induces the Differentiation into Astrocytes Via Jak-STAT3 Pathways // Neurochem Res, 2015.
- Diergaarde L., Schoffelmeer A.N.M., De Vries T.J. Pharmacological manipulation of memory reconsolidation: Towards a novel treatment of pathogenic memories // European Journal of Pharmacology. 2008. № 585. Р. 453-457.
- Грудень М.А., Сторожева З.И. Интеграция экпрессии генов, регулирующих нейрогенез в процессе фиксации пространственного памятного следа // Академический журнал Западной Сибири. 2018. Т. 14, № 3 (74). С. 78-83.
- Okuno H. Regulation and function of immediate-early genes in the brain: beyond neuronal activity markers // Neurosci Res. 2011. V. 69, № 3. Р. 175-186.
- DOI: 10.1016/j.neures.2010.12.007
- Sewell R.D., Gruden M.A., Pache D.M. et al. Does the human leukaemia differentiation factor fragment HLDF6 improve memory via brain DNA and protein synthesis? J Psychopharmacol. 2005. V. 19, № 6. Р. 602-608.
- Kolobov V.V., Storozheva Z.I., Gruden M.A., Sherstnev V.V. Regional Features of the Expression of Genes Involved in Neurogenesis and Apoptosis in the Brain of Adult Rats // Bulletin of Experimental Biology and Medicine. 2012. № 153. Р. 707-711.
- Squire L.R., Genzel L., Wixted J.T., Morris R.G. Memory consolidation // Cold Spring Harb Perspect Biol. 2015. V. 3, № 7(8): a021766
- Rosenbaum R.S., Murphy K.J., Rich J.B. The amnesias // Wiley Interdiscip Rev Cogn Sci. 2012. V. 3, № 1. Р. 47-63.
- Sekeres M.J., Winocur G., Moscovitch M.The hippocampus and related neocortical structures in memory transformation // Neurosci Lett. 2018. № 680. Р. 39-53.
- Rochefort C., Lefort J.M., Rondi-Reig L. The cerebellum: a new key structure in the navigation system // Frontiers in Neural Circuits. 2013. № 7. Р. 1-12.
- Gruden' M.A., Storozheva Z.I., Ratmirov A.M., Sherstnev V.V. Pattern of Notch2, Numb, and Cas8 Gene Expression in Relevant Structures of the Rat Brain during Formation of Spatial Memory // Bull Exp Biol Med. 2017. V. 3, № 6. Р. 785-788.
- Nobs L., Baranek C., Nestel S. et al. Stage-specific requirement for cyclin D1 in Glial progenitor cells of the cerebral cortex // Glia. 2014. V. 62, № 5. Р. 829-839.
