Особенности мезоструктуры, пигментного и липидного состава представителей сем. Onagraceae

Автор: Богданова Е.С., Розенцвет О.А., Нестеров В.Н., Зубова Светлана Николаевна
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 1 т.27, 2018 года.
Бесплатный доступ
Исследованы особенности мезоструктуры, пигментного и липидного состава представителей родов Oenothera и Epilobium сем. Onagraceae. Высокие значения фотосинтетических пигментов, гликолипидов и жирных кислот отмечали в листьях растений E. adenocaulon и E. hirsutum. Результаты кластерного анализа показали, что мезоструктура фотосинтетического аппарата определяется видовыми особенностями, а биохимические показатели имеют межвидовое сходство.
Липиды, мезоструктура, пигменты
Короткий адрес: https://sciup.org/148315102
IDR: 148315102 | УДК: 581.192+582.962 | DOI: 10.24411/2073-1035-2018-10006
Текст краткого сообщения Особенности мезоструктуры, пигментного и липидного состава представителей сем. Onagraceae
1 Семейство Onagraceae (кипрейные или ослинниковые) включает около 25 родов и до 700 видов растений. Как правило, это многолетние травы, реже кустарники и небольшие деревья. Многие виды этого семейства относятся к адвентивной флоре (Асминг, 2007).
2. МАТЕРИАЛЫ И МЕТОДЫ
Изучению адвентивной флоры в последнее время уделяется большое внимание. Считают, что чужеродные растения чутко реагируют на трансформацию экосистем и приобретают селективные преимущества по сравнению со многими аборигенными видами. Это позволяет им в пределах нового ареала резко повысить свою численность и успешно захватить новые территории. Внедрение адвентивных видов в растительные сообщества снижает биоразнообразие, в результате обедняется их видовой состав и упрощается структура (Ямалов и др., 2013). Управление этими негативными процессами в сообществах является на сегодняшний день актуальной проблемой науки о растительности. Для этого необходимы знания, как об экологии адвентивного вида, так и о его структурно-функциональных особенностях (Broz et al., 2007). Следует отметить, что работ подобного рода, посвященных адвентивным растениям, крайне мало (Shikov et al., 2006).
Цель настоящей работы состояла в исследовании особенностей мезоструктуры, пигментного и липидного состава растений сем. Ona-graceae. В задачу исследования входило также выявление возможной взаимосвязи между ме-зоструктурными параметрами фотосинтетического аппарата и биохимическими характеристиками растений в зависимости от их эколого-ценотической приуроченности и генетического статуса вида.
В качестве объектов исследования были выбраны растения Oenothera biennis L. (ослинник двулетний или энотера двулетняя), Oenothera villosa Thunb. (энотера мохнатая), Chamaene-rion angustifolium (L.) Scop. (иван-чай узколистный), Epilobium adenocaulon Hausskn. (кипрей железистостебельный), Epilobium hirsutum L. (кипрей волосистый). Растения отбирали на территории Сызранской и Ульяновской обл., в первой половине дня в середине июля 2016 г. Для анализов пигментов и липидов использовали листья 15–20 растений. Из усредненной массы составляли три биологические пробы по 0,5–2 г сырой массы. Далее растительный материал фиксировали в жидком азоте при температуре -80ºС.
Содержание фотосинтетических пигментов определяли на спектрофотометре «ПромЭко-Лаб ПЭ_3000 УФ» (Россия) в ацетоновой вытяжке. При длинах волн 662 и 644 нм определяли содержание хлорофилла (Хл а , б ), 470 нм – содержание каротиноидов (Кар) (Maslova et al., 1986).
Липиды экстрагировали смесью хлороформа и метанола (1:2) с одновременным механическим разрушением тканей. Разделение липидов осуществляли методом тонкослойной хроматографии (Кейтс, 1985). Количество фосфолипидов (ФЛ) определяли по содержанию неорганического фосфора, гликолипидов (ГЛ) и нейтральных липидов (НЛ) – денситометрически «Денскан-04» (Ленхром, Россия). Хроматограммы анализировали в режиме параболической аппроксимации по градуировочным зависимостям, используя моногалактозилдиацилг-лицерин (МГДГ) и трипальмитат в качестве стандартов (Гребенкина и др., 2012; Розенцвет и др., 2013).
Для определения мезоструктуры фотосинтетического аппарата растений, из средней части листьев вырезали высечки и фиксировали в 90% спирте, хранили при температуре -20ºС. Мезоструктуру фотосинтетического аппарата определяли согласно методики (Мокроносов, Борзенкова, 1978). Подсчет количества клеток в единице площади листа проводили в камере Горяева, подсчет количества хлоропластов в клетках мезофилла проводили на давленых препаратах согласно методу (Борзенкова, Храмцова, 2006).
Метанолиз жирных кислот (ЖК) осуществляли кипячением в 5 % -ном растворе HCl в метаноле. Полученные эфиры анализировали на хроматографе «Хроматэк Кристалл 5000.1» (Россия) в изотермическом режиме с использованием капиллярной колонки длиной 105 м и диаметром 0.25 мм «RESTEK» (США). Температура колонки – 180 о С, испарителя и детектора – 260 о С, скорость тока газа-носителя (гелий) – 2 мл/мин.
Статистическую обработку результатов анализов проводили с использованием программ Statistica 6.0 for Windows, Microsoft Excel 2007.
3. РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В настоящей работе исследованные растения представлены двумя родами – Oenothera и Epilobium . Растения рода Oenothera – O . biennis – произрастают по сухим разбитым пескам, вырубкам, обочинам песчаных дорог, боровым полянам. Распространению вида O . villosa способствовало разведение в садах. Сегодня этот вид можно встретить вдоль автомобильных дорог, на песчаных отмелях в долинах крупных рек, иногда также пойменные луга и заросли кустарников. Изредка произрастает на опушках сухих сосновых лесов. Внедряется во вторичные местообитания, но практически не проникает в нетронутые лесные или луговые сообщества. Являясь пионерным растением техногенных экотопов, вид закрепляет почву и, вероятно, уменьшает интенсивность эрозионного процесса (Черная Книга…, 2016). Оба вида данного рода являются двулетними травянистыми ксеромефитами, с одинаковым ареалом распространения (североамериканский) и генетическим статусом. Эти адвентивные виды относятся к сорным растениям.
Род Epilobium в нашей работе представлен травянистыми многолетниками. Растения C . angustifolium произрастают преимущественно на вырубках, прогалинах в хвойных и смешанных лесах, вблизи канав, на осушенных торфяниках, вдоль железнодорожных насыпей. Растения E . adenocaulon распространены в сообществах открытого типа, на илистых грунтах по берегам водоемов, на болотах, на полянах вдоль троп и лесных дорог, где уничтожен травяной покров. Встречаются также на различных вторичных, антропогенных местообитаниях: у дорог, канав, на железнодорожных откосах, чаще всего можно встретить в сообществах открытого типа, на илистых грунтах по берегам водоемов и в лесах (Скворцов, 1995; Скворцов,
1996). Растения E . hirsutum произрастают по берегам болот, рек.
Растения рода Epilobium различаются по отношению экологической группе, ареалу происхождения и генетическому статусу. Так, по режиму увлажнения C. angustifolium является мезофитом, E. adenocaulon – гигромезофитом, а E. hirsutum – гигрофитом. Растения представляют три разных ареала распространения: голарктический бореально-неморальный, североамериканский, Европейско-западноазиатский плюро-зональный, соответственно. По генетическому статусу C. angustifolium и E. hirsutum являются аборигенными видами, а E. adenocaulon – адвентивным. Все исследованные виды существенно различались по содержанию влаги в надземной части растений. Оводненность листьев растений варьировала в интервале 65,6– 80,2% от сырой массы. У ксеромезофитов, гигромезофитов и гигрофитов, произраставших в более влажных условиях, содержание влаги было примерно равным (72,8–77,1%). Меньше влаги содержали листья мезофитного растения C. angustifolium (65,6%).
Данные мезоструктурного анализа показали, что растений O . biennis имели самое большое число клеток в расчете на единицу площади листа (табл. 1). Объем и площадь поверхности клеток палисадной ткани у растения рода Oenothera были более чем 2 раза выше, чем у растений рода Epilobium . При этом число хлоропластов и их объем в растениях разных родов также различались, но в меньшей степени – не более 30% (27,2–32,5 и 20,0–24,1 тыс. мкм 3 ). Наиболее существенные различия между растениями отмечены по числу хлоропластов на единицу площади листа. Так у растений O . biennis данный показатель был в 8–10 раз больше по сравнению с другими видами. Это способствовало большему клеточному объему хлоропласта (КОХ), а также большим значениям индекса мембран клеток и индекса мембран хлоропластов.
Таблица 1
Параметры мезоструктуры листьев растений родов Epilobium и Oenothera
|
Параметры |
O. biennis |
O. villosa |
C. angustifo-lium |
E . hirsutum |
E . adenocau-lon |
|
Число клеток в единице площади (S) листа тыс. 2 см |
980,8±8,9 |
121,6±7,8 |
160,0±2,6 |
166,6±4,8 |
345,5±4,5 |
|
Объем клетки П) тыс. 3 мкм |
14,3±0,3 |
13,7±1,7 |
8,9±0,9 |
4,1±2,0 |
6,2±1,2 |
|
Площадь поверхности клетки П тыс. мкм2 |
3,6±0,6 |
3,5±0,3 |
2,8±0,2 |
1,5±0,5 |
1,9±0,9 |
|
Число ХЛ в клетке тыс. 2 см |
31,4±1,4 |
35,8±1,8 |
26,1±1,6 |
23,5±1,5 |
26,1±1,6 |
|
Объем ХЛ тыс. мкм3 |
27,2±3,0 |
32,5±2,0 |
24,1±2,1 |
23,2±3,2 |
20,0±4,2 |
|
Площадь поверхности ХЛ тыс. мкм2 |
45,2±4,5 |
50,0±2,0 |
41,2±6,0 |
39,7±3,5 |
48,0±3,6 |
|
Число ХЛ в единице S 2 листа млн см |
30,8±3,0 |
4,4±3,1 |
4,2±0,2 |
3,9±0,9 |
9,0±1,2 |
|
КОХ П, мкм3 |
455,4±20,0 |
382,7±4,8 |
341,0±5,0 |
174,5±1,7 |
237,5±3,7 |
|
Индекс мембран клеток П см2/см2 |
35,3±3,0 |
4,3±1,3 |
4,5±0,5 |
2,5±0,5 |
6,6±1,2 |
|
Индекс мембран ХЛ П см2/см2 |
13,9±2,0 |
2,2±1,0 |
1,7±0,7 |
1,6±0,6 |
4,3±2,9 |
Примечание: П – палисадная ткань; ХЛ – хлоропласты; N – число хлоропластов; S – площадь листа; КОХ – клеточный объем хлоропласта.
Известно, что размеры клеток мезофилла, включая палисадную и губчатую ткани, имеют важное значение для интенсивности физиологических процессов листа, поскольку они определяют поверхностно-объемное отношение мезофилла, плотность упаковки тканей, величину межклеточных воздушных пространств и другие параметры важные для диффузии газов и распределения света внутри листа (Иванова, 2012). В частности, у древесных растений наблюдаются мелкие размеры клеток, обуславливающих более плотную упаковку мезофилла, меньший объем воздухоносных межклетников (Pyankov, et al., 1999). Для травянистых растений той же экологической группы (мезофиты), что и древесные растения, характерны большие размеры клеток мезофилла, что позволяет им повысить скорость роста листа растяжением клеток. Травянистые растений сем. Onagracea характеризуются индивидуальными особенно- стями мезоструктуры, что, по-видимому, связано со специализацией видов по эколого-ценотическому и генетическому статусу.
Анализ пигментного фонда показал, что содержание зеленых и желтых пигментов варьировало в широких пределах (2,9–13,1 для хлорофиллов и 0,8–2,6 для каротиноидов мг/г в расчете на сухую массу) (рис. 1). В листьях растений рода Oenothera суммарное содержа-
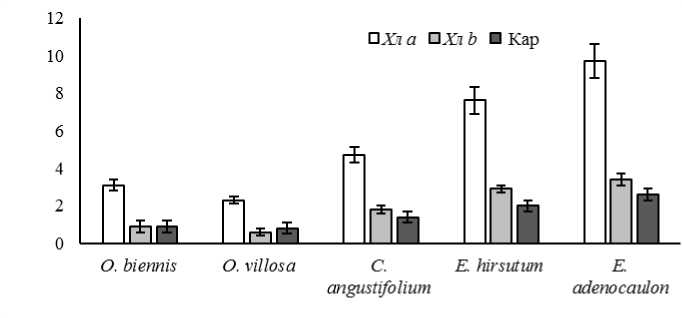
Рис. 1. Содержание хлорофиллов и каротиноидов в листьях растений родов Epilobium и Oenothera (мг/г сухой массы)
ние пигментов было меньше по сравнению с растениями рода Epilobium в 2 и более раз, несмотря на то, что их клетки были более объемными, содержали больше хлоропластов. Со- держание пигментов менялось в ряду ксероме-зофиты < мезофиты < гигромезофиты < гигрофиты, что дает основание предположить зависимость содержания пигментов от экологического статуса растений, связанного с режимом увлажнения. Однако различий между аборигенными и адвентивными растениями по данному показателю не выявлено.
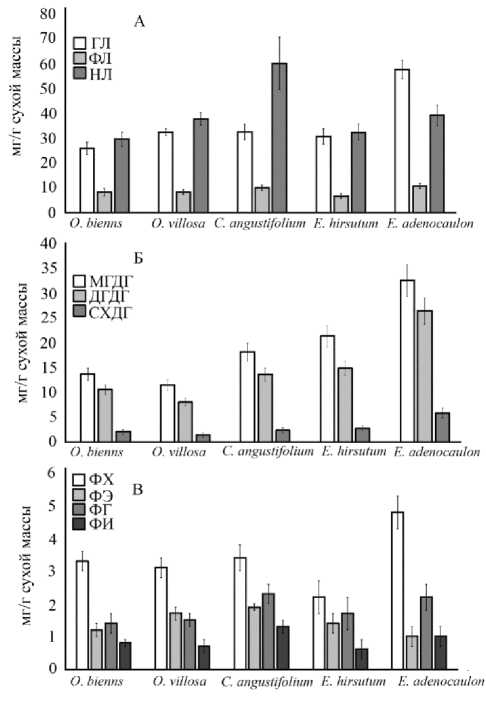
Рис. 2. Содержание суммарных и индивидуальных липидов в листьях растений родов
Epilobium и Oenothera
Соединения липидной природы и их надмолекулярным клеточным образованиям – биологическим мембранам – отводят важнейшую роль в функционировании основных клеточных биохимических механизмов. Данные механизмы определяют и регулируют физическое состояние клетки, ее взаимодействие, как с соседними клетками, так и внешней средой (Rozentsvet et al., 2016).
Среди суммарных липидов доля полярных липидов, отвечающих структуру мембран – гликолипидов (ГЛ) и фосфолипидов (ФЛ), составляет 25,4–62,1% и 7,6–12,7% от суммы липидов, соответственно. Наибольшее количество полярных липидов отмечено в растениях E . ad-enocaulon (Рис. 2 А). Кроме указанных липидов, в состав мембран входят компоненты, относящиеся к стериновой группе. Общее содержание стеринов варьировало от 1,3 до 3,1 мг/г сухой массы, что составляло от суммы мембранных липидов не более 5%.
Внутренние мембраны хлоропластов растений содержат моногалактозилдиацилглицерин (МГДГ), дигалактозилдиацилглицерин (ДГДГ) и сульфохиновозилдиацилглицерин (СХДГ).
Кроме указанных липидов, в состав мембран хлоропластов, входит единственный ФЛ фос-фатидилглицерин (ФГ). Для всех исследованных видов обнаружено классическое соотношение компонентов ГЛ. Так в листьях растений относительное содержание МГДГ составляло около 50–60 %, ДГДГ – 30–40%, а СХДГ 5–12% от суммы ГЛ (Dörmann, Benning 2002; Sakurai et al., 2007) (Рис. 2 Б). Вклад ФГ в пул ГЛ составлял не более 5% от суммы ГЛ. Сравнивая данные между родами можно видеть, что листья растений рода Epilobium, а именно E. hir-sutum и E. adenocaulon, содержали в 1,5–2,0 раза больше ГЛ, чем растения рода Oenothera. Можно также отметить четкую корреляцию между содержанием липидов в мембранах хлоропластов и содержанием фотосинтетических пигментов (r=0,9 при P < 0.04). Содержание и тех и других компонентов увеличивается в ряду ксеромезофиты – мезофиты –гигромезофиты – гигрофиты. В качественном составе липидов, представленных фосфатидилхолином (ФХ), фосфатидилэтаноламином (ФЭ), фосфатидилг-лицерином (ФГ), фосфатидилинозитом (ФИ) и отвечающих за структуру непластидных мембран, также не выявлено каких-либо особенностей в исследованных видах. В количественном отношении показано большее содержание ФХ у растений E. adenocaulon (Рис. 2 В).
Физические свойства мембран в большой степени определяются уровнем ненасыщенности ЖК. Среди ЖК исследованных растений было идентифицировано 14 компонентов (табл. 2). Как и предполагалось, на долю кислот с длиной углеводородной цепи 16 и 18 атомов углерода приходилось 90–98% от суммы ЖК. В составе насыщенных ЖК преобладает пальмитиновая кислота С 16:0. В листьях исследованных растений ее содержание составляло от 15,7 до 20,6% от суммы ЖК. Среди ненасыщенных ЖК доминирует линоленовая кислота (С 18:3n3), за ней следуют линолевая и олеиновая кислоты (С 18:2n6 и С 18:1n9). Содержание С 18:3n3 у растений E . hirsutum и E. adenocaulon было выше ~на 30%, по сравнению с другми видами. Индекс ненасыщенности (ИН) ЖК варьировал в интервале значений 1,8–2,3 и был больше у растений E . hirsutum и E . adenocaulon .
Таблица 2
Жирнокислотный состав листьев растений родов Epilobium и Oenothera
|
Кислоты |
O. biennis |
O. villosa |
C. angustifolium |
E. hirsutum |
E. adenocaulon |
|
С 14:0 |
1,8±0,8 |
1,4±0,2 |
2,9±0,4 |
1,1±0,2 |
1,1±0,1 |
|
С 16:0 |
18,9±0,9 |
18,3±0,1 |
20,6±0,3 |
15,9±0,5 |
15,7±0,1 |
|
С16:1 |
2,3±0,3 |
2,9±0,6 |
1,7±0,4 |
2,9±0,5 |
2,6±0,4 |
|
С18:0 |
2,1±0,1 |
2,4±0,2 |
2,5±0,2 |
2,6±0,3 |
2,0±0,2 |
|
С 18:1n9 |
2,4±0,3 |
1,7±0,4 |
3,1±0,1 |
1,1±0,1 |
1,2±0,1 |
|
С 18:2n6 |
19,0±0,2 |
16,6±0,3 |
12,9±0,5 |
7,9±0,5 |
6,7±0,5 |
|
С 18:3n3 |
51,8±0,4 |
54,7±0,4 |
51,6±0,3 |
65,7±0,4 |
69,7±0,4 |
|
С 20:0 |
0,9±0,3 |
0,8±0,4 |
2,5±0,2 |
1,5±0,2 |
1,0±0,1 |
|
Другие кислоты |
0,4±0,2 |
0,6±0,2 |
1,9±0,6 |
0,3±0,1 |
0,5±0,1 |
|
Индекс ненасыщенности |
1,9 |
2,0 |
1,8 |
2,2 |
2,3 |
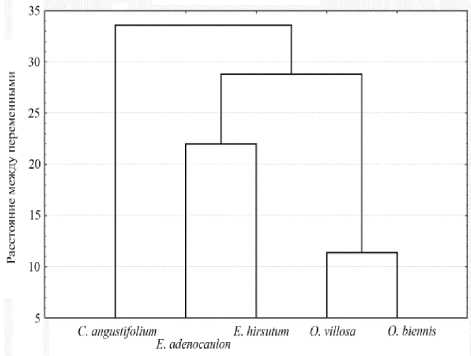
Рис. 3. Степень родства биохимических признаков растений родов Epilobium и Oenothera по данным кластерного анализа
Следует отметить, что Е. adenocaidon может превосходит по численности многие другие виды, заселяющие пионерные местообитания – Tussilago farfara , Chamaenerion angustifolium и Epilobium montanum (Myerscough, Whitehead, 1966) благодаря способности прорастать при низких (10°С) температурах, низком уровне освещения, а также в засушливых условиях. Не исключено, что эти свойства обеспечиваются большей эффективностью фотосинтеза за счет большего содержания фотосинтетических пигментов, локализованных в мембранах хлоропластов, а также более гибкими мембранами, обеспечивающими большую мобильность функциональных комплексов фотосинтетического аппарата за счет большей ненасыщенности ЖК.
Кроме измерения структурных и биохимических параметров, была предпринята попытка установить взаимосвязь между исследованными биохимическими параметрами и генетическим статусом видов (рис. 3). Это позволяет оценить экологический потенциал растений, способность адаптироваться в разнообразных условиях.
Для кластерного анализа были использованы все измеренные биохимические показатели, такие как содержание зеленных и желтых пигментов, а также содержание отдельных компонентов мембранных и запасных липидов. По данным рис. 3 можно видеть, что наибольшей геометрической близостью характеризовались растения рода Oenothera, в листьях которых было отмечено наименьшее содержание пигментов и липидов. Близость результатов между данными видами связана с одинаковой эколого-ценотической приуроченностью, к тому же оба вида относятся к адвентивным растениям. В следующем кластере объединены растения видов E. hirsutum и E. adenocaulon, несмотря на то, что они имеют разный генетический статус. На дендрограмме положение вида C. angustifo-lium находится между кластерами растений Oenothera и Epilobium.
Полученные данные позволяют заключить, что параметры мезоструктуры определяются видовыми особенностями, в то время как биохимические показатели имеют определенное межвидовое сходство. На основании измеренных показателей растений сем. Onagraceae, каких-либо особенностей, характеризующих растения как представителей адвентивной или аборигенной флоры, не выявлено.
Список литературы Особенности мезоструктуры, пигментного и липидного состава представителей сем. Onagraceae
- Асминг С.В. Морфолого-биологические особенности и экология видов рода Epilobium L. // Вестн. ОГУ. 2007. № 10. С. 128-134.
- Борзенкова Р.А., Храмцова Е.В. Определение мезоструктурных характеристик фотосинтетического аппарата растений. Екатеринбург: Изд-во Уральского ун-та. 2006. 25 с.
- Гребенкина Т.М., Нестеров В.Н., Розенцвет О.А., Богданова Е.С. Изменение состава липидов и пигментов Plantago media (Plantaginaceae) в течение светлого времени суток // Растит. ресурсы. 2012. Вып. 4. С. 565-578.
- Кейтс М. Техника липидологии. М.: Мир, 1975. 323 с.
- Мокроносов А.Т., Борзенкова Р.А. Методика количественной оценки и функциональной активности фотосинтезирующих тканей и органов // Тр. по прикладной ботанике, генетике и селекции ВНИИ растениеводства. Л., 1978. Т. 61, вып. 3. С. 119.
