Особенности микроморфологии и анатомии листа Populus tremula L. (sect. Populus) и P. nigra L. (sect. Aigeiros duby, Salicaceae)
 и P. nigra L. (sect. Aigeiros duby, Salicaceae)")
Автор: Молганова Наталья Александровна, Овеснов Сергей Александрович
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2022 года.
Бесплатный доступ
Изучена микроморфология и анатомия, эпидермы абаксиальной поверхности листа Populus tremula L. (sect. Populus) и P. nigra L. (sect. Aigeiros Duby, Salicaceae) с помощью светового и сканирующего электронного микроскопов. Основные клетки эпидермы P. nigra по Н.А. Анели криволинейные кривостеночные; коэффициент извилистости антиклинальных стенок - 14.3±0.3 мкм/мкм2; площадь от 437.5 до 1300.0 мкм2 (среднее 820.9±39.9); P. tremula - извилистые, близкие к измятостеночным; 23.2±0.4 мкм/мкм2; от 187.5 до 612.5 мкм2 (среднее 351.7±12.6). Замыкающие клетки устьиц у обоих видов чечевицевидные, разнонаправленные, располагаются на одном уровне с основными. У P. tremula довольно мелкие (их длина 17.9±0.4 мкм), а у P. nigra они крупнее (24.6±0.4 мкм). Плотность устьиц у P. tremula колебалась от 333 до 571 шт./ мм2, у P. nigra - от 100 до 183 шт./ мм2. Около устьиц имеется складчатый (стриатный) микрорельеф. У P. nigra прямые или извилистые складки от 5 до 25 шт. расходятся в разные стороны на расстояние от 10 до 27 мкм. Обнаружены три типа микрорельефа. У P. tremula небольшое число околоустьичных складок (от 3 до 17 шт.) расходились перпендикулярно устьичной щели в разные стороны на расстояние ~ 10-20 мкм.
Populus tremula, populus nigra, микроморфология листа, анатомия листа
Короткий адрес: https://sciup.org/147238656
IDR: 147238656 | УДК: 581.6 | DOI: 10.17072/1994-9952-2022-2-93-100
Features of the micromorphology and anatomy of the leaf Populus tremula L. (sect. Populus) and P. nigra L. (sect. Aigeiros duby, Salicaceae)
The micromorphology and anatomy of the epidermis of the abaxial surface of the leaf of Populus tremula L. (sect. Populus) and P. nigra L. (sect. Aigeiros Duby, Salicaceae) using light and scanning electron microscopes. The main cells of the epidermis of P. nigra according to N.A. Aneli are curvilinear curvilinear; the coefficient of tortuosity of the anticline walls is 14.3±0.3 μm/mm2; the area is from 437.5 to 1300.0 mm2 (average 820.9±39.9); P. tremula - sinuous, close to indented; 23.2± 0.4 μm/mm2; from 187.5 to 612.5 mm2 (average 351.7±12.6). The closing cells of the stomata in both species are lentil-shaped, multidirectional, located on the same level with the main ones. In P. tremula they are rather small (their length is 17.9± 0.4 microns), and in P. nigra they are larger (24.6± 0.4 microns). Stomata density in P. tremula ranged from 333 to 571 pcs/mm2, in P. nigra - from 100 to 183 pcs/ mm2. There is a folded (striate) microrelief near the stomata. In P. nigra, straight or sinuous folds ranging from 5 to 25 pieces diverge in different directions at a distance of 10 to 27 microns. Three types of microrelief were found. At P. tremula a small number of periosteal folds (from 3 to 17 pcs.) diverged perpendicular to the stomatal slit in different directions at a distance of ~ 10-20 microns.
Текст научной статьи Особенности микроморфологии и анатомии листа Populus tremula L. (sect. Populus) и P. nigra L. (sect. Aigeiros duby, Salicaceae)
В научной литературе систематически появляются разнообразные работы, посвященные исследованиям видов и (чаще) культиваров из рода Populus ( Salicaceae ) [Этапы формирования…, 2005; Скворцов, 2010; Zeng et al., 2016; Евлаков, Царев, Заплетин, 2017; Куприянова, 2020 и др.]. Однако вопросы, связанные с идентификацией и концепцией видов данного рода решены еще не полностью. Таким образом, остается продолжать попытки получения данных о морфологических, анатомических особенностях видов и гибридогенных таксонов рода Populus на региональном материале. В 2021 г. изучена микроморфология и анатомия аборигенных видов: Populus tremula L . (sect. Populus ) и P. nigra L. (sect. Aigeiros Duby) в Пермском крае.
-
P. tremula отмечена в Восточной и Западной Европе, в Крыму, на Кавказе, в Сибири, на Дальнем Востоке, на севере п-ова Корея. Ее ареал простирается от лесотундры до степей, на севере заходит за полярный круг [Свиязева, 1977с; Скворцов, 2010]. В Пермском крае P. tremula встречается часто, широко представлена в древостоях различных типов леса и синтаксонах, образует осинники [Овеснов, 2007].
-
P. nigra распространена в Западной и Восточной Европе, на Кавказе, в Малой Азии, Иране, Северной Африке, Восточной Сибири, доходя до бассейна среднего Енисея, в южной части Западной Сибири, на севере Казахстана [Свиязева, 1977b; Скворцов, 2010]. В Пермском крае произрастание данного вида ограничено в природе поймами и долинами рек [Овеснов, 2007]. Он произрастает одиночно, значительных массивов, которые можно рассматривать как лесонасаждения, не образует.
В естественных условиях в Пермском крае отмечается еще один вид – P. alba L. (sect. Populus ), встречающийся очень редко на юго-западе региона [Овеснов, 2007]. Его ареал охватывает Западную Европу, среднюю и южную части Восточной Европы, Крым, Кавказ, южную часть Западной Сибири, Казахстан, Среднюю и Малую Азию, Иран, Афганистан, Западный Китай, Гималаи, северо-западную Африку [Сви-язева, 1977a; Скворцов, 2010]. По литературным данным P. alba [Скворцов, 2010; Zeng et al., 2016 и мн. др.] в местах совместного произрастания с P. tremula повсеместно скрещиваются, образуя гибридогенный таксон P. × canescens (Ait.) Smith. Идентификацию родительских видов и нотовидов осложняет интрогрессивная гибридизация [Fossati et al., 2004; Lexer et al., 2005]. Локальные популяции P. alba Пермского края находятся на севере ареала, поэтому менее стабильны, в связи с чем не изучались.
Материал и методы
В рамках нашего исследования были изучены деревья P. tremula одной локальной популяции в долине р. Уинка в черте г. Перми. У P. nigra были исследованы 4 локальные популяции: в долине р. Его-шиха, на Липовой горе в г. Перми; а также в Кунгурском р-не Пермского края около д. Захаровка на склоне к р. Ирень и в районе Спасской горы. Для данного исследования были отобраны листья со вполне развитых брахибластов, с хорошо освещенной нижней части кроны, на высоте 1.5–2.5 м. С нефиксированных листьев в центральной части с их абаксиальной стороны между главной жилкой и краем листа была снята эпидерма, по общепринятым методикам [Анели, 1975] приготовлены временные препараты и изучены в проходящем свете на белом поле под микроскопом Биомед-5. Часть листьев была высушена, из них вырезаны круглые фрагменты диаметром около 1 см. Они изучены с абаксиальной стороны с помощью сканирующего электронного микроскопа Hitachi ТМ3000 при ускоряющем напряжении 15 кВ в режиме обратных рассеивающихся электронов (BSE). Яркость и контрастность полученных микрофотографий были изменены в растровом графическом редакторе Adobe Photoshop. Для определения количественных параметров клеток с помощью средств векторного графического редактора CorelDraw были измерены периметры, длины и площади клеток. Для получения статистических данных было измерено основных клеток эпидермы у P. tremula – 58, а у P. nigra – 253, устьичных аппаратов у P. tremula – 24, а у P. nigra – 185. Статистическая обработка проведена общепринятыми методами [Доспехов, 1985] с помощью программы Microsoft Office Excel 2010.
Результаты и их обсуждение
Основные клетки эпидермы на изученных препаратах P. tremula были мелкими, что по данным А.А. Паутова [1996] характерно для всей совокупности видов P. tremula L., P. davidiana Dode, P. grandidentata Michaux, P. sieboldii Miq., P . tremuloides Michaux, которые формируют секцию Trepidae Dode [по Р.В. Камелину, 1973]. Средняя площадь основных клеток эпидермы 351.7±12.6 мкм2. Отдельные клетки на препарате варьировали по площади от 187.5 до 612.5 мкм2. Коэффициент вариации данного признака высокий 27.4% [по А.А. Паутову, 2011]. Периметр клеток 78.9±1.5 мкм. Коэффициент вариации периметра – средний – 14.9%.
У P. nigra основные клетки эпидермы на изученных препаратах были крупнее и несколько больше колебались по площади от 437.5 до 1300.0 мкм2, в среднем 820.9±39.9 мкм2. Периметр 113.7±2.4 мкм. Коэффициенты вариации данных признаков несколько выше, чем у P. tremula – 37.0 и 15.8% соответственно.
Изученные микрофотографии позволили оценить форму основных клеток эпидермы, исходя из классификации Н.А. Анели [1975]. У P. nigra они оказались криволинейными кривостеночными. На некоторых сторонах антиклинальных стенок у части клеток есть одна вершина, неглубоко вклинивающаяся в соседнюю клетку. Иногда эти вершины совершенно не выражены (рис. 1).
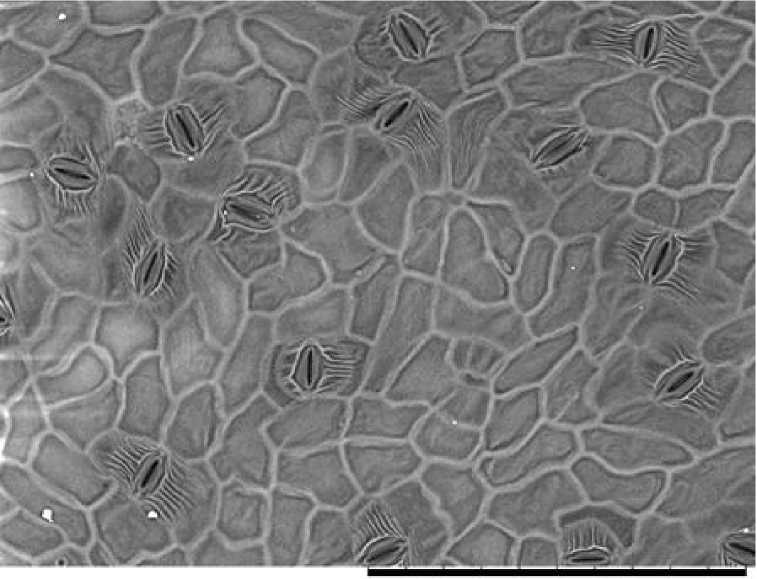
Рис. 1. Микроскульптура абаксиальной сторона листа P. nigra , Кунгурский район Пермского края, д. Захаровка
[Microsculpture of the abaxial side of the P. nigra leaf, Kungursky district of Perm Krai, Zakharovka village:]
У P. tremula клетки оказались извилистыми (рис. 2), более близкими к измятостеночным по Н.А. Анели [1975]. Средний коэффициент извилистости антиклинальных стенок клеток изученных листьев P. tremula равнялся 23.2±0.4 мкм/мкм2, а у P. nigra – 14.3±0.3 мкм/мкм2.
У ранее изученных авторами листьев брахибластов культиваров P. laurifolia с абаксиальной стороны в г. Перми клетки были более извилистыми (рис. 3), измятостеночными. Средний коэффициент извилистости равен 23.7 мкм/мкм2 [Молганова, Овеснов, 2021].
Виды рода Populus имеют неодинаковые устьица, так, например, P. tremula характеризуется парацит-ными, латероцитными или аномоцитными типами [Паутов, 2009; Куприянова, 2020]. Соответственно, данный признак не пригоден для идентификации видов.
На исследованных препаратах плотность устьиц у P. tremula колебалась от 333 до 571 шт./мм2. Из данных литературы известно, что плотность и размер устьиц зависит у листьев Populus от экологических условий произрастания [Pearce et al., 2006; Di Baccio, Minocci, Sebastiani, 2010; Евлаков, Царев, Заплетин, 2017 и др.] и от развития (мощности) деревьев [Этапы формирования…, 2005]. В исследованиях А.А.
Паутова с соавторами число устьиц на абаксиальной стороне листьев P. tremula было 255–352 шт./мм2 [Этапы формирования…, 2005]. У P. nigra в г. Перми и на юге Пермского края плотность устьиц значительно ниже – от 100 до 183 шт./ мм2. По литературным данным [Евлаков, Царев, Заплетин, 2017], у культивара ‘Регенерата’ ( P. nigra х P. deltoides Bartram ex Marshall) плотность устьиц около 140 шт./мм2, а ‘Пионер’ ( P. nigra ) – немногим более 100 шт./мм2.
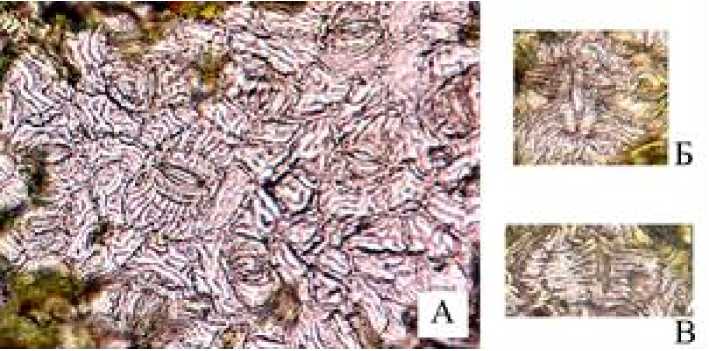
Рис. 2. Абаксиальная сторона листа P. tremula :
А – участок с основными клетками эпидермы, Б, В – околоустьичная складчатость
[The abaxial side of the P. tremula leaf:
A – the area with the main cells of the epidermis, B, C – parotid folding
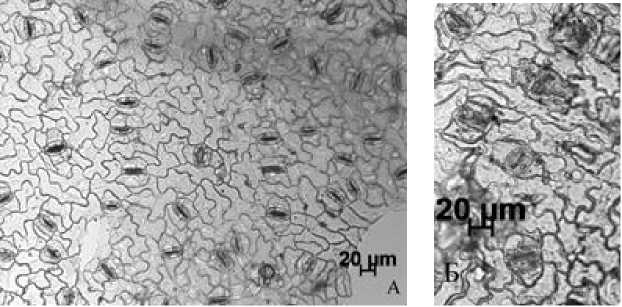
Рис. 3. Абаксиальная сторона листа P. laurifolia : А – на ауксибластах, Б – на брахибластах
[Abaxial side of P. laurifolia leaf:
A – on auxiblasts, B – on brachiblasts]
Замыкающие клетки устьиц чечевицевидные, разнонаправленные, располагаются на одном уровне с основными. Замыкающие клетки устьиц P. tremula довольно мелкие (их длина 17.9±0.4 мкм), а у P. nigra они крупнее (24.6±0.4 мкм). У P. laurifolia длина замыкающих клеток составила 28.6±0.5 мкм [Молгано-ва, Овеснов, 2021]. Кривая распределения длины замыкающих клеток устьиц почти правильная, имеет характер, близкий к симметричному и одновершинному (рис. 4). Устьиц, значительно отличающихся по величине, описанных в литературе [Sitholey, Pandey, 1971; Stomata morphological…, 2015], на изученных препаратах не было обнаружено.
У изученных видов на абаксиальной стороне листа около устьиц имеется складчатый (стриатный) микрорельеф, который формирует так называемую околоустьичную кутикулярную складчатость. Особенности микрорельефа являются видоспецифичными и, следовательно, значимыми для таксономической диагностики [Stace, 1984; Сапач, 2005]. Их можно использовать совокупно с другими признаками [Dehgan, 1980; Olowokudejo, 1993; цит. по: Паутов и др., 2014]. В листе P. nigra выделяются устьица с тремя типами микрорельефа (рис. 1; 5, А). У большей части складки расходятся в противоположные стороны на расстояние от 10 до 27 мкм (средняя длина складок 17.8±4.7 мкм). Число складок от 5 до 25 шт. Иногда складки могут быть прямыми, а иногда извилистыми. В некоторых случаях они отходят перпен- дикулярно устьичной щели, а иногда на микрофотографиях заметны отклонения складок в сторону. Исходя из литературных данных [Паутов, 2011], у Hydrangea macrophylla (Thunb.) Ser. число и длина складок зависит от размера устьиц и величины их побочных клеток. У P. nigra некоторая зависимость наблюдалась между длиной устьиц и числом складок (коэффициент корреляции средний положительный 0.4). Между длиной устьиц и длиной складок зависимости не выявлено (r 0.05).
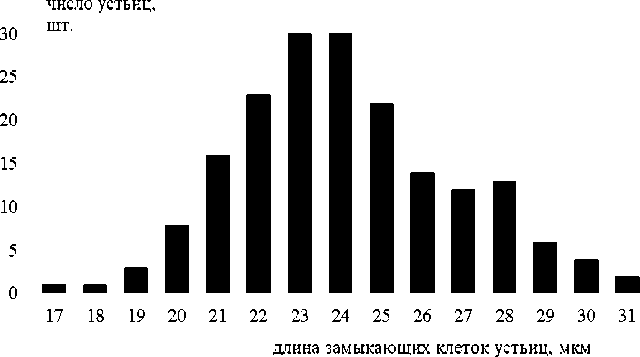
Рис. 4. Распределение числа устьиц по длине замыкающих клеток на абаксиальной стороне листа у P. nigra
[Distribution of the number of stomata along the length of the closing cells on the abaxial side of the leaf in P. nigra ]
Редко у некоторых устьиц складки расходились радиально. Число складок при этом было большим – от 13 до 30. Они простирались в разные стороны на ~17–49 мкм. У приблизительно пятой части устьиц, попадающих в поле зрения, складки были почти не заметны или от устьиц отходили по 4 недлинные складки (от 5 до 12 мкм) от полюсов устьичных клеток.
У P. tremula небольшое число околоустьичных складок (от 3 до 17 шт.) расходились перпендикулярно устьичной щели в противоположные стороны на расстояние ~10–20 мкм. У части устьиц складки не были заметны (рис. 5, Б).
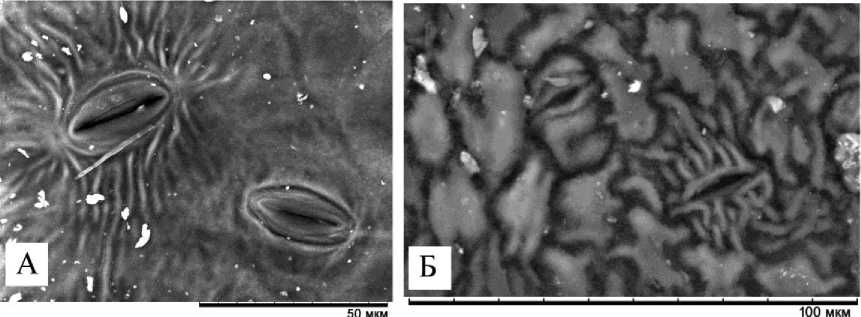
Рис. 5. Типы околоустьичных складок: А – P. nigra ; Б – P. tremula [Types of periarticular folds: А – P. nigra ; Б – P. tremula ]
Заключение
Основные клетки эпидермы у P. tremula и P. laurifolia сходны между собой по степени извилистости больше, чем с таковыми у P. nigra. На изученных микропрепаратах P. nigra и P. tremula околоустьичная складчатость формировалось по-разному. Клетки на препаратах P. nigra с деревьев с территории г. Перми и с юга Пермского края по периметрам основных клеток эпидермы, коэффициентам извилистости антиклинальных поверхностей, длине замыкающих клеток устьиц, характеру и длине околоустьичных складок достоверно не различались.
Список литературы Особенности микроморфологии и анатомии листа Populus tremula L. (sect. Populus) и P. nigra L. (sect. Aigeiros duby, Salicaceae)
- Анели Н.А. Атлас эпидермы листа. Тбилиси: Мецниереба, 1975. 112 с.
- Доспехов Б.А. Методика полевого опыта (с основами статистической обработки результатов исследований). М.: Агропромиздат, 1985. 352 с.
- Евлаков П.М., Царев А.П., Заплетин В.Ю. Изучение фотосинтетических особенностей и интенсивности транспирации у различных сортов и клонов тополя (Populus L.) // Труды Санкт-Петербургского научно-исследовательского института лесного хозяйства. 2017. № 4. С. 4-13.
- Камелин Р.В. Флорогенетический анализ естественной флоры горной Средней Азии. Л: Наука, 1973. 357 с.
- Куприянова Е.А. Сравнительное фармакогностическое исследование представителей рода тополь (Populus L.): автореф. дис. ... канд. фарм. наук. Самара, 2020. 18 с.
- Молганова Н.А., Овеснов С.А. Особенности микроморфологии и анатомии листа Populus laurifolia Ledeb. // Агротехнологии XXI века: стратегия развития, технологии и инновации: материалы Всерос. науч.-практ. конф. Пермь, 2021. С. 499-503.
- Овеснов С.А. Сем. Salicaceae - Ивовые // Иллюстрированный определитель растений Пермского края. Пермь: Книжный мир, 2007. С. 279-303.
- Паутов А.А. Закономерности филоморфогенеза вегетативных органов растений. СПб.: Изд-во СПбГУ, 2009. 219 с.
- Паутов А.А. Основные направления и закономерности преобразования структуры листа в эволюции тополей: автореф. дис. ... д-ра биол. наук. СПб., 1996. 45 с.
- Паутов А.А. Расположение складок микрорельефа на побочных клетках устьиц Hydrangea macro-phylla (Thunb.) Ser. (Hydrangeaceae) // Вестник Санкт-Петербургского университета. Сер. 3. Биология. 2011. № 2. С. 39-44.
- Паутов А.А. и др. Микрорельеф поверхности листьев цветковых растений: устьичные кольца и выступы // Ботанический журнал. 2014. Т. 99. № 6. С. 625-640.
- Сапач Ю.О. К вопросу о постоянстве микрорельефа поверхности листа цветковых растений // Герценовские чтения: материалы межвуз. конф. СПб., 2005. С. 30-31.
- Свиязева О.А. Populus alba L. - Тополь белый, или серебристый // Ареалы деревьев и кустарников СССР. Л.: Наука, 1977a. Т. 1. С. 77.
- Свиязева О.А. Populus nigra L. - Тополь черный, или осокорь // Ареалы деревьев и кустарников СССР. Л.: Наука, 1977b. Т. 1. С. 80-81.
- Свиязева О.А. Populus tremula L. - Осина // Ареалы деревьев и кустарников СССР. Л.: Наука, 1977с. Т. 1. С. 84-85.
- Скворцов А.К. Систематический конспект рода Populus в Восточной Европе, Северной и Средней Азии // Бюллетень Главного ботанического сада. 2010. № 196. С. 62-73.
- Этапы формирования складчатого микрорельефа поверхности листа Populus tremula L. (Salicaceae Mirb.) // Вестник Санкт-Петербургского университета. Сер. 3. Биология. 2005. № 4. С. 3-8.
- Di Baccio D., Minocci A., Sebastiani L. Leaf structural modifications in Populus x euramericana subjected to Zn excess // Biologia Plantarum. 2010. Vol. 54 (3). P. 502-508. doi: 10.1007/s1053 5-010-0088-x.
- Fossati T. et al. Development of molecular markers to assess the level of introgression of Populus tremula into P. alba natural populations // Plant Breed. 2004. Vol. 123. P. 382-385.
- Lexer C. et al. Barrier to gene ow between two ecologically divergent Populus species, P. alba (white poplar) and P. tremula (European aspen): the role of ecology and life history in gene introgression // Molecul. Ecol. 2005. Vol. 14. P. 1045-1057.
- Pearce D.W. et al. Stomatal characteristics of riparian poplar species in a semi-arid environment // Tree Physiology. 2006. Vol. 26 (2). P. 211-218. doi: 10.1093/treephys/26.2. 211.
- Sitholey R.V., Pandey Y.N. Giant stomata // Annals of Botany. 1971. Vol. 35. P. 641 -642. URL: http://aob.oxfordjournals.org/content/35/3/641.s hort.
- Stace C.A. The taxonomic importance of the leaf surface // Syst. Assoc. 1984. Spec. Vol., № 25. P. 67-94.
- Stomata morphological traits in two different genotypes of Populus nigra L. / G. Russo, P. De Angelis, J. Mickle & M. Lumaga // I Forest-Biogeosciences and Forestry. 2015. Vol. 8. P. 547-551. DOI: 10.3832/ifor1104-007.
- Zeng Y.F. et al. Genetic structure of Populus hybrid zone along the Irtysh River provides insight into plastid-nuclear incompatibility // Sci. Rep. 2016. Vol. 6. P. 28043. https://doi.org/10.1038/srep28043.
