Особенности микроспорогенеза у тетраплоидной линии кукурузы КРП-1

Автор: Лобанова Людмила Петровна, Колесова Алла Юрьевна, Шацкая Ольга Александровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-2 т.21, 2019 года.
Бесплатный доступ
Тетраплоидная линия кукурузы КрП-1 характеризуется многочисленными отклонениями в структуре зрелых пыльцевых зерен. Цель работы заключалась в изучении мужского мейоза у данной линии, особенности которого в значительной степени определяют качество пыльцы и успешность оплодотворения. У исследованных растений аномалии мейоза обнаружены на всех стадиях. Они представлены тремя основными типами: нарушениями кариокинеза и цитокинеза и образованием микроспороцитов атипичной формы. Выявлен сложный комплекс нарушений мейотических делений, включающий аномальное расхождение хромосом, неполный цитокинез или его отсутствие, изменение ориентации веретен деления и синхронизации делений в мета-анафазе. Выделены две группы аномалий, редко встречающиеся у кукурузы: частичный цитокинез в виде «насечек» или «тоннелей» и образование микроспороцитов атипичной формы. Последнее предполагает наличие специфической мутации, изменяющей форму мейотических клеток. На основе полученных структурных характеристик мейоза можно оценить потенциальную генотипическую, морфологическую и функциональную гетерогенность пыльцы. Многие обнаруженные нарушения карио- и цитокинеза в мейозе могут быть причиной частичной стерильности пыльцы или привести к образованию нередуцированных и анеуплоидных гамет. Однако высокий процент образования типичных микроспор свидетельствует о возможности использования тетраплоида КрП-1 в селекции в качестве донора диплоидных гамет.
Тетраплоидная кукуруза, микроспорогенез, кариокинез, ориентация веретен деления, трехполюсные веретена, выпадение цитокинеза, тоннельный цитокинез, атипичная форма микроспороцитов
Короткий адрес: https://sciup.org/148314170
IDR: 148314170 | УДК: 633.15
Features of microsporpogenesis in maize tetraploid line KRP-1
Tetraploid maize line KrP-1 is characterized by numerous deviations in the structure of mature pollen grains. The aim of the work was to study male meiosis in this line, the characteristics of which largely determine the quality of pollen and the success of fertilization. In the studied plants, anomalies of meiosis were found at all stages. They are represented by three main types: disorders of karyokinesis and cytokinesis and the formation of atypical microsporocytes. A complicated complex of disorders of meiotic divisions was revealed, including an abnormal chromosome discrepancy, incomplete cytokinesis or its absence, a change in the orientation of the division spindle and synchronization of divisions in meta-anaphase. Two groups of anomalies are rarely found in maize: partial cytokinesis in the form of “notches” or “tunnels” and the formation of atypical microsporocytes. The latter suggests the presence of a specific mutation that changes the shape of meiotic cells. Based on the obtained structural characteristics of meiosis, it is possible to assess the potential genotypic, morphological, and functional pollen heterogeneity. Many discovered violations of karyo-and cytokinesis in meiosis can be the cause of partial sterility of pollen or lead to the formation of unreduced and aneuploid gametes. However, a high percentage of formation of typical microspores indicates the possibility of using the KrP-1 tetraploid in breeding as diploid gamete donors.
Текст научной статьи Особенности микроспорогенеза у тетраплоидной линии кукурузы КРП-1
Одним из важных критериев, используемых при изучении системы размножения растений, является состояние их генеративной сферы. Изучение аномалий мейотического деления используется не только в области цитогенетики, но и для решения многих задач клеточной биологии и селекции. Прежде всего, это относится к исследованию разнообразных внутриклеточных процессов, определяющих качество мужских и женских гамет, следовательно, успешность оплодотворения и семенную продуктивность. Изучение влияния полиплоидии на проявление генеративных признаков представляет значительный интерес в связи с оценкой фертильности полиплоидов, их генетической изменчивостью и возможностью использования в практической селекции.
Изучение генеративных признаков тетра-плоидной линии кукурузы КрП-1 на протяжении нескольких лет проводились на кафедре генетики Саратовского государственного университета. Анализ качества пыльцы показал повышенную степень дегенерации и достаточно широкий спектр отклонений в ее структуре по сравнению с контрольными диплоидными линиями [1; 2; 3]. Разнообразные аномалии обнаружены также на стадии сформировавшихся спорад микроспор [4]. Все это делает актуальным исследование особенностей микроспорогенеза у тетраплоидных растений и определение его зависимости от уровня плоидности генома.
Известно, что в микроспорогенезе морфология и онтогенетическая судьба микроспор определяются двумя ключевыми процессами: правильностью расхождения хромосом и успешностью деления цитоплазмы. Нарушения при расхождении хромосом в мейозе ведут к изменению баланса генов, аномалиям развития микроспор и их плоидности [5; 6; 7]. Аномалии цитокинеза изменяют обособление дочерних клеток и пространственную ориентацию деления цитоплазмы, что также может приводить к изменению плоидности, дифференцировки и аномалиям развития клеток [8].
Цель работы заключалась в изучении особенностей карио- и цитокинеза в материнских клетках микроспор у тетраплоидных растений кукурузы линии КрП-1.
МАТЕРИАЛ И МЕТОДИКА
Материалом исследования послужили материнские клетки микроспор (микроспороциты) тетраплоидной линии кукурузы КрП-1 (Краснодарская популяция-1). Эта линия была получена в Краснодарском научно-исследовательском институте сельского хозяйства им. П.П. Лукьяненко и прошла многолетний отбор на повышение фертильности в Саратовском университете. Всего было проанализировано 9 растений. Пыльники фиксировали ацетоалкоголем (1:3). Для приготовления каждого препарата брали пыльники на стадии мейоза из одного цветка. С каждого растения анализировали 5-10 цветков и от 164 до 860 микроспороцитов. Анализ микроспорогенеза проводили на временных препаратах, окрашенных ацетокармином на микроскопе «AxioStar-plus» при увеличении 10×40. Всего было проанализировано 3909 микроспороцитов на разных стадиях мейоза. Фотографирование проводили при тех же увеличениях с помощью программы визуализации «Zoombrowser».
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализ показал, что у изученных растений отклонения от нормы присутствуют на всех стадиях мейоза. Стадии мейоза различаются специфичной морфологией нарушений и их количеством. Доля нарушений на каждой стадии определялась как процент от общего количества клеток, обнаруженных на данной стадии. Типы нарушений и их средняя частота в исследованной выборке растений представлены в таблице.
На стадии профазы-I большинство микроспороцитов имеют нормальное строение. Это крупные округлые клетки с густой цитоплазмой и крупными ядрами, в которых хорошо просматривается ядрышко и клубок хроматиновых нитей. В редких случаях обнаруживаются двуядерные микроспороциты, ядра которых находятся на некотором удалении друг от друга. Возможно, их образование связано с подавлением цитокинеза в предмейотических митозах археспо-риальных клеток. Вторым зарегистрированным типом аномалий на этой стадии были клетки атипичной формы, у которых длина в 2-3 раза превосходила ширину. Частота таких вытянутых клеток составила 0,5 %.
На стадии метафазы-I нарушения кариокинеза встречаются у 1,5 % микроспороцитов. Хромосомы при этом могут быть диффузно распределены по микроспороциту (рис. 1а) или от- дельные из них выброшены за пределы ахрома-тинового веретена (рис.1 б,в).
Основным нарушением на стадии анафазы-I является разброс хромосом по отчетливо видимому веретену деления (рис. 1 г,д ). Такой тип аномалий составил 1,9 % от общего числа проанализированных анафаз. Выбросы отдельных хромосом за пределы ахроматинового веретена встречались с частотой 0,5 %. С такой же частотой зарегистрировано образование микроспороцитов с мостом между группами хромосом (рис. 1 е ). Общее количество аномальных анафаз в первом мейотическом делении составило в среднем 3,0 %.
В телофазе-I количество аномальных картин деления возрастает до 12,1 %. Встречаются клетки с отставанием хромосом (рис. 1 ж,з ) и микроядрами (1,2 %), возникающими вследствие нарушений кариокинеза в анафазе-I. Доминируют на этой стадии микроспороциты с нарушенным цитокинезом, общая доля которых составляет 10,3 %. Обнаружено 3 группы нарушений цитокинеза: изогнутое положение клеточной пластинки (рис. 1 и ), неполный цитокинез в виде внутриклеточного «тоннеля» (рис. 1 к ) или «насечки» с внешней стороны (рис. 1 л ) и полное выпадение цитокинеза (рис. 1 м ). Такие нарушения являются причиной образования либо неравных по величине двух клеток, либо двуядерных монад.
Основным типом нарушений на стадии метафазы-II становятся асинхронные деления ядер (2,6 %). Чаще всего одно ядро находится на стадии метафазы-II, а второе – анафазы-II (рис. 2 а ). Количество клеток с аномальным положением хромосом (рис. 2 б-г) в метафазе-II составило 2,2 %, что несколько выше, чем на стадии метафазы-I.
Частота аномальных делений в анафазе-II увеличивается до 15,6 %, что почти в 4 раза больше, чем в анафазе первого мейоза. Увеличивается и спектр нарушений. Помимо выбросов хромосом за пределы веретена деления и отставания отдельных хромосом при их расхождении к полюсам (рис. 2 д ), были зарегистрированы трехполюсные веретена (рис. 2 е,ж ). Частота диад с трехполюсными веретенами в одной или обеих клетках достигает 1 %. В первом мейозе такая аномалия не обнаружена. Значительная часть нарушений представлена асинхронными делениями ядер в мета-анафазе или ана-те-лофазе (5,8 %) и/или нарушением ориентации веретен деления (рис. 2 а,д ). В последнем случае веретена располагаются перпендикулярно или под углом друг к другу, а не параллельно, как в норме у кукурузы.
Стадия телофазы-II характеризуется наличием в микроспороцитах микроядер (2,2 %) или, напротив, слившихся ядер (рис. 2 и,к ), а так-
Таблица. Типы и частота нарушений мужского мейоза у тетраплоидных растений
|
Стадии мейоза |
Число проанализированных микроспороцитов, шт. |
Общее количество нарушений, % |
Тип нарушений |
Частота типов нарушений, % |
|
Профаза-I |
1747 |
1,1 |
Двуядерные клетки |
0,6 |
|
Атипичная форма клетки |
0,5 |
|||
|
Метафаза-I |
404 |
2,1 |
Диффузное распределение хромосом |
1,2 |
|
Выбросы отдельных хромосом за пределы ахроматинового веретена |
0,3 |
|||
|
Атипичная форма клетки |
0,6 |
|||
|
Анафаза-I |
303 |
3,0 |
Разброс отдельных хромосом по веретену деления |
1,9 |
|
Выбросы отдельных хромосом за пределы ахроматинового веретена |
0,5 |
|||
|
Мосты |
0, 5 |
|||
|
Атипичная форма клетки |
0,1 |
|||
|
Телофаза-I |
361 |
12,1 |
Микроядра |
1,2 |
|
Изогнутая клеточная пластинка |
1,0 |
|||
|
Неполный цитокинез1 |
8,6 |
|||
|
Отсутствие цитокинеза |
0,7 |
|||
|
Атипичная форма клетки |
0,6 |
|||
|
Диада |
301 |
10,1 |
Монада двухъядерная |
0,7 |
|
Диада неравных клеток |
0,7 |
|||
|
Диада с микроядрами |
0,7 |
|||
|
Неполный цтокинез |
7,5 |
|||
|
Атипичная форма клетки |
0,5 |
|||
|
Метафаза-II |
227 |
6,3 |
Диффузное распределение хромосом |
1,7 |
|
Выбросы отдельных хромосом за пределы веретена деления |
0,5 |
|||
|
Асинхронные деления |
2,6 |
|||
|
Непараллельные веретена2 |
1,2 |
|||
|
Атипичная форма клетки |
0,3 |
|||
|
Анфаза-II |
320 |
15,6 |
Разброс отдельных хромосом по веретену деления |
1,6 |
|
Выбросы отдельных хромосом за пределы веретена деления |
0,5 |
|||
|
Мосты |
0,9 |
|||
|
Трехполюсные веретена |
1,0 |
|||
|
Асинхронность деления |
5,8 |
|||
|
Непараллельные веретена |
5,5 |
|||
|
Атипичная форма клетки |
0,3 |
|||
|
Телофаза-II |
246 |
4,2 |
Микроядра, сверхчисленные ядра |
2,2 |
|
Неполный цитокинез |
0,9 |
|||
|
Отсутствие цитокинеза |
1,0 |
|||
|
Атипичная форма клетки |
0,1 |
Примечание. 1 – цитокинез в виде «тоннеля» или «насечек»; 2 – веретена делений расположены в анафазе-II перпендикулярно или под углом друг к другу
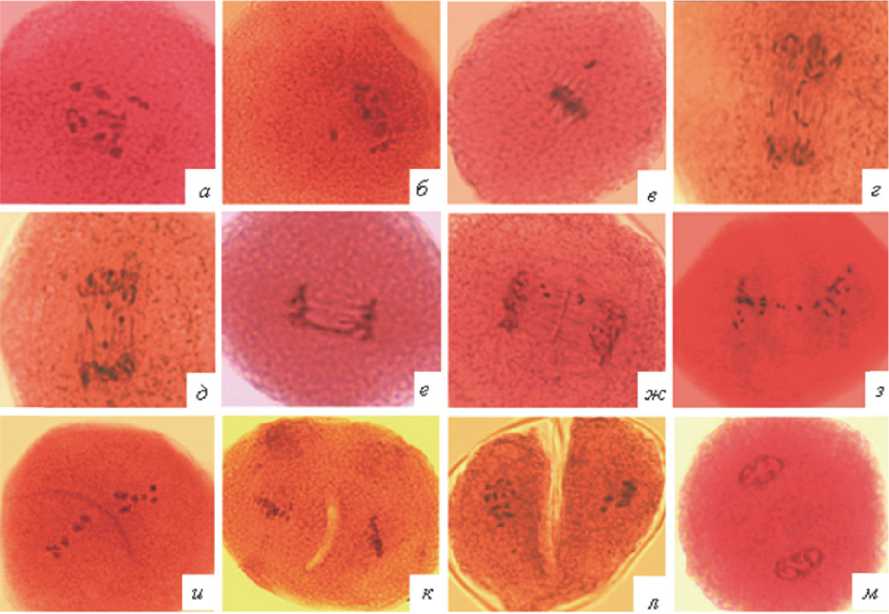
Рис. 1. Нарушения кариокинеза и цитокинеза в первом мейотическом делении: а – метафаза, разброс хромосом; б, в – метафаза, выбросы отдельных хромосом за пределы веретена; г, д – анафаза, разброс хромосом по веретену; е – хромосомный мост; ж, з – телофаза, отставания хромосом; и – изогнутая клеточная пластинка; к – цитокинез в виде «тоннеля»; л – цитокинез в виде «насечки»; м – диада, отсутствие цитокинеза
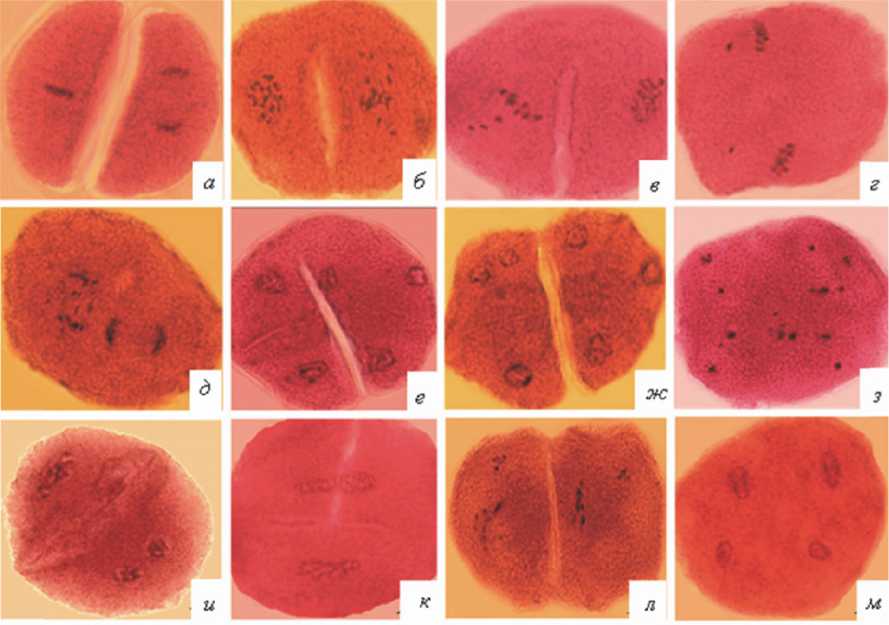
Рис. 2. Нарушения кариокинеза и цитокинеза во втором мейотическом делении:
а – асинхронные деления ядер в мета-анафазе; б, в – метафаза, диффузное распреление хромосом;
г – метафаза, выбросы отдельных хромосом за пределы веретена; д – анафаза, разброс хромосом по веретену и перпендикулярное расположение веретен деления; е, ж – трехполюсные веретена; з – образование микроядер; и, к – телофаза, слияние ядер; б-м – нарушения цитокинеза же нарушениями цитокинеза (1,9 %) (рис. 2б-м). Нередко в микроспороцитах на этой стадии наблюдается одновременно несколько аномалий. Такие комплексные изменения являются результатом нарушений карио- и цитокинеза, произошедших в первом и втором мейотиче-ских делениях.
Аномалии расхождения хромосом в мейозе зарегистрированы у всех изученных растений. Частота микроспороцитов с нарушениями кариокинеза у разных растений варьировала от 4 до 12 % (рис. 3). Максимальное число таких микроспороцитов обнаружено на стадиях анафазы-I и II (см. табл.). Причиной, по крайне мере, части аномалий может быть пониженный уровень гомологичной коньюгации хромосом, обнаруженный ранее у тетраплоидных растений кукурузы [5; 6]. Образовавшиеся при этом унивалентные хромосомы не могут быть правильно ориентированы в метафазе-I и беспорядочно распределяются между полюсами на стадии анафазы.
Аномальное распределение хромосом в анафазах-I и II определяется также различными нарушениями строения веретена деления. Связь нарушений расхождения хромосом в мейозе и аномалий веретен деления доказана на огромном материале, в том числе и кукурузе [9; 10; 11; 12]. У проанализированных нами растений обнаружены микроспороциты с беспорядочно разбросанными нитями веретена, трехполюсными веретенами, сближенными или расположенными под разными углами друг к другу.
Анализ цитокинеза показал значительную вариабельность отдельных тетраплоидных растений по данному признаку. У четырех растений нарушения в цитокинезе не выявлены, а у пяти частота микроспороцитов с аномальным цитокинезом варьирует от 6 до 69 % (рис. 4).
Среди различных вариантов цитокинеза два типа нарушений имеют наиболее значимые последствия для развития микроспор и пыльцы. Это отсутствие цитокинеза после первого и/или второго деления мейоза и неполное заложение перегородок в виде 1-2 «насечек» с внешней стороны или 1-2 «тоннелей» в центре клетки. Такие аномалии обычно наблюдались в первом деле-
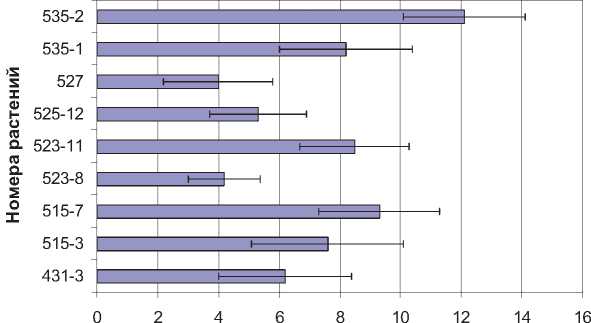
Микроспороциты с нарушениями расхождения хромосом, %
Рис. 3. Частота хромосомных аномалий в микроспорогенезе
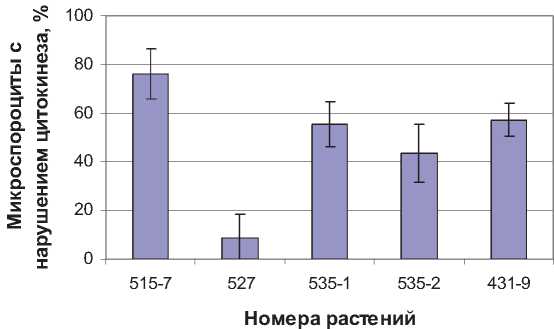
Рис. 4. Частота нарушений цитокинеза в микроспорогенезе
нии мейоза и были представлены в основном неполным заложением перегородок. Во втором делении нарушения цитокинеза встречались значительно реже. Только у одного растения № 431-9 частота микроспороцитов с неполным цитокинезом или его отсутствием в телофазе-II составила 20 %.
Известно, что причиной аномального цитокинеза у растений являются нарушения структуры веретена деления и образования фрагмопла-ста. Когда фрагмопласт/клеточная пластинка не полностью пересекает цитоплазму клетки, происходит остановка цитокинеза, и дочерние мембраны могут образовывать полый цилиндр («тоннель»), частично пронизывающий объем цитоплазмы, или «насечки» с внешней стороны [8; 13]. В проведенном нами исследовании микроспороциты с аномально расположенными веретенами деления, наблюдались у большинства растений, что, несомненно, приводило к нарушениям образования фрагмопласта, а в последующем и к различным изменениям формирования клеточной перегородки.
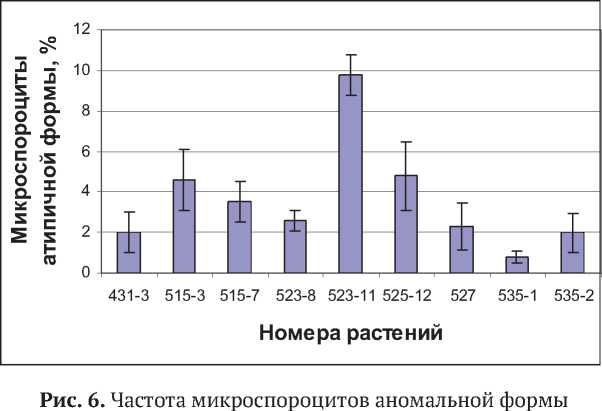
Важной особенностью микроспорогенеза у всех 9 проанализированных растений было развитие микроспороцитов атипичной формы. Они встречались на всех стадиях мейоти-ческого цикла, начиная с профазы-I (рис. 5а-г). В этом случае вместо округлых клеток формировались более крупные и удлиненные клетки. Ядро всегда располагалось в расширенной части микроспороцита, и его деление обычно проходило нормально. Цитоплазма вытянутой части клетки имела более бледную окраску. Частота микроспороцитов атипичной формы у разных растений варьировала от 0,8 до 9,8 % (рис. 6).
Практически у всех изученных ранее форм кукурузы разной плоидности подобные структуры в микроспорогенезе не встречались [14; 15; 16; 17]. Только у некоторых инбредных линий обнаружены единичные микроспороциты ненормальной вытянутой формы [18; 19]. Описанные авторами изменения, вероятно, могут быть отнесены к разряду случайных или спонтанных нарушений. Появление таких аномалий в микроспорогенезе у всех девяти проанализированных растений тераплоида КрП-1 со средней частотой 3,6 % предполагает наличие достаточно стабильного механизма образования клеток атипичной формы. В наших исследованиях
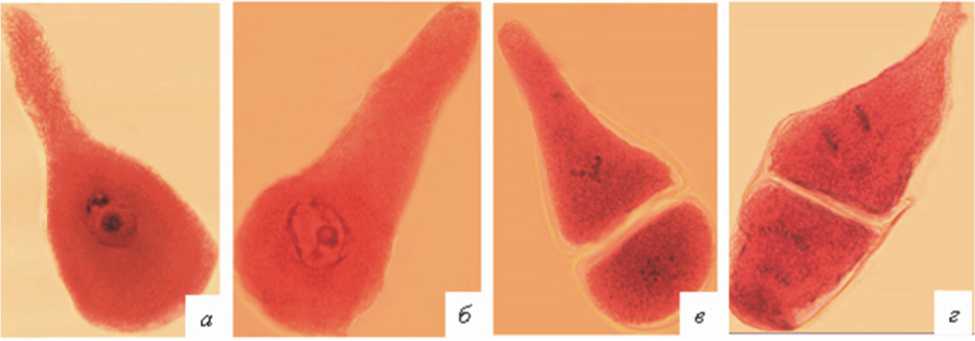
Рис. 5. Микроспороциты атипичной формы на разных стадиях мейоза: 1, 2 – профаза I; 3 – метафаза II; 4 – анафаза II
также было обнаружено, что данный признак передается при скрещивании тетраплоидной линии кукурузы КрП-1 с диплоидными линиями потомству, и появляется у триплоидных растений, что свидетельствует о его генетической обусловленности.
Не исключено, что появление микроспороцитов атипичной формы вызвано специфической мутацией, влияющей на форму мейо-тических клеток. Возможно, их образование является следствием аберрантного цитоскелета. Это предположение строится на основании данных о функциях цитоскелета, одна из которых – определение формы клетки. У растений при растяжении и росте клеток большое количество микротрубочек появляются в периферических слоях цитоплазмы и механически укрепляют ее. Известно, что деполимеризация микротрубочек, вызванная колхицином, сильно изменяет форму клеток [20]. Нельзя также игнорировать и возможную роль микрофиламентов в становлении формы клетки. Археспориальные клетки являются активно растущими клетками, и нарушение образования устойчивого внутриклеточного скелета (каркаса) может сказаться на внешнем виде образовавшихся микроспороцитов. Возможно, что данная тетраплоидная линия является уникальной для исследования мейоза и может пополнить генетическую коллекцию эмбриологических мутаций.
ЗАКЛЮЧЕНИЕ
Проведенный структурно-морфологический анализ позволил выявить особенности мейоза в микроспорогенезе растений тетраплоидной линии кукурузы КрП-1. Полученные результаты свидетельствуют, что аномалии мейоза у исследованной формы представлены тремя основными типами: нарушениями кариокинеза, цитокинеза и формированием микроспороцитов атипичной формы. Достоверные различия между отдельными растениями по количеству разных типов нарушений могут указывать на возможность отбора по этим признакам, как в сторону их увеличения, так и уменьшения.
Многие аномальные продукты мейоза не способны формировать жизнеспособную пыльцу, но определенные нарушения карио- и цитокинеза в микроспороцитах могут стать важным фактором мейотической реституции. Нельзя исключить и вероятность развития части аберрантных микроспороцитов в пыльцу с несбалансированными по числу хромосом гаметами. Однако высокий процент образования типичных микроспор свидетельствует о возможности использования тетраплоида КрП-1 в селекции в качестве донора диплоидных гамет. Несомненно, отклонения в мейозе тетраплоидных рас- тений обусловлены влиянием дополнительного генетического материала и определенным хромосомным дисбалансом, но изменения формы микроспороцитов могут указывать и на проявление в мейозе специфической мутации. Не исключено, что изученная тетраплоидная линия является интересным объектом для исследования закономерностей мейоза, онтогенеза пыльцы и понимания разнообразных задач клеточной биологии.
Список литературы Особенности микроспорогенеза у тетраплоидной линии кукурузы КРП-1
- Lobanova L.P., Kolesova A.Y. Maize tetraploid pollen studies // Maize Genetics Cooperation Newsletter. 2010. Vol. 84. P. 36.
- Особенности развития пыльцы у тетраплоидной кукурузы / Л.П. Лобанова, А.Ю. Колесова, И.С. Анохина, Н.Л Долотова // Бюллетень Ботанического сада Саратовского государственного университета. 2010. Вып. 9. С. 156-161.
- Цитоэмбриологическое исследование тетраплоидной кукурузы, склонной к партеногенезу / А.Ю. Колесова, Л.П. Лобанова, Ш.В. Магеррамов, А.А Полубабкина // Сборник статей межд. науч.-практ. конф. посвященной 130-й годовщине со дня рождения академика Н.И.Вавилова. Саратов: Саратовский ГАУ, ООО "Амирит", 2017. С. 205-207.
- Характеристика спорад микроспор у тетраплоидной линии кукурузы КрП-1 / Л.П. Лобанова, А.Ю. Колесова, О.А. Шацкая, Ш.В Магеррамов // Известия высших учебных заведений. Поволжский регион. 2018. № 4 (24). С. 62-71.
- Лаптев Ю.П. Гетероплоидия в селекции растений. М.: Колос, 1984. 248 с.
