Особенности ольфакторной коммуникации самцов лесной генетты (Genetta pardina) при парных ссаживаниях
 при парных ссаживаниях")
Автор: Петрина Татьяна Николаевна, Петрин Андрей Алексеевич, Рожнов Вячеслав Владимирович
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Статья в выпуске: 4, 2014 года.
Бесплатный доступ
Изучали обонятельные контакты самцов лесной генетты (Genetta pardina) при кратковременных парных ссаживаниях животных. Выявлен высокий уровень активности обнюхиваний животными друг друга в июле-августе. Области с высокой концентрацией кожных желез животные обнюхивают более интенсивно. Существенных отличий в обонятельных реакциях резидентов и интродуцентов не выявлено.
Ольфакторная коммуникация, обонятельные контакты, парные ссаживания, лесная генетта
Короткий адрес: https://sciup.org/146116536
IDR: 146116536 | УДК: 591.58:599.742.5
Peculiarities of olfactory contacts between the pardine genett (Genetta pardina) males in the pair encounter experiments
The olfactory contacts between males of the Pardine genet Genetta pardina were studied in the pair encounter experiments. The high level of olfactory activity was registered in July-August. The sniffing was more intense in the areas with high concentration of skin glands. There were no differences in the olfactory reaction between residents and invasive males.
Текст научной статьи Особенности ольфакторной коммуникации самцов лесной генетты (Genetta pardina) при парных ссаживаниях
Введение. Сведений об особенностях биологии представителей рода Genetta мало; по многим видам они практически отсутствуют. Для этих небольших полудревесных ночных хищных, ведущих преимущественно одиночный образ жизни, остается невыясненным вопрос о характере внутривидовых взаимоотношений. Социальные связи между особями у видов, ведущих в основном одиночный образ жизни, в значительной степени обеспечивает опосредованная коммуникация (Рожнов, 2004, 2011). Ведущую роль приобретает передача запаховой информации от особи к особи при прямых контактах посредством взаимных обнюхиваний, число которых возрастает в период активности репродуктивной системы (Roeder, 1980; Рожнов, Найденко, 1998). В этой связи актуальным представляется изучение особенностей ольфакторной коммуникации самцов лесной генетты (Genetta pardind) при прямых взаимодействиях.
Методика. Работа выполнена на научно-экспериментальной базе «Черноголовка» Института проблем экологии и эволюции им. А.Н. Северцова РАН в мае-августе 2013 г. В эксперименте участвовало 3 половозрелых самца генетт возрастом 6, 10 и 12 лет. Животных содержали в индивидуальных вольерах площадью 4 м2 с убежищем и сложным интерьером. В ходе эксперимента одного из зверьков (интродуцента) подсаживали в вольеру к другому (резиденту) на 1 ч и регистрировали все контакты между животными. Всего проведено 67 ч наблюдений.
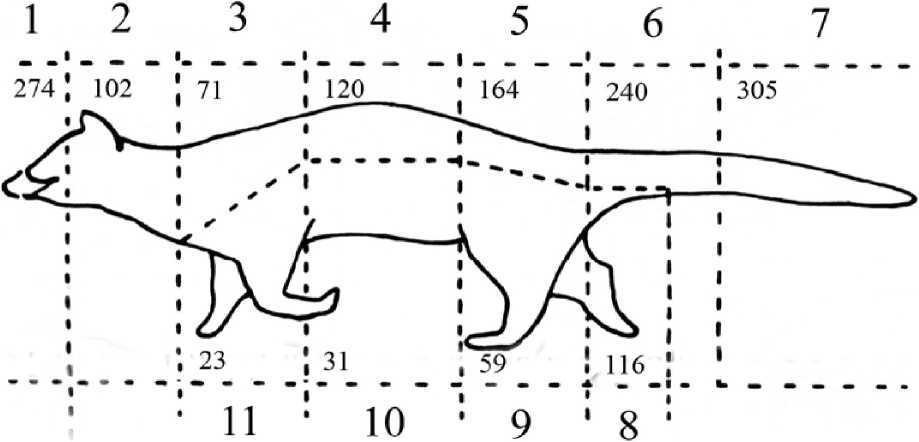
Рис. 1. Общее число обонятельных контактов самцов лесных генетт, зарегистрированное за весь период наблюдений
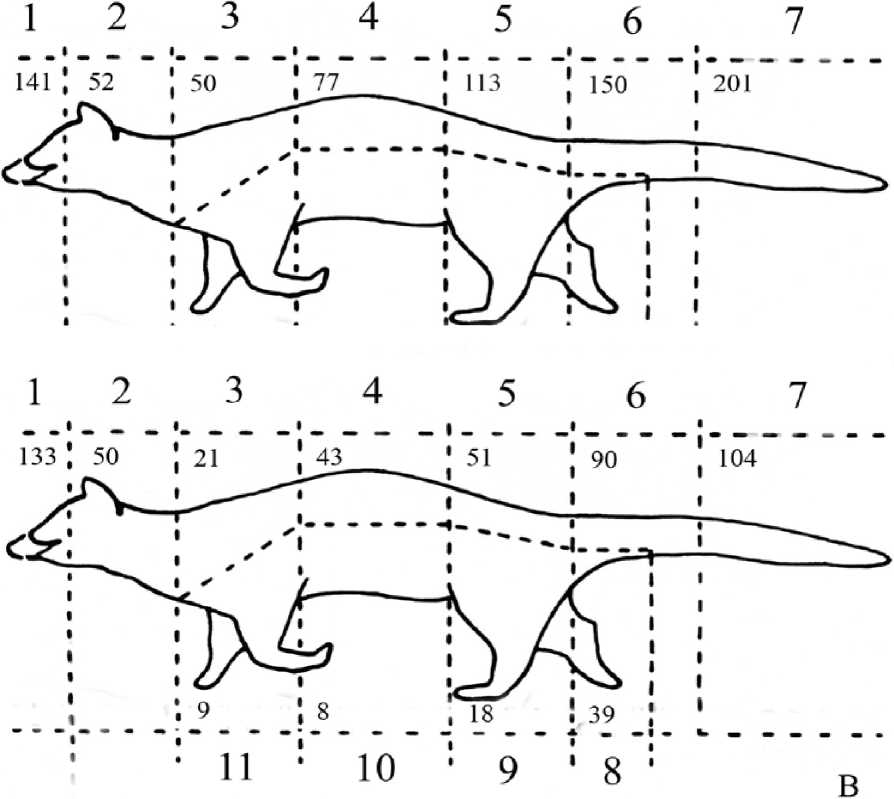
Рис. 2 . Общее число обонятельных контактов резидента (А) и интродуцента (В) за весь период наблюдений
На теле генетт мы выделили 11 областей (рис. 1, 2), на которые направлены обонятельные контакты других особей: 1 - носо-глазничная (от кончика носа до глаз, включительно); 2 - заглазнично-шейная (от заглазничной области до уровня лопаток); 3 - область лопаток; 4 -спина; 5 - поясница; 6 - основание хвоста и его первая треть; 7 -собственно хвост; 8 - ано-генитальная область; 9 - задние конечности; 10 - брюхо; 11- передние конечности.
Обработку результатов проводили общепринятыми методами статистического анализа с использование программ Microsoft Excel и Statistica (StatSoft).
Результаты. В общей сложности было зафиксировано 1505 обонятельных контакта: 939 - со стороны резидента и 566 - со стороны интродуцента. Первый контакт между самцами происходил в среднем на 8 мин. после ссаживания животных (7,9±1,5; lim 1-52, п=67).
Таблица 1
Среднее значение числа обонятельных контактов между самцами генетты за весь период наблюдений
|
Области обнюхивания |
все самцы п=134 |
резидент п=67 |
интродуцент п=67 |
||||
|
М±т |
о |
М±т |
о |
М±т |
о |
||
|
1 |
носо-глазничная |
2,0±0,3 |
3,3 |
2,1±0,4 |
3,4 |
2,0±0,4 |
3,4 |
|
2 |
заглазнично-шейная |
0,8±0,1 |
1,5 |
0,8±0,2 |
1,6 |
0,7±0,2 |
0,9 |
|
3 |
лопаточная |
0,5±0,1 |
1,2 |
0,7±0,2 |
1,5 |
0,3±0,1 |
0,5 |
|
4 |
спинная |
0,9±0,1 |
1,4 |
1,1±0,2 |
1,6 |
0,6±0,1 |
U |
|
5 |
поясничная |
1,2±0,2 |
2,1 |
1,7±0,3 |
2,6 |
0,8±0,2 |
1,1 |
|
6 |
основание хвоста и его первая треть |
1,8±0,2 |
2,8 |
2,2±0,4 |
3,3 |
1,3±0,2 |
1,9 |
|
7 |
собственно хвост |
2,3±0,3 |
3,2 |
3,0±0,5 |
3,8 |
1,5±0,3 |
2,1 |
|
8 |
ано-генитальная |
0,9±0,1 |
1,4 |
1,1±0,2 |
1,7 |
0,6±0,1 |
0,9 |
|
9 |
задние конечности |
0,4±0,1 |
0,9 |
0,6±0,1 |
1,1 |
0,3±0,1 |
0,5 |
|
10 |
брюхо |
0,2±0,004 |
0,5 |
0,3±0,1 |
0,7 |
0,1±0,04 |
0,3 |
|
11 |
передние конечности |
0,2±0,05 |
0,5 |
0,2±0,1 |
0,6 |
0,1±0,05 |
0,3 |
За все время наблюдений значительная доля обнюхиваний самцов генетт была направлена на хвост (20%) и носо-глазничную области (18%). При этом следует отметить, что у резидента значительное число обонятельных реакций, помимо этих двух областей (21 и 15%, соответственно), было направлено на область основания хвоста интродуцента (16% от общего числа обонятельных контактов), поясницу (12%), спинную (8%) и ано-генитальную (8%) области (рис.З)
резидент
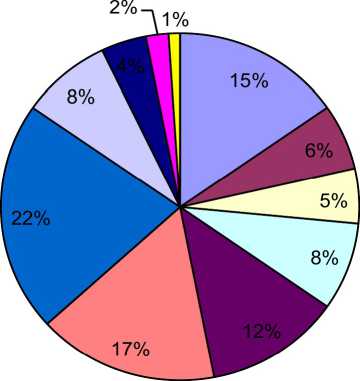
Рис. 3 . Распределение обонятельных контактов по областям тела самцов-резидентов (расшифровку обозначений см. в табл. 1).
интродуцент
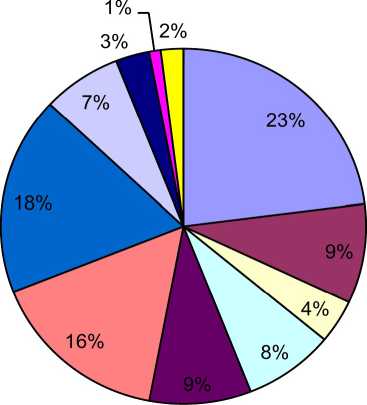
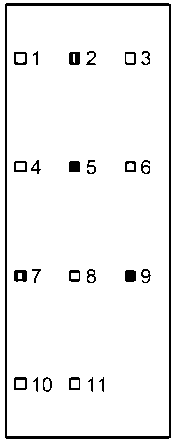
Рис. 4. Долевое распределение обонятельных контактов по областям тела самцов-интродуцентов (расшифровку обозначений см. в табл. 1).
У интродуцента значительное число обонятельных реакций было направлено на носо-глазничную область резидента (23%, от общего числа обонятельных контактов), хвост (18%), основание хвоста (16%), поясницу (9%) и заглазнично-шейный отдел (9%) (Рис. 4).
Достоверные различия в ольфакторной коммуникации между реципиентом и интродуцентом за весь период наблюдений были отмечены лишь для области лопаток (р<0,05; U=1797; П1=П2=67); резидент обнюхивал эту область чаще.
Большая часть обонятельных контактов между животными произошла в июле и августе. Распределение контактов резидент/интродуцент по месяцам (в скобках - число ссаживаний) следующее: май 116/92 (12), июнь 91/39 (19), июль 352/203 (18), август 380/232 (18). Всплеск общей ольфакторной активности животных в июле достоверен (р<0,05; U=397; ni=38; П2=39). При рассмотрении по зонам тела в этом месяце достоверно увеличилось обнюхивание 1, 2, 4, 6, 7 и 8 областей (U=357,5; 467; 479; 414,5; 411,5; 420 при ni=38; п2=39, соответственно).
Из трех самцов разного возраста, самый старший имел наибольшее среднее значение обонятельных контактов, направленных на поясницу (1,4±0,5), основание хвоста (2,3±0,6) и хвост (3,0±0,7). Самый молодой - на носо-глазничную (2,7±0,5) и заглазнично-шейную области (0,9±0,2).
Обсуждение. Представителей рода Genetta относят к ночным хищникам, ведущих преимущественно одиночный образ жизни. Социальные группы образуют (1) мать с детенышами на период выращивания потомства и (2) самец с самкой на непродолжительный период спаривания. Вероятны встречи самцов друг с другом в период гона неподалеку от одной и той же самки, а также в период расселения молодых. Вопрос о территориальном распределении в природе этих хищников до сих пор остается открыт. Сведения о привязанности животных к определенной территории малочисленны и противоречивы: одни авторы считают, что самки более привязаны к территории, чем самцы (Carpenter, 1970; Fuller et al., 1990), другие предполагают отсутствие такой привязки в целом (Wemmer, 1977).
Значительная часть обнюхиваний направлена на хвостовую и носо-глазничную область, что объясняется способом приближения животных друг к другу. Как правило, при первом контакте зверь приближается к оппоненту со стороны хвоста. Если происходит обнюхивание носо-глазничной области, это свидетельствует о достаточно сильной мотивации к «знакомству» при достаточно низкой агрессии. Те области, на которые приходится значительная доля обонятельных контактов генетт, имеют большое количество неспецифических и специфических желез, выделяющих секрет с сильным запахом (Соколов, Чернова, 2001).
Данные, полученные нами в результате парных ссаживаний самцов, демонстрируют увеличение ольфакторной активности животных в июле-августе. Это отражает, на наш взгляд, их физиологическое (гормональное) состояние и готовность к контактам с особями своего вида в этот период. Ранее сходную активность мы отмечали в исследованиях ольфакторной коммуникации генетт разного пола (Рожнов, Петрина, 2005; Петрина и др., 2014), в которых были зарегистрированы максимальные значения числа обонятельных контактов животных в июле и августе. С учетом данных по встрече беременных особей и молодняка в природе (Wemmer, 1977), а также размножению разных видов генетт в условиях неволи (Гилева, 2006; Петрина и др., 2012), можно выделить два периода, благоприятных для спаривания - с июня по сентябрь и с января по апрель.
Существенных отличий в ольфакторной коммуникации самцов-резидентов и самцов-интродуцентов мы не выявили. Отмечено некоторое смещение в большую сторону доли обнюхивания носоглазничной области у интродуцента, что косвенно может служить показателем сниженной агрессии. Об этом же, на наш взгляд, свидетельствует увеличение среднего значения этих контактов у молодой особи. По нашим наблюдениям, молодая особь, как правило, менее агрессивна.
Заключение. Ольфакторная коммуникация самцов генетт в целом укладывается в общую картину этого типа взаимодействия у Genetta pardina. Увеличение ольфакторной активности генетт может быть связано с физиологическим состоянием животного (его гормональной активностью) и совпадает с благоприятным для спаривания периодом. Интенсивному обнюхиванию подвергаются области с высокой концентрацией желез, что способствует получению животными информации друг о друге. Значительная доля назо-нозальных контактов может свидетельствовать о том, что взаимодействие происходит на фоне снижения агрессии. В целом, отсутствие выраженных отличий в обонятельных реакциях резидентов и интродуцентов может свидетельствовать в пользу гипотезы об отсутствии строгой территориальности у самцов этого вида.
Список литературы Особенности ольфакторной коммуникации самцов лесной генетты (Genetta pardina) при парных ссаживаниях
- Гилева М.Ю. 2006. Размножение пятнистых генетт (Genetta tigrina) в Екатеринбургском зоопарке//Научные исследования в зоологических парках. Вып. 19.
- Петрина Т.Н., Петрин А.А, Рожнов В.В. 2012. Способность самцов лесной генетты (Genetta pardina) различать пол особи своего вида по запаху экскрементов//Материалы научно-практической конференции «Экология, эволюция и систематика животных». Рязань: НП «Голос губернии». С. 350-351.
- Петрина Т.Н., Петрин А.А, Рожнов В.В. 2014. Поведение и поведенческая экология млекопитающих//Материалы 3-й научной конференции 14-18 апреля 2014 г., г. Черноголовка. М.: КМК. C. 93.
- Рожнов В.В. 2004. Опосредованная коммуникация млекопитающих: о смене парадигмы и новом концептуальном подходе в исследовании маркировочного поведения//Зоол. журн. Т. 83. № 2. С. 132-158.
- Рожнов В.В. 2011. Опосредованная хемокоммуникация в социальном поведении млекопитающих. М.: КМК. 288 с.
- Рожнов В.В., Найденко С.В. 1989. Динамика контактов между черными хорями при парных ссаживаниях в течение года//Поведение, коммуникация и экология млекопитающих. Сб. науч. работ. М.: ИПЭЭ РАН. С. 132-154.
- Рожнов В.В., Петрина Т.Н. 2005. Обонятельные контакты в семейной группе лесной генетты (Genetta pardina)//Поведение и поведенческая экология млекопитающих. Материалы научной конференции. М.: КМК. С. 214-216.
- Соколов В.Е., Неклюдова Т.И. 1988. Морфология кожи и специфических кожных желез некоторых представителей семейства виверровых (Viverridae) Вьетнама//Экол.-функц. морфол. кожн. покр. млекопитающих. М.: Наука. С. 4-62.
- Carpenter, G.P. 1970. Some observation on the Rusty Spotted Genet Genetta rubiginosa zuluensis. Lammergeyer V. 11. P. 60-63.
- Fuller T.K., Biknevicius A.R., Kat P.W. 1990. Movements and behavior of large spotted genets (Genetta maculata Gray 1830) near Elmenteita, Kenya (Mammalia, Viverridae)/Tropical Zoology. V. 3. P. 13-19.
- Roeder J.-J. 1980. Marking behavior and olfactory recognition in genets (Genetta genetta L., Carnivora -Viverridae)//Behaviour. V. 72. № 3-4. P. 200-210.
- Wemmer C.M. 1977. Comparative ethology of the Large-spotted genet (Genetta tigrina) and some related Viverrids. Smithsonian Contrib. Zool. № 239. P. 1-93.
