Особенности онтогенеза трех однолетних видов рода Pimpinella (Apiaceae) в связи с их экологией
 в связи с их экологией")
Автор: Петрова Светлана Евгеньевна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 2, 2014 года.
Бесплатный доступ
Изучен онтогенез трех однолетних видов рода Pimpinella (Apiaceae). Pimpinella eriocarpa и P. barbata отнесены к группе эфемеров (продолжительность развития около 2,5 месяцев). P. anisum является длительно вегетирующим яровым однолетником (онтогенез занимает около 4-5 месяцев). Все три изученных вида имеют световые мезоморфные черты строения. У P. eriocarpa, P. barbata, обитающих в природе на сухих открытых пространствах, мезоморфный облик - следствие ускоренного развития: весь жизненный цикл растения успевают пройти в относительно влажные месяцы до наступления летней засухи. По особенностям строения и ритму развития P. eriocarpa и P. barbata имеют значительно большее сходство друг с другом, чем с P. anisum.
Онтогенез, эфемеры, длительно вегетирующие однолетники, мезофиты
Короткий адрес: https://sciup.org/146116515
IDR: 146116515 | УДК: 581.412:
Ontogenesis of three annual species of Pimpinella (Apiaceae) in connection with their ecology
Ontogenesis of three annual species of Pimpinella (Apiaceae) have been studied. P. eriocarpa and P. barbata are characterized as ephemeral plants (entire life cycle takes nearly 2,5 months), whereas P. anisum is long vegetating annual plant (ontogenesis takes nearly 4-5 months). All species have features of heliophytic and mesomorphical organisation. P. eriocarpa, P. barbata, inhabiting open dry ecotops, are mesophyts. They are adapted to fast completion of entire life cicle in relatively wet months before the summer drought. P. eriocarpa and P. barbata in structure and rhythm of development are more close to each other than to P. anisum.
Текст научной статьи Особенности онтогенеза трех однолетних видов рода Pimpinella (Apiaceae) в связи с их экологией
Род Pimpinella - бедренец - является одним из крупнейших в семействе Apiaceae (насчитывает около 150 (Чиков, 2002) - 180 (Пименов, Остроумова, 2012) видов, распространенных в Азии, Европе, Африке, в Америке и Австралии как заносные), однако его окончательный объем до сих пор является дискуссионным. Жизненные формы бедренцев разнообразны, но наиболее часто встречаются многолетние растения; однолетники редки. Однако последние вызывают определенный интерес как с позиций систематики, так как потенциально могут оказаться обособленной таксономической группой, так и с точки зрения экологической и структурной эволюции малолетников, до сих пор до конца не изученной. Сравнительный анализ хода онтогенеза, особенностей макро- и микроструктуры представителей однолетних бедренцев может способствовать решению указанных проблем.
Объектами исследования стали три вида бедренцев: Р. eriocarpa Bank., Р. barbata (DC.) Boiss. и P. anisum L. P. eriocarpa распространен в Иране, Ираке, Персии, Палестине и Сирии (Rechinger, 1987); обитает в пустынях, на сухих открытых склонах, в степях, на высоте до 1650 м. Р. barbata распространен в Иране, Ираке, Персии (Rechinger, 1987); встречается в сухих долинах, степях, на открытых склонах, поднимается на высоту до 1600 м. Р. eriocarpa и Р. barbata содержат потенциально интересный для практического использования состав эфирных масел (Askari et al., 2004; Fakhari, Sonboli, 2006). P. anisum культивируется в южных районах. Встречается на юге Европы, в Китае, Турции, Йемене, Израиле, Кипре (Пименов, Остроумова, 2012). Сейчас известен только в культуре и как дичающее. Имеет важное хозяйственное значение: эфирное масло, содержащееся в плодах применяется в парфюмерии, в ликероводочном производстве и в пищевой промышленности (Шишкин, 1950).
Описания отдельных фаз онтогенеза Р. anisum приводятся в различных источниках по лекарственным растениям (Чиков, 2002), однако имеющиеся сведения представляются краткими и неполными. Сколько-нибудь подробная информация по морфологии, анатомии, онтоморфогенезу видов Р. eriocarpa и Р. barbata в литературе отсутствует.
Методика. Онтогенез Р. eriocarpa и Р. barbata изучали на особях, выращенных в ботаническом саду МГУ из семян (мерикарпиев), собранных в Иране в 2001 году проф. М.Г. Пименовым и к.б.н. Е.В. Клюйковым и посеянных осенью 2003 года. Зрелые плоды Р. anisum были переданы в том же году сотрудником ботанического сада к.б.н. Т.А. Остроумовой и посеяны весной того же года в горшки без предварительной стратификации. С целью дальнейшего анатомического анализа вегетативных органов разновозрастные растения фиксировали в 70% этиловом спирте. Для уточнения жизненных форм отдельных видов был просмотрен гербарный материал в гербариях ЕБС РАН (МПА), МГУ (MW), ботанического сада МГУ.
Периодизацию жизненного цикла проводили по методике, предложенной Работновым (1950), используя также ряд современных работ по онтогенезу однолетников (Онтогенетический..., 2005, 2011). Характеристику жизненных форм давали с учетом классификации Серебрякова (1962). Анатомическое строение листьев изучали на временных препаратах. Срезы (поверхностные, поперечные) делали лезвием от руки. Плотность устьиц подсчитывали, находя среднее арифметическое из сделанных промеров (не менее 30), затем делали пересчет на 1 мм2. Коэффициент палисадности (КП) - отношение высоты клеток столбчатого мезофилла к общей толщине мезофилла -выражен в процентах.
Онтогенез . Латентный период. В зрелых семенах Р. eriocarpa зародыш относительно крупный, длиной 0,52 мм, длина эндосперма составляет в среднем 1,20 мм. Соотношения длин зародыша и эндосперма 0,4, семядолей и осевой части - 0,9. Зародыш Р. barbata - 0,30-0,37 мм, эндосперм хорошо развит, 1,22 мм дл., отношение длины зародыша к длине эндосперма около 0,28. Зародыш у Р. anisum наиболее крупный, длиной 0,83 мм, что в некоторой степени " - 85 -
Вестник ТвГУ. Серия "Биология и экология". 2014. № 2 коррелирует с большим размером семени и плода, длина эндосперма около 2,95 мм. Соотношение длин зародыша и эндосперма - 0,28, семядолей и осевой части зародыша -0,75.
Виргинильный период. Семена Р. eriocarpa и Р. barbata, посеянные в ящики с землей поздней осенью, начали прорастать в апреле. В природе (Иран, Ирак, Персия) проростки появляются в марте. Семена Р. anisum прорастают в благоприятных условиях через 17-30 дней после посева даже без стратификации, однако в первое время растут очень медленно. Прорастание у всех видов надземное гипокотилярное. Сначала появляется зародышевый корешок, затем вытягивается гипокотиль, который выносит семядоли на поверхность почвы. У Р. eriocarpa (рис. 1А) и Р. barbata (рис. 2А) пластинки семядолей ланцетовидные, у первого - 8,5 х 1,85 мм, у второго - 5,5 х 1,0 мм; длина черешка составляет соответственно 7 и 5 мм. Пластинки семядолей у проростков Р. anisum продолговато-ланцетные, 8 х 1,8 мм, со слегка закругленной верхушкой, основание пластинки плавно переходит в черешок.
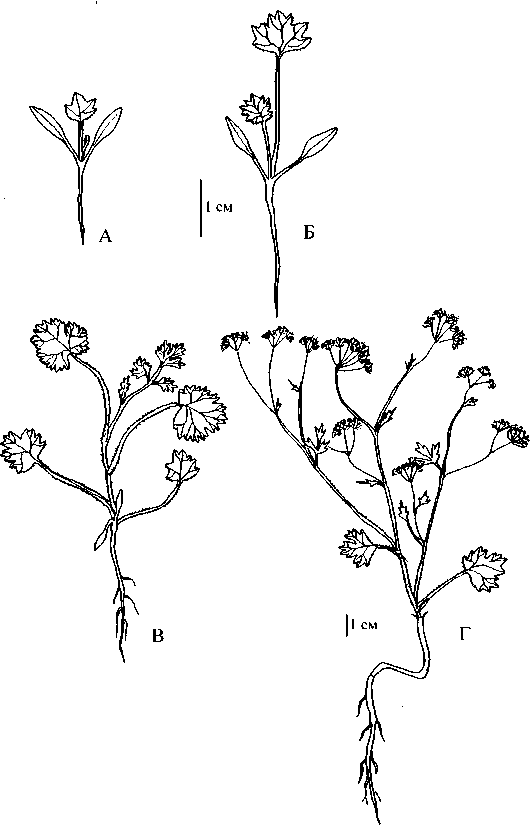
Рис. 1 . Разновозрастные особи Р. eriocarpa;
А, Б - ювенильные, В - имматурная, Г - генеративная
С развертыванием из почечки первого листа, что наблюдается (в условия ботанического сада) к началу мая, растения вступают в ювенильную фазу развития. Листовая пластинка первого листа Р. eriocarpa -3,5 х 4 мм, округлая цельная с крупнозубчатым краем или же неравно трех-четырехлопастная, с клиновидным основанием, черешок длиной около 2,5 мм, влагалище короткое (рис. 1Б). Черешок и крупные жилки листовой пластинки опушены простыми волосками. Гипокотиль плавно переходит в короткий 10-15 мм, неразветвленный главный корень. Боковые корни появляются лишь с развитием последующих двух-трех листьев. Первый лист у ювенильных особей Р. barbata опушенный, с более или менее округлой трехлопастной по краю крупнозубчатой листовой пластинкой, клиновидно переходящей в несколько превышающий ее по размеру черешок (рис. 2А). Главный корень относительно короткий и тонкий. Первый лист Р. anisum имеет в очертании яйцевидно-почковидную цельную с сердцевидным основанием и острозубчатым краем пластинку и длинный черешок. Главный корень углубляется в почву на 15-20 мм.
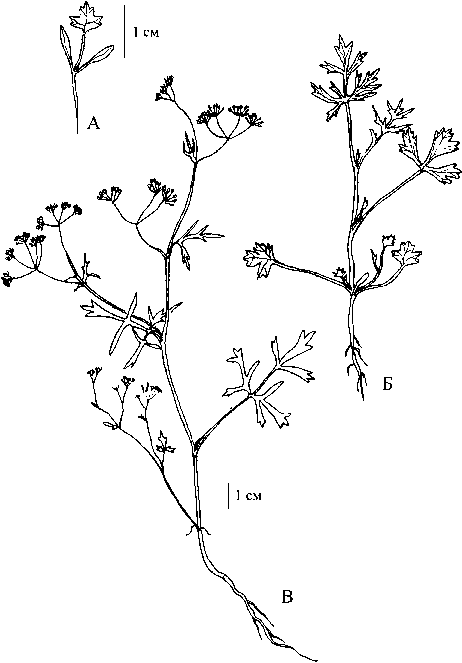
Рис. 2 . Разновозрастные особи Р. barbata; А - ювенильная, Б - имматурная, В - генеративная
У Р. eriocarpa и Р. barbata на главном побеге вслед за двумя метамерами с укороченными междоузлиями и двумя первыми (сходными по структуре) ювенильными листьями развиваются метамеры с удлиненными междоузлиями. По мере наращивания побега степень сложности листовой пластинки на них постепенно возрастает. Так, у Р. eriocarpa (рис. 1В) нижние стеблевые листья черешковые, опушенные в основном по жилкам шиловидными волосками, с округлой трехлопастной или трехраздельной, по краю зубчатой пластинкой. Их сменяют листья перисторассеченные с острозубчатыми, иногда трехлопастными сегментами. Одновременно с последующим разрастанием листовых пластинок из пазух третьего-четвертого листа начинают трогаться в рост боковые побеги.
Для Р. barbata (рис. 2Б) характерно более быстрое увеличение степени расчлененности листовой пластинки, чем для Р. eriocarpa. Уже третий стеблевой лист трехрассеченный, а последующие перисто- и дважды перисторассеченные с ланцетовидными конечными сегментами. Листья опушены неравномерно: верхние в меньшей степени, чем нижние. В пазухе каждого листа имеются пазушные почки, реализация которых происходит с самых ранних этапов онтогенеза по ходу нарастания побега вслед за формированием листовых пластинок. То есть для вида характерны силлептические побеги обогащения и базитонное ветвление.
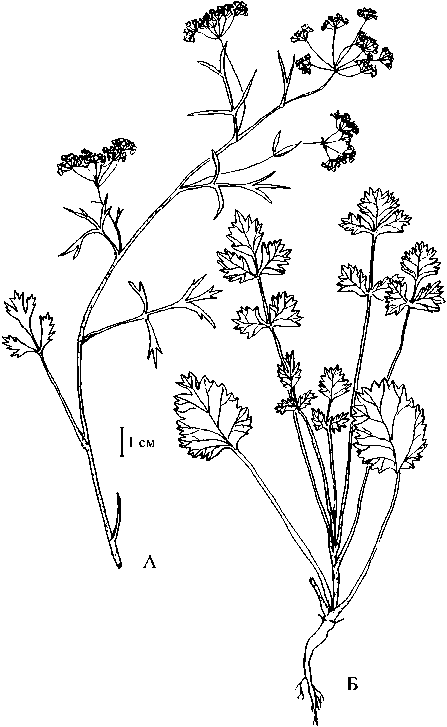
Рис. 3 . Разновозрастные особи Р. anisum-.
X - верхняя часть репродуктивного побега генеративной особи, Б - имматурная особь
По-иному (в отличие от Р.eriocarpa, Р.barbata) идет нарастание и ветвление побега у Р. anisum. Часто до 5 нижних междоузлий слабо удлиняются, фактически создавая полурозеточную структуру, боковые побеги развиваются только на срединных и верхних метамерах (из пазух 9-11 листа). Изменение морфологии листьев происходит по схеме: три-четыре первых листа цельные с неравнокрупнопильчатым краем, до четырех вышележащих - тройчатосложные с единственной парой боковых листочков, резко отставленной от терминального, последующие листья перистосложные с двумя парами боковых листочков (рис. ЗБ).
Описанные выше изменения: развитие удлиненных междоузлий, изменение формы листьев и начало роста боковых побегов указывают на то, что растения перешли в качественно иную фазу развития, имматурную. Семядоли при этом, как правило, еще сохраняются. Что касается главного корня, то он достигает длины 4-6 см, имеет несколько боковых ответвлений первого порядка, но в целом относительно слабо развит.
Взрослые вегетативные особи отличаются от имматурных большими линейными размерами главного и боковых побегов, иногда в этом периоде начинают отмирать семядоли.
Генеративный период. Формирование побеговой системы заканчивается дифференциацией соцветий - малолучевых двойных зонтиков, что знаменует переход растений в генеративный период развития. У Р. eriocarpa и Р. barbata развитие идет быстро, в условиях культуры (ботанический сад МГУ) уже к началу июня растения переходят к цветению, плоды созревают в первых числах июля. В природе период цветения нередко начинается в марте - апреле, зрелые плоды можно видеть в мае - июне. Генеративным особям Р. eriocarpa (рис. 1Г) присущ удлиненный (с эфемерной малочленной розеткой из 2 листьев) сильноразветвленный цветоносный побег. Ветвление мезотонное, наиболее сильно развитые боковые цветоносные побеги отходят от средних метамеров, почки же в пазухах нижних листьев развиваются медленнее. Стебель округлый со слегка выступающими ребрами, редко опушенный в основном в нижней части короткими простыми шиловидными волосками, верхние междоузлия голые, цветоносы опушены более длинными, отстоящими волосками. До терминального соцветия развивается 6-7 листьев. Самые верхние листья отличаются от нижележащих (описанных выше) редким опушением, маленькой или совсем редуцированной пластинкой, сидячей на узком цилиндрическом влагалище с тонко пленчатым краем. Корневая система представлена коротким (до 5 см) веретеновидным главным корнем с небольшим числом боковых ответвлений, редко имеющих оси второго порядка.
Для генеративных растений Р. barbata (рис. 2В) характерно
-89- наличие сильноразветвленного (до четвертого порядка) удлиненного главного побега. Ветвление часто базитонное, первыми раскрываются наиболее крупные почки, сформированные в пазухах нижних и средних листьев, из них развиваются удлиненные олиственные боковые цветоносные побеги обогащения. До терминального соцветия образуется около 7-9 листьев, однако самые нижние, сформированные еще на ювенильном этапе развития, рано засыхают и опадают. Самые верхние листья - сидячие дважды тройчатые, с почти редуцированной пластинкой из линейных сегментов. Стебель более или менее округлый, неравномерно опушенный длинными волосками. Корневая система представлена главным и боковыми корнями, как правило, разветвленными не выше второго порядка. После цветения и плодоношения растение отмирает. Р. eriocarpa и Р. barbata при развитии в культуре, несмотря на сохранение типичного для каждого вида плана строения и архитектоники монокарпического побега, в целом были менее разветвленными по сравнению с растениями, произрастающими в естественной среде обитания.
У Р. anisum репродуктивный период по длительности охватывает около трети всего большого жизненного цикла. Цветение начинается в июне, плоды созревают к концу августа. Генеративный побег удлиненный (в основании часто сохраняются укороченный метамеры, на которых может быть до 5 листьев, образованных в ювенильную фазу развития, самые нижние из них рано засыхают и опадают), высотой до 30—50 см, слабо акро- или мезотонно ветвящийся. Всего до терминального соцветия развивается 9-10 листьев. Вдоль побега хорошо выражена ярусная (одновременно и возрастая) гетерофиллия. При этом четко можно разделить листья, образованные в виргинильный период (описание их дано выше (рис. ЗБ)) и листья, сформированные при переходе растения к репродукции, то есть кроющие литья паракладий (рис. ЗА). Последние короткочерешковые или сидячие дважды или трижды перисторассеченные с линейными конечными сегментами, самые верхние из них мелкие трехраздельные или цельные с узким по краям пленчатым влагалищем. Стебель округлый, тонкоребристый, опушенный. Корневая система представлена стержневым слабо ветвящимся корнем.
Анатомия. Семядоли проростков у всех исследованных видов играют роль первых ассимилирующих органов и имеют типичную листовую организацию. Пластинки семядолей амфистоматические, бифациальные с отчетливо дифференцированным мезофиллом. Губчатая ткань включает 4 слоя слегка вытянутых или слаболопастных овальных (на поперечном сечении) клеток, относительно рыхло прилегающих друг к другу так, что между ними образуются небольшие межклетники. Максимальная толщина (около 0,28 мм) семядольной пластинки характерна для семядолей Р. eriocarpa (из-за большой длины -90- клеток палисадной ткани). Самые тонкие пластинки присущи Р. anisum, отличающиеся слабой дифференциацией мезофилла и небольшими размерами самих клеток. Семядоли отдельных видов различаются и характером устьичного аппарата. У Р. eriocarpa, Р. barbata он преимущественно диацитный, у Р. anisum - преимущественно аномоцитный. Замыкающие клетки устьиц невыступающие, наиболее мелкие они (также как и собственно эпидермальные клетки) у Р. anisum.
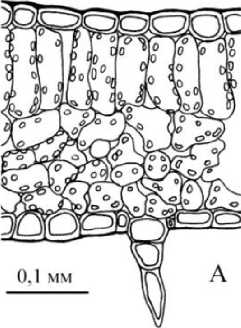
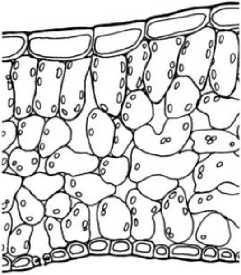
0,1 мм
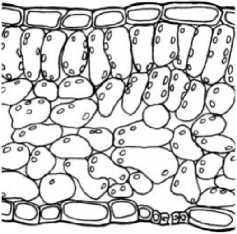
В
Рис. 4 . Микроструктура листовой пластинки дефинитивного листа: А - у Р. eriocarpa , Б - у Р. barbata, В - у Р. anisum
Пластинки дефинитивных листьев у Р. eriocarpa (рис. 4А) около 0,22 мм толщиной, амфистоматические, бифациальные, с толстым слоем кутикулы, мезофилл из одного слоя длинных палисад и 4 слоев губчатой хлоренхимы, КП около 43-45%. У Р. barbata (рис. 4Б)
листовые пластинки толщиной 0,26 мм, амфистоматические, бифациальные или изолатеральные, в последнем случае помимо одного адаксиального слоя столбчатой хлоренхимы имеется также слой перпендикулярно вытянутых клеток с абаксиальной стороны, КП 30% или 52%, соответственно. Губчатая хлоренхима рыхлая, четырехслойная, состоит из разнообразных по форме клеток. Клетки эпидермы листьев Р. eriocarpa, Р. barbata крупные с извилистыми антиклинальными стенками; число устьиц относительно невелико: 73±7 и 106±9 на 1 мм2 у Р. eriocarpa, 45±9 и 75±б на 1 мм2 у Р. barbata. Пластинки дефинитивных листьев Р. anisum (рис. 4В) 0,23 мм толщиной, амфистоматические, бифациальные, с 1-2 слоями столбчатой и 4-5 губчатой хлоренхимы, КП 30%, 48%. Листья у всех видов опушены дву-, многоклеточными шиловидными волосками.
Заключение. В результате исследования можно сделать вывод, что Р. eriocarpa и Р. barbata по характеру большого жизненного цикла и ритму сезонного развития принадлежат к группе эфемеров. Онтогенез у них проходит в сжатые сроки, как правило, не более 2,5 месяцев. При этом растения не успевают накопить достаточное количество ассимилятов для мощного развития как вегетативных, так и генеративных органов, в связи с чем оба вида характеризуются относительно мелкими листьями, коротким слаборазветвленным корнем, низким и слабым побегом. Увеличение репродуктивной мощи при небольшом числе метамеров достигается ранним образованием и быстрой реализацией пазушных почек, при этом почти все метамеры главного побега развивают боковые побеги флорального обогащения (базимезотонное ветвление). У обоих видов выражена гетерофиллия, и хотя листья подвергаются закономерным изменениям от простых через более сложные к вновь более простым, необходимость быстрого органообразования накладывает отпечаток на их структуру: они оказываются очень пластичными и разнообразными по форме, при чем эта форма, как правило, не укладывается в какой-либо типовой вариант. В большом жизненном цикле можно выделить несколько периодов и фаз развития: латентный, виргинильный (проростки, ювенильные, имматурные, виргинильные растения) и генеративный. Наблюдать их можно лишь во времени, так как при переходе к образованию репродуктивных единиц все растение уже фактически представляет собой синфлоресценцию. В целом, Р. eriocarpa и Р. barbata имеют сходную экологическую модель развития, а также много общего в структурной организации побеговой системы, определенные различия касаются лишь строения листьев и скорости развития боковых побегов. Следует отметить, что при выращивании растений в условиях средней полосы России (ботанический сад МГУ) сроки прохождения основных этапов онтогенеза смещаются (приблизительно на месяц) по сравнению с тем, что наблюдается в естественных условиях обитания. Однако
Вестник ТвГУ. Серия "Биология и экология". 2014. № 2 общий ритм развития сохраняется, при этом растения успевают успешно процвести и завязать полноценные плоды.
По-иному протекает онтогенез у Р. anisum. Развитие его растянуто во времени и занимает около 4-4,5 месяцев, то есть вид можно отнести к группе длительно вегетирующих яровых однолетников (Серебряков, 1962). Основные этапы, которые проходит растения во время большого жизненного цикла, можно увидеть не только в процессе, но и в структуре уже полностью сформированного побега. Так, в отличие от предыдущих видов самые нижние укороченные метамеры, сформированные на ювенильном этапе, и несколько слабо вытянутых вышележащих, образованных в имматурную фазу развития, не несут боковых цветоносных побегов, являясь как бы застывшим отражением ранних этапов онтогенеза. Несколько верхних метамеров с пазушными паракладиями представляют репродуктивную зону, сформированную на заключительных этапах развития - в генеративном периоде. В гетерофиллии Р. anisum выражена постепенная, но достаточно четко подразделяемая по форме, смена листьев, которая напоминает возрастные серии ряда многолетних видов рода, например Р. rhodantha, Р. saxifraga.
Анатомическое строение листьев у всех изученных видов говорит об адаптации к условиям высокой инсоляции и среднему уровню увлажнения, то есть виды принадлежат к экологической группе световых мезофитов. Мезоморфный облик Р. eriocarpa и Р. barb ata обусловлен в отличие от Р. anisum иными причинами: обитатели весьма жарких и засушливых экотонов первые два вида применяют стратегию избегания неблагоприятных условий иссушения, переходя к эфемерному образу жизни и проходя весь онтогенез при нормальном уровне увлажнения до начала засухи. Р. anisum, давно вошедший в культуру и фактически уже не встречающийся в диком виде, не испытывает недостатка воды на протяжении всего вегетационного сезона, в связи с чем имеет типичную структуру мезофита. Возможно, исходно Р. anisum были свойственны другие фенологические и экологические особенности, которые затем были утрачены в процессе введения в культуру.
В заключении можно еще раз подчеркнуть, что изученные виды имеют разные модели развития, связанные с адаптацией к разным экологическим условиям среды. Р. eriocarpa, Р. barbata принадлежат к группе эфемеров; все этапы онтогенеза у них проходят за относительно короткий период времени (интересно, что все перечисленные особенности проявились у видов и при выращивании в средней полосе, однако при этом сроки прохождения основных этапов онтогенеза сместились на более поздние). Р. anisum относится к группе длительно вегетирующих яровых однолетников. Все три изученных вида имеют световые мезоморфные черты строения, однако Р. eriocarpa, Р. barbata являются как бы «обусловленными» мезофитами.
По морфологии, особенностям онтогенеза и ритму развития ближе друг к другу оказываются Р. eriocarpa и Р. barbata; Р. anisum стоит особняком. Выявленные особенности важны для уточнения систематики рода Pimpinella, а также для практического использования однолетних видов бедренцев.
Вестник ТвГУ. Серия "Биология и экология". 2014. № 2
Об авторах:
Петрова С.Е. Особенности онтогенеза трех однолетних видов рода Pimpinella (Apiaceae) в связи с их экологией / С.Е. Петрова // Вести. ТвГУ. Сер. Биология и экология. 2014. № 2. С. 84-95.
Список литературы Особенности онтогенеза трех однолетних видов рода Pimpinella (Apiaceae) в связи с их экологией
- Онтогенетический атлас растений. М.: Йошкар-Ола. Т. 5, 2007. 372 с.; Т. 6, 2011. 336 с.
- Пименов М.Г., Остроумова Т.А. 2012. Зонтичные (Umbelliferae) России. М.: Т-во научных изданий КМК. 477 с.
- Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах//Тр. Ботан. ин-та АН СССР им. В.Л. Комарова. Сер 3. Геоботаника. М.; Л. Вып. 6. С. 7-204.
- Серебряков И.Г. 1962. Экологическая морфология растений. М.: Высшая шола. 378 с. Чиков П.С. 2002. Лекарственные растения. М.: Медицина. 450 с.
- Шишкин Б.К. 1950. Зонтичные -сем. Umbelliferae Juss.//Флора СССР. М.; Л., Т. 16. 468 с.
- Askari F., Sefidkon F., Meshkizadeh S. 2004. Essential oil composition of Pimpinella eriocarpa Banks & Soland from Iran//Iranian J. Pharm. Res. V. 3. Suppl. 2. № 2. P. 81-82
- Fakhari A.R., Sonboli A. 2006. Essential oil composition of Pimpinella barbata (DC.) Boiss. from Iran//J. Essen. Oil Res. V. 18. № 6. P. 679-681.
- Rechinger K.H. 1987. Umbelliferae//Flora Iranica. B. 162. Austria, Graz: Akadem. Druck-u. Verlagsanst. 499 S.
