Особенности организации популяций высокогорного эндема Anemonastrum biarmiense (Juz.) Holub на территории Южно-Уральского государственного природного заповедника
 Holub на территории Южно-Уральского государственного природного заповедника")
Автор: Юсупова О.В., Абрамова Л.М., Каримова О.А.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 2 (26), 2016 года.
Бесплатный доступ
Представлены результаты изучения онтогенетической структуры ценопопуляций высокогорного эндема Урала Anemonastrum biarmiense (Juz.) Holub на территории Южно-Уральского государственного природного заповедника (ЮУГПЗ). Популяции многочисленные, с плотностью 6-25 экз./м2. Усредненный онтогенетический спектр левосторонний с максимумом на виргинильных особях. Ценопопуляции A. biarmiense нормальные неполночленные. Сравнение индексов восстановления и старения показало, что все популяции являются молодыми, с большой долей прегенеративных растений. Anemonastrum biarmiense на территории ЮУГПЗ представлен реликтовыми популяциями, расположенными в труднодоступной горной местности, которые имеют распространение в подгольцовом поясе, среди альпийских высокотравных лугов, также заходят в горно-тундровую область, произрастая в мохово-лишайниковых сообществах.
Эндем, южно- уральский государственный природный заповедник, ценопопуляции, онтогенетическая структура
Короткий адрес: https://sciup.org/14992819
IDR: 14992819 | УДК: 582.675.1:502.051:502.4
Features of organization of populations of alpine endemic Anemonastrum biarmiense (Juz.) Holub in the South Urals State Natural Reserve
Anemonastrum biarmiense - Ural alpine endemic in a number of places going down to the mountain forest belt, is a rare plant of the Urals. The species is distributed in the upper belts of mountains from the South Urals to the southern part of the Polar Urals. In the South Urals the species is found in all high mountain ridges above the level of the forest boundary (mountains - Iremel, Yamantau, Shatak, ridges - Taganay, Zyuratkul, Nurgush, Urenga, Zigalga, Mashak et al.). In the mountain-forest belt it grows in mountain pine, larch and birch forests, on the steppe slopes, on the banks of mountain rivers (on the ridges Krakow, at the foot of mountains Iremel, in upper courses of the rivers Yuryuzan and Belaya). It is included in the Red Books of the Sverdlovsk region (III category), Komi Republic (II category), Khanty-Mansi Autonomous Area (III category), Yamal-Nenets Autonomous Area (III category), Tyumen region (III category). It was included in the Red Data Book of the Middle Urals (III category). The aim of the study was to reveal the features of ontogenetic structure of coenopopulations of A. biarmiense, growing in the remote mountain area of the South Ural Natural Reserve. In 2015 there were studied: Mashaksky midmountain area of the central part and Eraktashsky mid-mountain area of the southern part of the South Ural State Natural Reserve. 12 coenopopulations of the species are studied in the investigated area. Leading population characteristics were defined: total and effective density of individuals, age structure. Studying of age structure of populations was carried out by standard techniques. L.A.Zhivotovsky’s “delta-omega” criterion based on sharing of indices of age (Δ) and efficiency (ω) was applied to assessment of condition of coenopopulations. In the course of study of this species there was an attempt of differentiation of its place of growth for the mountain-forest and mountain-tundra belt. Coenopopulations 1-7 are located in the mountain-forest belt, coenopopulations 8-12, respectively, in the mountain-tundra belt. Total density in coenopopulations of A. biarmiense, located in the mountain-forest belt varies from 6,9 to 15,3 ind./m2,effective density - 2-6 ind./m2, in mountain-tundra belt the density values vary from 10,3 to 25,0 ind./m2, effective density - 3,6-6,2 ind./m2. In most populations the pregenerative fraction prevails. The averaged ontogenetic spectrum ofA. biarmiense in the mountain-tundra belt is sinistral ancipitous with a maximum in virginal individuals (25,1%), the second peak is in mid-age generative individuals (21,1%); in the mountain-forest belt it is sinistral unimucronate, absolute maximum - in virginal individuals (37,4%). The age structure of concrete coenopopulations of A. biarmiense has two types of spectrum: sinistral and centered. Assessment of age Δ (delta) and efficiency ω (omega) has shown that all coenopopulations of A. biarmiense are young (Δ = 0,11-0,26; ω = 0,33-0,58). Comparison of indices of restoration and aging, reflecting dynamic processes of coenopopulations was carried out. Index of restoration fluctuates from 1,0 to 4,21. This fact testifies to good replenishment by young individuals and prevalence of pregenerative fraction. Index of aging is equal or close to zero that testifies to intensive dying off of individuals in old generative condition.
Текст научной статьи Особенности организации популяций высокогорного эндема Anemonastrum biarmiense (Juz.) Holub на территории Южно-Уральского государственного природного заповедника
Эндемичные таксоны как наиболее оригинальные компоненты любой флоры являются объектами повышенного внимания при флористических и ботанико-географических исследованиях. Изучение эндемичных видов дает ценный материал для познания закономерностей генезиса флоры, путей ее формирования и развития.
Благодаря пограничному положению, большой протяженности с севера на юг и созданному барьерному эффекту, Урал является областью контакта и взаимодействия компонентов различных флор – европейской, сибирской, арктической, ту-ранской. В позднем неогене – антропогене он был областью формообразования для ряда групп растений, следствием чего явилось возникновение флористического эндемизма [1]. Южно-Уральский горный узел занимает второе место по высоте на Урале после Приполярного. В 1979 г. с целью сохранения уникальных природных комплексов Южного Урала – горно-таежных темнохвойных лесов, участков высокогорной растительности и горных болот – был организован Южно-Уральский государственный природный заповедник площадью 2528 км2, расположенный в центральной, наиболее высокой и географически сложной части Южного Урала [2].
Настоящая работа посвящена высокогорному эндему Урала Anemonastrum biarmiense (Juz.) Holub (семейство Ranunculaceae Juss.), входящему в типовую подсекцию рода Anemonastrum (или ряд Narcissiflorae подрода Omalocarpus рода Anemone), которая представляет собой трудную в систематическом отношении группу очень близких между собой викарирующих видов, ранее объединявшихся в один полиморфный вид, – ветреница нарциссоцветная (Anemone narcissiflora L.) [3]. В последнее время для Урала указывают два вида: Anemonastrum biarmiense (ветренник пермский) и A. crinitum (Juz.) Holub (в. косматый) [4]. Последний южносибирский вид представлен на Южном Урале реликтовыми популяциями, имеющими смешанный гибридный характер (наряду с растениями, имеющими типичные для данного вида признаки, произрастают растения с переходными к уральскому виду A. biatmiense признаками). Как считал П.В. Куликов [4], по-видимому, «в реликтовых южноуральских местонахождениях в. косматый в значительной мере оказался поглощен в результате гибридизации с в. пермским, и их совместные популяции представляют собой скопления гибридов, преобладающих над “чистыми” родительскими формами». Аналогичная ситуация наблюдалась и в наших исследованиях. Поэтому мы не разделяли эти два вида, а рассматривали его как Anemonastrum biarmiense в широком понимании и исследовали два типа его местообитаний – в горнотундровом и горно-лесном поясе Южного Урала.
Anemonastrum biarmiense – высокогорный эндем Урала, в ряде мест спускающийся в горнолесной пояс, является редким растением Урала [5, 6]. Психрофит высокогорно-луговой. Ветреник пермский распространен в верхних поясах гор от Южного Урала до южной части Полярного Урала. На Южном Урале вид встречается на всех высоких горных хребтах, превышающих уровень границы леса (горы Иремель, Ямантау, Шатак, хребты Таганай, Зю-раткуль, Нургуш, Уреньга, Зигальга, Машак и др.). В горно-лесном поясе произрастает в горных сосновых, лиственничных и березовых лесах, на остеп-ненных склонах, по берегам горных рек (на хребтах Крака, у подножия гор Иремель, в верховьях рек Юрюзань и Белая). Внесен в Красные книги Свердловской области (III категория) [7], Республики Коми (II категория) [8], Ханты-Мансийского автономного округа (III категория) [9], Ямало-Ненецкого автономного округа (III категория) [10], Тюменской области (III категория) [11]. Был внесен в Красную книгу Среднего Урала (III категория) [12].
Изучение экологии многих видов ветреницы показало [13], что ведущим экологическим фактором, ограничивающим их развитие и распространение, является водный режим почвы. В большом жизненном цикле ветрениц отчетливо выделяются три онтогенетических периода – латентный, вирги-нильный и генеративный. Сенильный период вследствие постоянного обновления подземных органов и значительного вегетативного потенциала растений не выражен [14]. Изученная численность популяций A. biarmiense в Республике Коми показала, что она высокая, и составляет – от 1,6 до 29,0 особей на 1 м² [15, 16]. В Республике Башкортостан исследованы две ценопопуляции на хребте Куркак. Было выявлено, что формируются центрированный и левосторонний спектры [17].
Цель исследования – выявление особенностей онтогенетической структуры ценопопуляций A . biarmiense , произрастающих в труднодоступной горной местности ЮУГПЗ.
Материал и методы
В 2015 г. были обследованы: Машакский среднегорный район центральной части и Еракташ-ский среднегорный район южной части ЮУГПЗ. Далее дана краткая характеристика данных районов.
В Машакский среднегорный район входят наиболее высокие хребты заповедника с гольцовыми вершинами – Зигальга (1427 м), Машак (1382 м), Кумардак (1382 м), Нары (1327 м). Данный район сложен устойчивыми к разрушению породами – кварцитами, межгорные пространства – сланцами и доломитами. Площадь составляет около 1050 км2 (42% площади заповедника). Высотно-поясной ряд ландшафтов образует три типа: горно-таежный, подгольцовый и горно-тундровый. Растительность представлена горнотаежными пихтово-еловыми лесами.
Еракташский среднегорный район расположен южнее Машакского. Включает средневысотные хребты Белятур (1030 м), Нарка (1171 м), Юша (1110 м), Еракташ (1200 м), Капкалка (1186 м). Район характеризуется меньшими абсолютными высотами. Площадь района – 580 км2, сложен кварцитами и песчаниками. Высотно-поясной ряд ландшафтов образует два типа: субгольцовый лесо-луговой (верхний уровень) и горно-таежный темнохвойный (нижний уровень). Растительность субгольцового пояса формируют чередующиеся друг с другом луговые высокотравные сообщества, еловые и березовые низколесья. Средние и нижние части склонов занимают горно-таежные ландшафты, растительность которых образуют коренные пихтово-еловые леса и производные березняки и осинники, встречаются отдельные небольшие массивы лиственничников и сосняков [18].
По А.И. Кайгародову, климат заповедника характеризуется как умеренно континентальный, показатель континентальности – 7 баллов по 10-ти балльной шкале Н.Н. Иванова. Среднегодовая температура воздуха по данным Инзерской метеостанции за период 1930–1992 гг. составила 1,2°– 2,0°С, среднемесячные температуры июля и января соответственно +17°С и -15,8°С. Среднегодовая сумма осадков за 1931–1992 гг. – 667– 616 мм [19, 20]. В распространении почв выражена вертикальная поясность. На наиболее высоких вершинах под горными тундрами формируются горно-тундровые почвы. Ниже на высоте 1000–1200 м на плоских вершинах под субальпийскими лугами и редколесьями распространены горно-луговые почвы. Под лесной растительностью почвенный покров обра- зуют горно-лесные бурые, горные дерново-подзолистые и горно-лесные серые почвы [21, 22].
Объект исследования – Anemonastrum biar-miense – компактнокорневищное многолетнее растение. Корневище укороченное, вертикальное или косо восходящее, утолщенное, покрытое волокнистыми остатками оснований черешков отмерших прикорневых листьев. Прикорневые листья развиваются одновременно с цветоносом, в числе 3-5 или более собранные в розетку, длинночерешковые, с пластинкой в очертании округло-почковидной, сверху почти голой, снизу покрытой рассеянными короткими волосками, рассеченной на три широко-ромбических 2–3-раздельных сегмента 2–5 см длиной и шириной, расположенных на хорошо развитых черешочках от 1,2 до 2 см длиной. Цветоносные стебли пазушные, прямостоячие, 15–65 см высотой, вместе с черешками прикорневых листьев покрытые довольно густыми длинными прямыми горизонтально оттопыренными волосками. На стебле под соцветием имеется обертка (покрывало) из четырех мутовчатых, накрест супротивных сидячих листьев, на 2/3 раздельных, с двумя–тремя надрезанными на верхушке долями. Соцветие зонтиковидное, из двух–шести (8) цветков 1,8-3 см в диаметре, очень редки цветки одиночные. Цветоножки в начале цветения приблизительно равные по длине листьям обертки, позднее в несколько раз длиннее их, при плодах сильно удлиненные, покрытые негустыми тонкими курчавыми волосками. Листочки околоцветника (5–6 шт.) белые, эллиптические или обратнояйцевидные, с обеих сторон голые. Плоды 5– 10 мм длиной, обратнояйцевидные, голые, сильно сплюснутые, окруженные широкой и тонкой крыловидной окраиной, на верхушке с коротким косо отогнутым столбиком. Цветет в мае-июне [23].
В исследуемом районе изучены 12 ценопо-пуляций (ЦП) вида. Название ЦП давалось по ближайшему к ней географическому объекту. Для изучения демографической структуры и плотности ЦП в каждой из них на трансекте закладывалось 30 пробных площадок размером 1 м2. Порядок заложения (линейный или шахматный) и шаг трансекты (1–5 м) зависели от площади, занимаемой конкретной ЦП. Определялись ведущие популяционные характеристики: общая и эффективная плотность особей, возрастной состав.
При определении онтогенетической структуры ЦП, согласно стандартным критериям [24–27], учитывались следующие возрастные состояния: проростки (р), ювенильные (j), имматурные (im), вир-гинильные (v), молодые генеративные (g 1 ), средние генеративные (g 2 ), старые генеративные (g 3 ), субсенильные (ss). На основании полученных данных построены онтогенетические спектры ЦП.
Для характеристики онтогенетической структуры ЦП применяли общепринятые демографические показатели: индекс восстановления [28], индекс старения [29]. Для оценки состояния ЦП был применен критерий «дельта-омега» Л.А. Животов-ского [30], основанный на совместном использовании индексов возрастности (∆) [25] и эффективности (ω) [30].
Результаты и обсуждение
Характеристика местообитаний ценопопуля-ций Anemonastrum biarmiense приведена по результатам проведенных исследований в ЮУГПЗ. В ходе изучения данного вида была попытка разграничения его места произрастания на горно-лесном и горно-тундровом поясе. ЦП 1–7 расположены в горно-лесном поясе и ЦП 8–12 соответственно в горно-тундровом поясе.
ЦП 1 (пик Казабиль) расположена в верхней части склона хр. Нары, в урочище Казабиль, на высоте 1045 м над ур.м., у подножия крупно-глыбовых осыпей. Координаты (по JPS навигатору): с.ш. 54,26666667º, в.д. 57,83388889º. Занимает склон северо-западной экспозиции, с уклоном 10°. Растительность представлена альпийским луговым вы-сокотравьем среди березового редколесья. Общее проективное покрытие (ОПП) травяного яруса составляет 50%. Средняя высота травостоя – 20–70 см. Каменистость почвы составляет 10–20%. Имеется мохово-лишайниковый ярус, развивающийся на камнях. В составе травостоя доминируют Acono-gonon alpinum All. , Bistotra major S.F. Gray , Anemonastrum biarmiense. Кустарниковый ярус слабо развит и представлен Rubus idaeus L.
ЦП 2 (Еракташские поляны) расположена на основной поверхности хребта Юша, у подножия облесенных скал, которые ведут к вершине Торна-таш, на высоте 1020 м над ур.м. Координаты: с.ш. 54,12777778º, в.д. 57,96277778º. Экспозиция склона юго-восточная, с уклоном 10°. Имеется незначительный полог деревьев, где преобладают Betula pubescens Ehrh (h–3,5 m) и Sorbus aucuparia L . (h– 2,5 m), сомкнутость крон – 0,2. Значительную часть растительности представляет луговое высокотра-вье с подгольцовым редколесьем. ОПП травяного яруса составляет 65–70%. Средняя высота травостоя достигает от 70 до 130 см. Каменистость почвы – 40–60%. Доминантами в травостое являются Calamagrostis arundinacea (L.) Roth , Chamerion an-gustifolium (L.) Rafin ., Hieracium albocostatum Norrl. ex Juxip , Bupleurum aureum Fisch ., Anemonastrum biarmiense, Crepis sibirica L. Имеется значительный мохово-лишайниковый ярус, развивающийся на камнях. Кустарниковый ярус представлен Rubus idaeus и Rosa majalis Herm., его ОПП составляет 20–30%.
ЦП 3 (хр. Юша). Ценопопуляция расположена в седловине хр. Юша, разделяющей вершины Каинтюбе и Торнаташ, в верхней части склона, на высоте 777 м над ур.м. с координатами с.ш. 54,12472222º, в.д. 57,92416667º. Экспозиция склона – северо-западная с уклоном 15°. Древесный ярус представлен единичными деревьями Pinus sylvestris L., Larix sibirica Ledeb., Betula pe-ndula Roth., Sorbus aucuparia. Большую часть растительности образует луговое высокотравье. ОПП травяного яруса варьирует в пределах 60–80%. Средняя и максимальная высота травостоя составляет 50– 210 см. В составе травостоя доминируют Potentilla erecta (L.) Raeusch., Anemonastrum biarmiense, Aco-nogonon alpinum, Calamagrostis arun-dinacea, Trollius europaeus L. Каменистость почвы составляет 5– 10%. Кустарниковый ярус представлен Vacci-nium myrtillus L., его ОПП равна 10–25%.
ЦП 4 (хр. Белятур) занимает верхнюю часть остепненного склона южной экспозиции одноименного хребта с уклоном в 30–40°. Координаты: с.ш. 54,14166667º, в.д. 57,90972222º, отметка 967 м над ур.м. На данном участке имеется разреженный полог деревьев, в составе 1-го яруса – Betula pendula и единично Larix sibirica, 2-го яруса – Betula pendula , Sorbus aucuparia , подроста – Picea obovata . Сомкнутость крон составляет 0,4–0,6. Травяной ярус представляют опушечные виды – Origanum vulgare L ., Inula aspera Poir ., Solidago virgaurea L.; степные и петрофитные виды – Dianthus versicolor Fisch. ex Link, Filipendula vulgaris Moench, Phleum phleoides (L.) Karst ., Tephroseris integrifolia (L.) Holub. ОПП меняется от 75 до 90%. Высота растений 70–100 см. Каменистость почвы составляет 30–50%. Кустарниковый ярус представлен Spiraea crenata L. , Chamaecytisus ruthenicus (Fisch.ex Woloszcz.) Klas-kova (dom.), Cotoneaster melanocarpus . ОПП кустарникового яруса варьирует от 45–90%. Мохово-лишайниковый ярус развит не одинаково, его ОПП – 5–25%.
ЦП 5 (Дунан-сунган) находится в верхней части одноименной вершины хр. Юша, в разнотравном луговом сообществе с участием степных видов. Имеет отметку на высоте 943 м над ур.м. с координатами с.ш. 54,06972222º, в.д. 57,88361111º. Занимает южную экспозицию, с относительным уклоном 20°. ОПП травяного яруса составляет 85%, средняя и максимальная высота травостоя – 45– 190 см. Доминируют виды: Potentilla erecta, Pulsatilla patens (L.) Mill. , Trollius europaeus L. , Anemonastrum biarmiense . ОПП кустарникового яруса составляет 15%. Данный ярус представлен Rosa majalis .
ЦП 6 (склон г. Медвежьей) расположена в подгольцовом поясе на хр. Машак, к югу от главной вершины г. Медвежьей, в средней части склона. Занимает северо-западную экспозицию, с уклоном 10–12°, на отметке 1220 м над ур.м. и координатами с.ш. 54,42861111º, в.д. 58,31972222º. Растительность имеет угнетенный характер. Основным сообществом является елово-зеленомошный лес. Выражен древесный ярус, в составе 1-го яруса – Betula czerepanovii Orlova , Picea obovata (доминант); 2-го яруса – Betula tortuosa Ledeb . Сомкнутость крон – 0,3–0,4. В составе травостоя доминируют Festuca austrourelensis Kulikov , Anemonastrum biarmiense, Trientalis europaeus . ОПП травяного яруса составляет 25%. Ярко выражен мохово-лишайниковый ярус, ОПП мохового покрова – 70%, лишайникового – 20%, в составе которого представлены Polytrichum commune, Dicranum polysetum, Cladonia stellaris.
ЦП 7 (Седловина) находится в седловине хр. Машак, разделяющей вершины 1333,6 м и Караульная. Занимает выровненный участок тундроподобного сообщества на высоте 1229 м над ур.м. с координатами с.ш. 54,35972222º, в.д. 58,25166667º. Экспозиция склона – северо-западная. ОПП травяного яруса составляет 60% с доминированием Ane- monastrum biarmiense, Hieracium umbellatum L., Campanula rotundifolia L. В кустарничковом ярусе доминирует Vaccinium myrtillus. Значительное покрытие имеет моховый ярус – 40%, в составе которого представлены Polytrichum commune, Dicranum polysetum.
ЦП 8 (хр. Нараташ) расположена на вершине одноименного хребта, среди скальных осыпей на месте елово-березового криволесья, на высоте 1162 м над ур.м. с координатами с.ш. 54,20194444º, в.д. 57,97083333º. Занимает склон южной экспозиции с уклоном в 45°. Каменистость почвы составляет 90%. Древесный ярус представлен Betula cze-repanovii (h–2,5 m) и Picea obovata (h–3 m), на последнюю приходится основная доля. Сомкнутость крон варьирует от 0,4 до 0,6. Травяной ярус образует петрофитные виды с участием горно-тундровой растительности. ОПП травяного яруса составляет 40–50%, с относительной высотой травостоя 20–50 см. Ярус сложен такими видами, как: Anemonastrum biarmiense, Festuca igoschiniae Tzvel ., Bistorta major, Aster alpinus L ., Asplenium septen-trionale (L.) Hoffm. Кустарники образуют два яруса, с ОПП в 70–90%. Верхний ярус образуют Juniperus communis L ., Cotoneaster melanocarpus Fisch. ex Blytt и нижний – представлен Vaccinium myrtillus (доминант), Vaccinium vitis-idea L. (доминант), Thymus baschkiriensis Klok. et Shost. Имеется развитый мохово-лишайниковый ярус, развивающийся на камнях.
ЦП 9 (г. Медвежья) расположена на одноименной вершине хр. Машак, на месте осоково-лишайниковой тундры, с отметкой 1307 м над ур.м. и координатами с.ш. 54,43694444º, в.д. 58,33138889º. Встречается в гольцовом поясе с выровненной поверхностью почвы и на склоне южной экспозиции с уклоном 5–7°. Каменистость почвы составляет 20%. Незначительно выражена вертикальная структура, где в состав травяного яруса входят: Saus-surea uralensis Lipsch ., Patrinia sibirica (L.) Juss ., Juncus trifidus L. , Anemonastrum biarmiense, Vaccinium uligi-nosum L ., Carex rupestris All. ОПП травяного яруса варьирует от 40 до 50%, средняя высота травостоя – 10–15 см. ОПП мохово-лишайникового яруса составляет 45-60%, в него входят: Cetraria islandica, Cladonia mitis, Cladonia rangiferina.
ЦП 10 (1333,6 м) занимает вершину ветрени-цево-лишайниковой тундры, на одноименной высоте хр. Машак. Имеет координаты: с.ш. 54,35752244º, в.д. 58,22777778º. Описана как на склоне южной экспозиции с уклоном от 8 до 15°, так и на выровненном участке у основания скальных выходов. ОПП травяного яруса составляет 40–60%, высота – 10–25 см. Травяной ярус образуют Anemonastrum biarmiense, Hieracium iremelense Juxip , Festuca igoschiniae, Rhodiola iremelica Boriss . ОПП моховолишайникового яруса составляет 30–60%, с большой долей лишайников – Cladonia mitis, Cladonia rangiferina, Cladonia stellaris, Rhytidium rugosum .
ЦП 11 (г. Безымянная) имеет привязку к горной тундре на вершине г. Безымянной, в южной оконечности хр. Машак, на высоте 1217 м над ур.м. Занимает юго-восточную экспозицию, с координатами с.ш. 54,34527778º, в.д. 58,21333333º, уклон –
18–20°. ОПП травяного яруса составляет 30%, высота – 15-25 см. Доминируют Anemonastrum biar-miense, Juncus trifidus L , Carex vaginata Tausch . ОПП мохово-лишайникового яруса – 80%, с высокой долей лишайников: Cladonia stellaris, Cladonia mittis, Cetraria cuculata, Cladonia amaurocraea, Cladonia rangiferina.
ЦП 12 (1363 м) расположена на одной из вершин хр. Машак, в осоково-лишайниковой тундре, на выровненном участке. Отметка 1363 м над ур.м. Координаты с.ш. 54,33277778º, в.д. 58,22611111º. ОПП травяного яруса 30-55%, высота растений – 10-15 см. Травяной ярус образуют – Carex vaginata (доминант), C. rupestris All. (доминант), Festuca igo-schiniae, Anemonastrum biarmiense, Juncus trifidus, Campanula rotundifolia. ОПП мохово-лишайникового яруса составляет 50-70%, с большой долей лишайников – 70%. В состав входят: Cetraria laevigata, Cladonia amaurocraea, Cladonia mitis, Cladonia rangiferina, Flavocetraria cucullata, Rhytidium rugo-sum .
Растения разных онтогенетических состояний потребляют ресурсы среды разными темпами. Вклад растений разных возрастных состояний в популяционную плотность взвешен соответственно их энергетической эффективности [30]. Общая и эффективная плотность, возрастной состав, демографические показатели представлены в таблице.
Общая плотность в ЦП A. biarmiense, расположенных в горно-лесном поясе, варьирует от 6,9 до 15,3 экз./м2, эффективная плотность – 2–6 экз./м2, в горно-тундровом – значения плотности меняются от 10,3 до 25,0 экз./м2, эффективная плотность – 3,6-6,2 экз./м2. В последнем случае все пять ЦП A. biarmiense являются очень плотными. В большинстве популяциях преобладает прегенера-тивная фракция, лишь в двух ЦП A. biarmiense доли почти равные, приходятся на прегенеративную и генеративную части.
Усредненный онтогенетический спектр A. biarmiense в горно-тундровом поясе левосторонний двухвершинный (рисунок), с максимумом на вирги-нильных особях (25,1%), второй пик приходится на средневозрастные генеративные особи (21,1%); в горно-лесном поясе левосторонний одновершинный, абсолютный максимум на виргинильных осо-
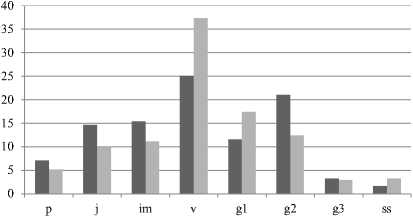
■ Горно-тундровый пояс
■ Горно-лесной пояс
Рис. Усредненные онтогенетические спектры в це-нопопуляциях A. biarmiense.
Распределение особей по онтогенетическим группам и демографические показатели в ценопопуляциях
|
№ ЦП |
к -8- ь CD g |
о С |
Онтогенетическое состояние, % |
Демографические показатели |
|||||||||||
|
р |
j |
im |
v |
g 1 |
g 2 |
g 3 |
ss |
∆ |
ω |
Тип ЦП |
Iв |
Iст |
|||
|
1 |
6,0 |
15,4 |
16,2 |
23,2 |
18,6 |
10,2 |
7,4 |
21,6 |
2,8 |
0,0 |
0,17 |
0,39 |
Молодая |
1,63 |
0,00 |
|
2 |
4,6 |
8,6 |
2,6 |
11,8 |
11,8 |
30,3 |
13,5 |
23,4 |
4,8 |
1,8 |
0,25 |
0,54 |
« |
1,29 |
0,02 |
|
3 |
4,1 |
7,7 |
8,2 |
16,4 |
9,1 |
19,8 |
11,7 |
28,9 |
4,7 |
1,3 |
0,25 |
0,54 |
« |
1,00 |
0,01 |
|
4 |
4,0 |
6,9 |
2,9 |
5,8 |
11,1 |
36,1 |
11,5 |
27,9 |
1,5 |
3,4 |
0,26 |
0,57 |
« |
1,29 |
0,03 |
|
5 |
5,0 |
8,6 |
2,4 |
7,2 |
18,9 |
23,3 |
11,6 |
31,7 |
4,4 |
0,4 |
0,26 |
0,58 |
« |
1,03 |
0,00 |
|
6 |
1,5 |
13,3 |
13,5 |
14,5 |
19,7 |
35,8 |
13,8 |
2,1 |
0,8 |
0,0 |
0,11 |
0,33 |
« |
4,21 |
0,00 |
|
7 |
2,0 |
10,0 |
3,6 |
24,2 |
19,2 |
20,5 |
11,6 |
11,9 |
3,6 |
5,3 |
0,20 |
0,40 |
« |
2,36 |
0,05 |
|
8 |
6,2 |
15,9 |
16,4 |
11,1 |
12,0 |
34,1 |
12,2 |
8,8 |
2,1 |
3,4 |
0,17 |
0,39 |
« |
2,47 |
0,04 |
|
9 |
5,9 |
10,1 |
1,7 |
4,6 |
8,3 |
38,9 |
17,2 |
19,5 |
6,3 |
3,6 |
0,27 |
0,58 |
« |
1,21 |
0,04 |
|
10 |
3,6 |
16,5 |
5,2 |
12,9 |
6,3 |
37,0 |
19,6 |
12,3 |
3,1 |
3,6 |
0,22 |
0,49 |
« |
1,61 |
0,04 |
|
11 |
4,7 |
20,5 |
1,3 |
10,7 |
10,1 |
32,6 |
24,5 |
17,0 |
1,4 |
2,4 |
0,23 |
0,55 |
« |
1,24 |
0,02 |
|
12 |
4,2 |
25,0 |
1,5 |
10,6 |
19,5 |
44,3 |
14,0 |
4,8 |
2,0 |
3,4 |
0,17 |
0,42 |
« |
3,57 |
0,03 |
Примечание: р – проростки, j – ювенильные, im – имматурные, v – виргинильные, g1 – молодые генеративные, g2 – средние генеративные, g3 – старые генеративные, ss – субсенильные.
бях (37,4%). В спектрах представлены растения всех возрастных состояний, кроме сенильного.
По классификации А.А. Уранова и О.В. Смирновой [31], изученные нами ЦП A. biarmiense относятся к нормальным неполночленным. Возрастная структура конкретных ЦП A. biarmiense имеет два типа спектра: левосторонний (ЦП 1, 2, 4, 5, 7–12), центрированный (ЦП 3, 6). Ценопопуляциям 1, 2, 5, занимающим высокотравные альпийские луга, присущ левосторонний двухвершинный спектр. В ЦП 1 пик приходится на ювенильные растения (23,2%), в ЦП 2 и 5 – на виргинильные (30,3 и 36,1%). Второй пик приходится на группу зрелых генеративных растений (21,2–27,9%). Вероятно, данный факт связан с пониженной конкурентоспособностью в высокотравном сообществе. Поскольку растениям необходимо завязать больше плодов для успешного расселения, следовательно, должно быть больше генеративных особей, наряду с прегенеративными. Ценопопуляции с левосторонним одновершинным спектром расположены в подгольцовом поясе, где есть в избытке инсоляция и небольшой полог деревьев, с низким травяным ярусом, занимающие сухие остепненные луга на южной стороне, что отражает наличие благоприятных условий для прорастания семян и успешного прохождения прегене-ративного периода. Центрированный спектр формируется в ЦП 3, 6. Максимум приходится на средневозрастные особи (28,9 и 31,7%).
В ходе исследований была изучена демографическая структура ЦП, которая обеспечивает способность популяционной системы к самоподдержа-нию и определяет ее устойчивость (см. таблицу).
Оценка возрастности ∆ (дельта) и эффективности ω (омега) показала, что молодыми являются все ЦП A. biarmiense (∆ = 0,11–0,26; ω = 0,33–0,58).
Проведено также сравнение индексов восстановления и старения, отражающих динамические процессы ценопопуляций. Индекс восстановления колеблется от 1,0 до 4,21. Данный факт свидетельствует о хорошем пополнении молодыми особями и преобладании прегенеративной фракции. Индекс старения равен или близок к нулю, что говорит об интенсивном отмирании особей в старом генеративном состоянии.
Таким образом, по итогам проведенных исследований можно сказать, что все выявленные ценопопуляции A. biarmiense являются молодыми, с большой долей прегенеративных растений. Популяции многочисленные, с общей плотностью от 6,9 до 25 экз./м2. Ценопопуляции, произрастающие в горно-лесном поясе, более молодые, чем в горнотундровом поясе. Возможно, так сказываются более благоприятные по водно-температурным характеристикам климатические условия, позволяющие семенам вида лучше прорастать, а проросткам – сохраняться в ценопопуляциях. В целом исследованные популяции редких видов находятся в хорошем состоянии, о чем свидетельствуют плотность и структура их популяций.
Список литературы Особенности организации популяций высокогорного эндема Anemonastrum biarmiense (Juz.) Holub на территории Южно-Уральского государственного природного заповедника
- Эндемичные растения Урала во флоре Свердловской области/Науч. ред. В.А. Мухин. Екатеринбург: Гощицкий, 2013. 612 с.
- Горичев Ю.П. Природные особенности Южно-Уральского государственного природного заповедника//Труды Южно-Уральского государственного природного заповедника. Уфа, 2008. С. 13-56.
- Korshinsky S. Tentamen Florae Rossiae orientalis, id est provinciarum Kazan, Wiatka, Perm, Ufa, Orenburg, Samara partis borealis, atque Simbirsk//Зап. Имп. Акад. наук. Физ.-мат. отд. Сер. 8. 1898. Т. 7. Вып. 1. С. 1-566.
- Куликов П.В. Конспект флоры Челябинской области (сосудистые растения). Екатеринбург; Миасс: Геотур, 2005. 537 с.
- Кучеров Е.В., Мулдашев А.А., Галеева А.Х. Охрана редких видов растений на Южном Урале. М.: Наука, 1987. 204 с.
- Горчаковский П.Л., Шурова Е.А. Редкие и исчезающие растения Урала и Приуралья. М.: Наука, 1982. 208 с.
- Красная книга Свердловской области: животные, растения, грибы. Екатеринбург: Изд-во «Баско», 2008. 254 с.
- Красная книга Республики Коми. 2-е изд./Коми НЦ УрО РАН, Сыктывкар, 2009. 800 с.
- Красная книга Ханты-Мансийского автономного округа: животные, растения, грибы. Екатеринбург: «Пакрус», 2003. 376 с.
- Красная книга Ямало-Ненецкого автономного округа: животные, растения, грибы. 2-е изд. Екатеринбург: Изд-во «Баско», 2010. 308 с.
- Красная книга Тюменской области: животные, растения, грибы. Екатеринбург: Изд-во Урал. ун-та, 2004. 496 с.
- Красная книга Среднего Урала (Свердловская и Пермская области). Редкие и находящиеся под угрозой исчезновения виды животных и растений. Екатеринбург: Изд-во Урал. ун-та, 1996. 280 с.
- Папонова И.Т. Некоторые вопросы клональной изменчивости ветреницы алтайской в Центральной части западного Предуралья//Онтогенез травянистых поликарпических растений: Сб. науч. тр. Свердловск: УрГУ, 1986. С. 107-112.
- Барыкина Р.П., Потапова Н.Ф. Биоморфологический анализ видов рода Anemone L. флоры бывшего СССР в ходе онтогенеза//Бюл. МОИП. Отд. биол. 1994. Т. 99. Вып. 5. С. 124-136.
- Вернигор Р.А. Изменчивость и структура популяций высокогорного уральского эндемика ветреницы пермской (Anemone biarmensis Juz.): Автореф. дис.…кан. биол. наук. Свердловск, 1981. 22 с.
- Плотникова И.А. Ветреник пермский//Биология и экология редких растений Республики Коми. Екатеринбург, 2009. С. 86-118.
- Каримова О.А., Жигунов О.Ю., Голованов Я.М., Абрамова Л.М. Характеристика ценопопуляций редких горно-скальных видов в Зауралье Республики Башкортостан//Вестник Томского государственного университета. Биология. 2013. № 2 (22). С. 70-83.
- Горичев Ю.П. Флора и растительность Южно-Уральского государственного природного заповедника. Уфа: Гилем, 2008. С. 67-69.
- Справочник по климату СССР. Вып. 9. Часть 2. Л.: Гидрометеоиздат, 1965. 362 с.
- Жданова Н.В., Лапиков В.В. Климатическая характеристика лесорастительных районов Башкортостана//Биоценотическая характеристика хвойных лесов и мониторинг лесных экосистем Башкортостана. Уфа: Гилем, 1998. С. 60-69.
- Мукатанов А.Х. Горно-лесные почвы Башкирской АССР. М.: Наука, 1982. 147 с.
- Почвы Башкирии. Т.1. Уфа, 1973. 459 с.
- Флора Сибири. Т.6. Portulacaceae -Ranunculaceae/Под ред. Л.И.Малышевой, Г.А.Пешковой. Новосибирск: ВО "Наука". Сибирская издательская фирма, 1993. 310 с.
- Работнов Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах//Тр. БИН АН СССР. Сер. 3. Геоботаника. М.; Л., 1950. Вып. 6. С. 7-204.
- Уранов А.А. Возрастной спектр фитоценопопуляции как функция времени и энергетических волновых процессов//Биол. науки. 1975. №2. С. 7-34.
- Ценопопуляции растений (основные понятия и структура)/О.В. Смирнова, Л.Б. Заугольнова, И.М. Ермакова и др. М., 1976. С.14-43.
- Наумова Л.Г., Злобин Ю.А. Основы популяционной экологии растений/Под. ред. Б.М.Миркина. Уфа, 2009. 88 с.
- Жукова Л.А. Популяционная жизнь луговых растений. Йошкар-Ола: РИИК «Ланар», 1995. 224 с.
- Глотов Н.В. Об оценке параметров возрастной структуры популяций растений//Жизнь популяций в гетерогенной среде. Ч.1. Йошкар-Ола: МарГУ, 1998. С. 146-149.
- Животовский Л.А. Онтогенетическое состояние, эффективная плотность и классификация популяций//Экология. 2001. №1. С.3-7.
- Уранов А.А., Смирнова О.В. Классификация и основные черты развития популяций многолетних растений//Бюл. МОИП. Отд. биол. 1969. Т. 79. Вып. 1. С. 119-135.
