Особенности перехода в состояние зимнего покоя голосеменных и покрытосеменных древесных растений

Автор: Пахарькова Н.В., Гетте И.Г., Андреева Е.Б., Сорокина О.А.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Экология
Статья в выпуске: 6, 2013 года.
Бесплатный доступ
С использованием метода термоиндуцированных изменений нулевого уровня флуоресценции хвои для определения продолжительности и глубины зимнего покоя древесных растений было обнаружено, что хвоя Picea obovata и Abies sibirica гораздо быстрее выходит из состояния зимнего покоя, чем Pinus sibirica, Pinus sylvestris и другие. Повреждение фотосинтетического аппарата хвои Picea obovata может произойти в весенний период, когда после нескольких дней теплой погоды растения преждевременно выходят из состояния зимнего покоя.
Флуоресценция, зимний покой
Короткий адрес: https://sciup.org/14083075
IDR: 14083075 | УДК: 574.24
The peculiarities of winter dormancy state transition of gymnosperm and angiosperm arboreal plants
Using the method of thermally induced changes in the needle fluorescence zero level to determine the duration and depth of arboreal plant winter dormancy it is discovered that needles of Picea obovata and Abies sibirica come out of hibernation faster than Pinus sibirica, Pinus sylvestris and others. The damage of the Picea obovata needle photosynthetic apparatus may occur in spring season, when after a few days of warm weather plants prematurely get out of winter dormancy.
Текст научной статьи Особенности перехода в состояние зимнего покоя голосеменных и покрытосеменных древесных растений
В Сибири основными лесообразующими породами являются пихта сибирская – Abies sibirica Ledeb., ель сибирская – Picea obovata Ledeb., сосна обыкновенная – Pinus sylvestris L., сосна сибирская кедровая – Pinus sibirica Du Tour., лиственница сибирская – Larix sibirica Ledeb., берёза повислая – Betula pendula Roth. и другие. Традиционно регистрацию сезонных изменений у древесных растений проводили фенологическими методами, например, фиксировали время раскрытия почек и развертывания листвы или хвои, связывая это с суммой накопленных положительных температур [2]. Исследования, проводимые на территории государственного заповедника «Столбы» в 1952–2008 годах, позволили выявить у деревьев разных видов временные сдвиги этих фенофаз за два последних десятилетия в среднем на 2–15 суток [3]. В связи с этим несомненный интерес представляют изменения всех фенофаз древесных растений, в том числе трудно диагностируемые периоды осенней фотопериодической реакции и перехода в состояние глубокого зимнего покоя. Известно, что при переходе от фазы активной вегетации к фазе зимнего покоя клетки фотосинтезирующей паренхимы претерпевают ряд изменений, таких как: изменение количества и соотношения пигментов, увеличение доли ненасыщенных жирных кислот в мембранах, повышение концентрации моно- и олигосахаров, синтез криопротекторных белков, увеличение вязкости цитоплазмы, изменение структуры фотосинтетического аппарата [4–7].
Состояние фотосинтетического аппарата (ФСА), играющего важнейшую роль в обеспечении биологической продуктивности растений, является хорошим индикатором, характеризующим состояние растений в целом, а также по состоянию ФСА можно проследить скрытые изменения, проходящие при переходе растений в состояние зимнего покоя и последующем выходе из этого состояния [8].
Цель работы. Изучение различной глубины зимнего покоя и особенностей перехода в это состояние у голосеменных и покрытосеменных древесных растений.
Методы, объекты и район исследования. В качестве района исследования была выбрана территория туристско-экскурсионного района заповедника «Столбы», расположенного на юго-западной окраине г. Красноярска. В годовой динамике температур в этом районе четко выражен достаточно продолжительный холодный период, когда преобладают отрицательные температуры и активная жизнедеятельность растений невозможна. Для исследований использовали двухлетнюю хвою голосеменных (пихты сибирской, ели сибирской, сосны обыкновенной, сосны сибирской) и феллодерму побегов второго года у березы и лиственницы, собранные в сентябре-апреле 2010–2011 и 2011–2012 годов с периодичностью в две недели.
При изучении перехода растений в состояние покоя и выхода из него хорошо зарекомендовал себя метод регистрации кривых термоиндуцированных изменений нулевого уровня флуоресценции (ТИНУФ). Данные ТИНУФ регистрировались на флуориметре «Фотон-11» при нагреве со скоростью 8 градусов в минуту. В качестве показателя глубины покоя использовалось отношение интенсивности нулевого уровня флуоресценции при 50°С и 70°С (коэффициент R2). Значения коэффициента R ниже единицы свидетельствуют о том, что растения находятся в состоянии зимнего покоя, а выше единицы – активно вегетируют [9]. Ранее в наших исследованиях были обнаружены характерные изменения термоиндуцированного ответа флуоресценции хлорофилла в периоды перехода феллодермы и хвои в состояние зимнего покоя и выхода из него [10, 11].
Для дополнительной характеристики фотосинтетического аппарата была прослежена динамика содержания фотосинтетических пигментов. Количество хлорофилла a , b и суммарное содержание каротиноидов были измерены на спектрофотометре SPEKOL 1300 Analytik Jenna AG (в ацетоновой вытяжке в пересчете на сухую массу) [12].
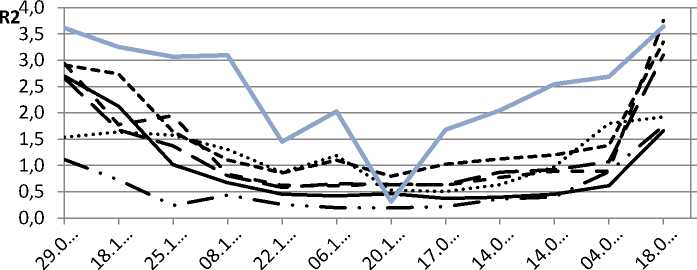
Результаты и их обсуждение. Сравнивая глубину зимнего покоя у различных видов голосеменных и покрытосеменных древесных растений путем анализа кривых сезонных изменений коэффициента R2 (рис.1), можно выделить некоторые закономерности. Среди голосеменных наиболее ранние сроки перехода в состояние покоя, большая глубина покоя и более поздний выход из этого состояния отмечены у сосны обыкновенной. Кроме того, эта кривая наиболее сглажена, т.е. глубина покоя сосны обыкновенной в наименьшей степени зависит от текущей температуры.
Для пихты сибирской, наоборот, характерны наиболее короткие сроки зимнего покоя и наименьшая его глубина, в значительной степени зависящая от температуры воздуха. У ели сибирской и сосны сибирской зарегистрированы промежуточные значения. Так, в 2010–2011 годах коэффициент корреляции значений R2 и текущей температуры воздуха для Pinus sylvestris равен 0,7; для Pinus sibirica – 0,73; для Picea obovata – 0,74 и для Abies sibirica –0,78.
Лиственные деревья умеренного пояса сбрасывают листву на период отрицательных температур, поэтому сравнение зимнего покоя для древесных представителей покрытосеменных с голосеменными (в данном случае – березы повислой с лиственницей сибирской) проводили по параметрам ТИНУФ хлорофилла феллодермы. Нужно отметить, что динамика коэффициента R2 у лиственницы сибирской свидетельствует о значительно меньшей глубине покоя по сравнению с березой повислой. Покой феллодермы лиственницы в наибольшей степени зависит от текущей температуры воздуха, коэффициент корреляции значений R2 и температуры для Larix sibirica равен 0,84, тогда как для Betula pendula – 0,68.
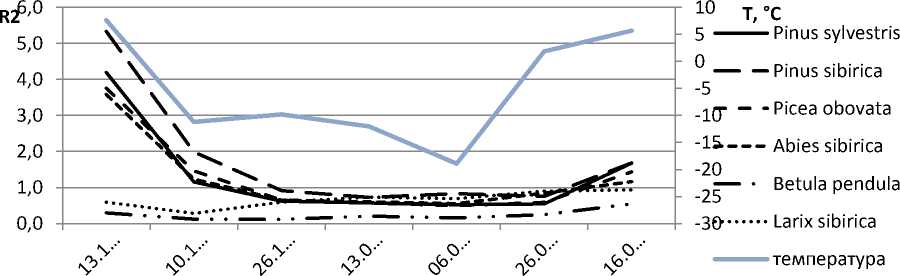
10 T, °C
5 Pinus sylvestris
0 Pinus sibirica
-5
-10 Picea obovata
-15 Abies sibirica
-20
Betula pendula
-25
-30 Larix sibirica
температура
А
Б
Рис. 1. Динамика коэффициента R2 в природных условиях: А – 2010–2011 гг.; Б – 2011–2012 гг.
Осенний период 2011 года характеризовался более ранним снижением температуры и более плавным ходом в течение зимнего периода 2012 года, без резких температурных скачков, которые были отмечены в 2010–2011 году. Кривые коэффициента R2 также имеют более сглаженный вид, все исследуемые виды с конца декабря по конец марта находились в состоянии покоя.
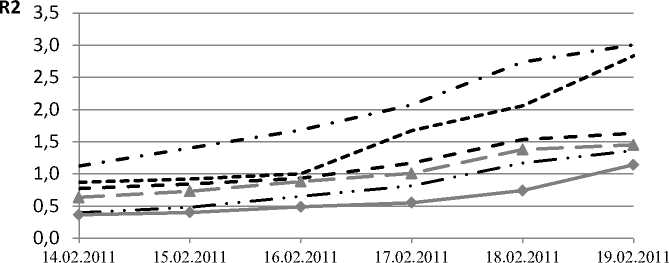
Для получения дополнительной информации побеги растений исследуемых видов искусственно выводили из состояния покоя в лабораторных условиях (при температуре +24°С и 12-часовом фотопериоде). Этот эксперимент проводили в фазе вынужденного покоя во второй половине февраля. В момент сбора образцов температура воздуха в районе исследования составляла около -10°С. Однако известно, что температуры хвои и окружающего воздуха могут не совпадать; так, при минусовых значениях температуры воздуха в солнечный день температура хвои и коры с южной стороны побегов некоторых древесных растений в полдень может достигать +5°С и более [13]. Как свидетельствуют данные, представленные на рисунке 2, побеги Abies sibirica уже находятся в состоянии вегетации.
Pinus sylvestris
Pinus sibirica
Picea obovata
Abies sibirica
Betula pendula
Larix sibirica
Рис. 2. Динамика коэффициента R2 при выведении из покоя в лабораторных условиях
Так же, как и в естественных условиях, наибольшую глубину покоя демонстрирует Pinus sylvestris , чья хвоя начинает вегетировать на четвертые сутки, а средние значения отмечены для Pinus sibirica и Picea obovata , которые выходят из состояния покоя через двое суток.
Медленнее всего из состояния покоя выходит феллодерма Betula pendula – 5 суток, а клетки фелло-дермы Larix sibirica проявляют фотосинтетическую активность через трое суток.
Для дополнительной характеристики фотосинтетического аппарата и оценки его способности к восстановлению фотосинтетической активности была прослежена динамика содержания фотосинтетических пигментов хлорофилла a , b и суммарное содержание каротиноидов в хвое. В целом можно отметить уменьшение содержания хлорофилла в зимний период (рис.3,А) и увеличение количества каротиноидов (рис. 3,Б). В октябре максимальное содержание хлорофиллов характерно для пихты сибирской, продолжающей в это время активно вегетировать. В декабре наименьшее количество хлорофилла имеет сосна обыкновенная, находящаяся в наиболее глубоком покое и потенциально не способная к восстановлению фотосинтеза.
Известно, что фотосинтетический аппарат вечнозеленых хвойных обладает комплексной системой защитных механизмов, которые помогают избегать фотоингибирования в условиях отрицательной температуры. Одной из важных составляющих этой комплексной системы являются каротиноиды, которые стабилизируют мембраны хлоропластов и белки антенных комплексов, поглощают и рассеивают «неиспользуемую» в этих условиях световую энергию [4, 6]. Максимальное содержание каротиноидов в хвое наблюдается в зимний период, но и весной их количество достаточно велико.
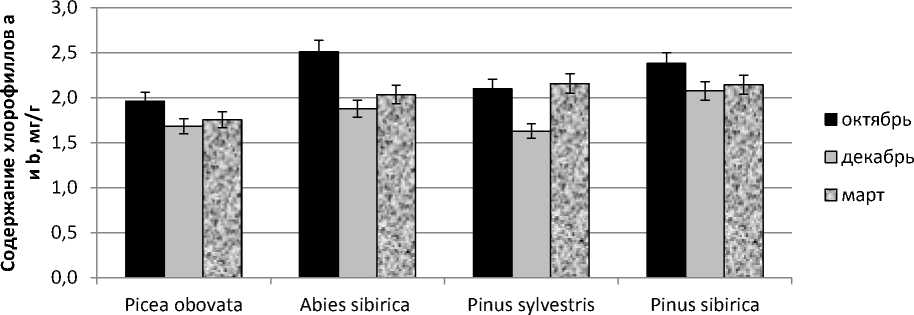
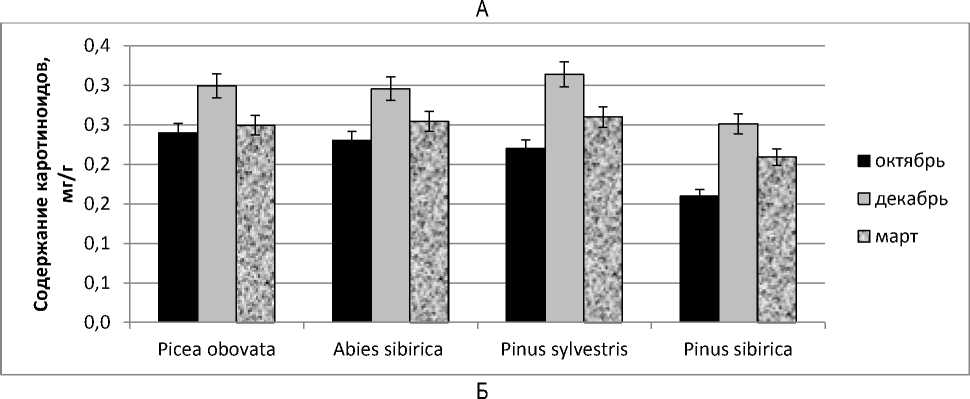
Рис. 3. Содержание пигментов в хвое: А – сумма хлорофиллов а и b;
Б – каротиноиды в зависимости от времени года
По изменению количества каротиноидов в хвое заметны отличия между видами из группы темнохвойных и светлохвойных. Так, в хвое Picea obovata содержание каротиноидов в декабре увеличивается в
1,25 раза по сравнению с октябрем, у Abies sibirica в 1,29 раза, а у Pinus sylvestris и Pinus sibirica в 1,43 и 1,57 раза соответственно, что связано с различными световыми условиями произрастания.
Заключение. Таким образом, мы можем сделать вывод о различной глубине покоя исследуемых древесных растений. Темнохвойные виды, особенно пихта, имеют меньшую глубину покоя и его продолжительность. Возможно, в условиях потепления климата деревья Abies sibirica выходят из состояния вынужденного покоя даже во время кратковременных зимних оттепелей, не характерных ранее для исследуемых районов. Возникающие при возобновлении фотосинтеза и газообмена потери воды не могут быть компенсированы в зимних условиях, что приводит к иссушению и гибели хвои. На наш взгляд, усыхание пихтовых лесов, отмечаемое с конца прошлого века во многих регионах Европы и России, может быть объяснено в том числе и нарушением прохождения стадии зимнего покоя в условиях изменения климата.
