Особенности питания серой жабы Bufo bufo (Anura, Amphibia) в различных биотопах
 в различных биотопах")
Автор: Ручин А.Б., Алексеев С.К.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 2 т.10, 2008 года.
Бесплатный доступ
Изучено питание серой жабы Bufo bufo в различных типах биотопов (широколиственный лес, черно- ольшаник и ельник). В целом пищевой комок состоял из представителей Annelida, Mollusca, Arthropoda. Позвоночные животные в пище не найдены. В широколиственном лесу преобладали Formicidae, Opiliones, Diplopoda, Chilopoda, имаго Carabidae и Staphylinidae. В ельнике и черноольшанике преобла- дающие группы были сходные (Formicidae, Diplopoda, взрослые особи Carabidae и Staphylinidae). Отме- чается склонность жабы к мирмекофагии. Основа пищи - бегающие формы, обитатели герпетобия и стратобия. Наблюдалось изменение в предпочитаемости жертв с увеличением размеров. Более круп- ные особи потребляли значительно больше представителей отрядов Opiliones, Diplopoda, Chilopoda. У мелких особей в пище преобладали небольшие по размеру членистоногие из Aranei, Acarina, Collembola и Ichneumonidae.
Короткий адрес: https://sciup.org/148198113
IDR: 148198113 | УДК: 597.822(282.247.41)
The peculiarity of the feeding common toad Bufo bufo (Anura, Amphibia) in different biotops
A feed of a common toad Bufo bufo in woods of different types is studied. The food consisted of representatives Annelida, Mollusca, Arthropoda. Vertebrate animals are not found in food. In a deciduous wood prevailed Formicidae, Opiliones, Diplopoda, Chilopoda, imago Carabidae and Staphylinidae. In a fir grove and an alder wood prevailing groups were similar (Formicidae, Diplopoda, imago Carabidae and Staphylinidae). Propensity of a toad to eating ants is marked. The basis of food was made with running forms, inhabitants of a surface of ground and a laying. Change in preference of victims with increase in the sizes was observed. Larger individuals consumed much more representatives from groups Opiliones, Diplopoda, Chilopoda. At fine individuals in food small arthropods on the size from such groups prevailed: Aranei, Acarina, Collembola and Ichneumonidae.
Текст научной статьи Особенности питания серой жабы Bufo bufo (Anura, Amphibia) в различных биотопах
В различных экосистемах амфибий можно отнести к одним из основных связующих звеньев в передаче энергии от разных аккумулирующих ее источников на более высокие трофические уровни [17]. В лесных биоценозах земноводные, имеющие высокую численность и биомассу, играют чрезвычайно значительную трофическую роль. Серая жаба – типичный лесной обитатель [2, 3, 7, 15, 16, 19-21]. Однако в ряде местообитаний ее можно встретить и в насаждениях антропогенного характера: рощах, парках, садовых участках, городских скверах, изредка – лесопосадках. Обычно серая жаба избегает открытых участков, хотя изредка ее отмечают в агроценозах [17]. Имея крупные размеры, этот вид является активным потребителем различных видов беспозвоночных [1, 3, 8, 11, 16]. Цель наших исследований – определение питания серой жабы в различных биотопах, а также определение зависимости качественного состава пищевого комка от размеров.
Материал и методы
Сборы проводились в разных биотопах, кратко охарактеризованных ниже.
Широколиственный лес . Характеризовался типичной серой лесной почвой (механический состав почвы: суглинистый). В древесно-кустарниковом ярусе произрастали Fraxinus excelsior L., Quercus robur L., Acer platanoides L., A. campestre L., Tilia cordata Mill., Ulmus glabra Huds., Padus aveum Mill., Salix caprea L., Corylus avellana L., Euonymus. europaea L., E. verrucosa Scop., Lonicera xylosteum L., Frangula alnus Mill. В травянистом ярусе: Aegopodium podagraria L. Galeobdolon luteum Huds., Mercurialis perennis L., Asarum europaeum L., Pulmonaria obscura Dumort., Milium effusum L., Campanula latifolia L., Carex pilosa Scop. и др.
Ельник . Характеризовался дерново-легкоподзолистой почвой среднеглинистого механического состава. В древесно-кустарниковом ярусе произрастали Picea abies (L.) Karst., Corylus avellana L., Sorbus aucuparia L., В травянистом ярусе: Ocsalis acetosella L., Maiantemum bifolium L., Asarum europaeum L., Gymnocarpium dryopterum L.
Черноольшаник . Характеризовался дерново-подзолистой глееватой почвой слабосуглинистого состава. В древесно-кустарниковом ярусе: Alnus glutinosa L., Betula pubescens Ehrh., Salix caprea L.; в травянистом ярусе – Filipendula ulmaria (L.) Maxim, Urtica dioica L., Dryopteris carthusiana Vill., Matteuccia struthiopteris L., Equisetum sylvaticum L., E. palustre L., Epilobium palustre L., Impatiens noli-tangere L., Homulus lupulus L., Chrysosplenium alternifolium L., Ficaria verna Huds., Caltha palustris L.
Для определения размерно-возрастных изменений спектра питания было условно выделено 3 класса особей по длине тела: 1730 мм (это обычно годовики), 31-50 мм (двухтрехлетние особи) и 51-70 мм (более взрослые особи).
Во всех случаях по возможности пищевые объекты определялись до вида. Когда определение было затруднено, объект относили к тому или иному роду или семейству. Использовались обычные определители по беспозвоночным [4, 5, 12, 14]. Рассчитывали относительное количество того или иного объекта в питании. Кроме того, была проведена их дифференцировка объектов питания по степени подвижности и пространственной группировке. При этом за основу подобного разделения взяли монографию [9].
Результаты и их обсуждение
В целом пищевой комок серой жабы состоял из представителей трех типов беспозвоночных: Annelida, Mollusca, Arthropoda. Позвоночные животные в пище не найдены. Однако, как видно из табл. 1, в разных типах леса в пищевом комке серой жабы преобладали различные группы животных. Так, в широколиственном лесу к ним можно отнести Formicidae, Opiliones, Diplopoda, Chilopoda, имаго Carabidae и Staphylinidae, на долю которых приходилось 75,57% от общего количества пищи. Определенную роль в питании жаб в данном типе леса играли Isopoda, Aranei, имаго Curculionidae и Ichneumonidae (11,03%). Именно указанные группы членистоногих и составляли основную часть потребляемых жертв.
В ельнике и черноольшанике преобладающие группы были сходные (Formicidae, Diplopoda, взрослые особи Carabidae и Staphylinidae), доля которых составляла 88,83% и 86,96%, соответственно. Остальные группы беспозвоночных животных в разных биотопах обнаруживались в комке жаб нерегулярно или в небольших количествах. В разных местообитаниях одна особь потребляла примерно одинаковое количество жертв (табл. 1).
Многие авторы [1, 3, 6, 10, 11, 13, 15, 17, 22] указывают для серой жабы ее склонность к мирмекофагии (избирательному поеданию муравьев). Нами также отмечено подобное явление. Причем оно оказалось общим для всех типов леса, но в широколиственном лесу муравьи потреблялись в меньшем количестве в отличие от других биотопов. В этом местообитании жабы «переключались» на жужелиц и стафилинид, которых было довольно много в напочвенном ярусе.
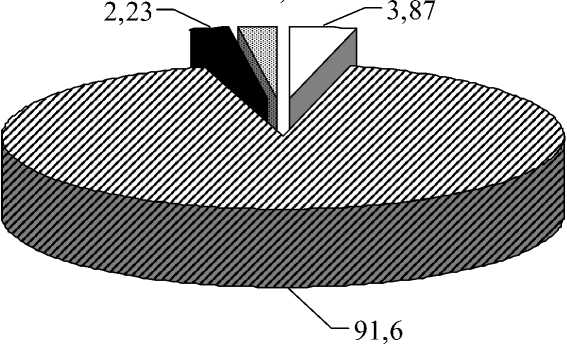
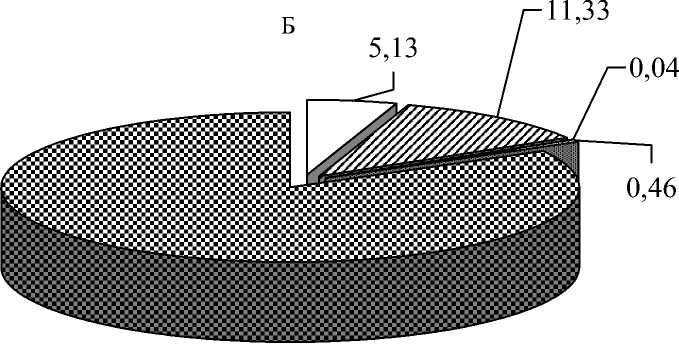
Наибольшую часть пищевого комка составляли бегающие формы, остальные – были представлены немногим числом. Основную часть объектов серая жаба съедает, схватывая с поверхности земли, в связи с чем в пище значителен процент беспозвоночных – обитателей герпетобия и стратобия (рис. 1).
Несмотря на условность выделенных размерных классов, установлены явные различия в их спектрах питания (табл. 2). Так, наблюдалось явное изменение в предпочитае-мости жертв с увеличением размеров. Более крупные особи потребляли значительно больше представителей отрядов Opiliones, Diplopoda, Chilopoda. У мелких особей в пище преобладали небольшие по размеру членистоногие из таких групп: Aranei, Acarina, Collembola и Ichneumonidae. В целом они составляли 42,16% от всех экземпляров добычи.
Таблица 1. Спектр питания серой жабы в разных биотопах (относительное количество, %)
|
Таксон добычи |
Широколиственный лес |
Ельник |
Черноольшаник |
|
1 |
2 |
3 |
4 |
|
Annelida |
|||
|
Oligochaeta |
0,24 |
0,18 |
0,16 |
|
Mollusca |
|||
|
Gastropoda |
0,48 |
1,26 |
0,48 |
|
Arthropoda |
|||
|
Crustacea |
|||
|
Isopoda |
2,54 |
– |
0,32 |
|
Arachnida |
|||
|
Opiliones |
4,00 |
1,26 |
0,32 |
|
Aranei |
2,30 |
1,44 |
1,43 |
|
Acarina |
1,57 |
– |
0,16 |
|
Myriapoda |
|||
|
Diplopoda |
6,30 |
4,32 |
3,18 |
|
Chilopoda |
3,15 |
0,72 |
1,59 |
|
Insecta |
|||
|
Collembola |
0,97 |
– |
0,63 |
|
Homoptera |
|||
|
Auchenorrhyncha |
0,61 |
– |
– |
|
Heteroptera |
1,94 |
0,72 |
0,63 |
|
Coleoptera, l. (неопред.) |
0,24 |
– |
0,16 |
|
Coleoptera, im. (неопред.) |
0,24 |
0,36 |
0,63 |
|
Carabidae, l. |
0,12 |
0,18 |
– |
|
Carabidae, im. |
11,01 |
5,59 |
9,70 |
|
Histeridae, im. |
0,12 |
– |
– |
|
Catopidae, im. |
– |
0,54 |
0,16 |
|
Anisotomidae, im. |
1,57 |
0,18 |
0,48 |
|
Silphidae, l. |
0,73 |
0,18 |
– |
|
Silphidae, im. |
1,33 |
0,36 |
0,48 |
|
Staphylinidae, l. |
0,36 |
0,18 |
– |
|
Staphylinidae, im. |
7,88 |
3,42 |
3,02 |
|
Cantharidae, im. |
0,12 |
– |
– |
|
Scarabaeidae, l. |
– |
– |
0,16 |
|
Scarabaeidae, im. |
0,12 |
– |
0,16 |
|
Phalacridae, im. |
– |
0,18 |
– |
|
Byrrhidae, im. |
0,12 |
0,18 |
– |
|
Buprestidae, im. |
0,12 |
– |
– |
|
Elateridae, l. |
0,12 |
– |
0,16 |
|
Elateridae, im. |
0,48 |
0,54 |
0,95 |
|
Coccinellidae, im. |
0,12 |
– |
– |
|
Pyrochroidae, im. |
0,24 |
– |
– |
|
Cerambycidae, im. |
0,24 |
– |
– |
|
Tenebrionidae, im. |
– |
– |
0,32 |
|
Mycetophagidae, im. |
– |
– |
0,16 |
|
Chrysomelidae, im. |
– |
– |
– |
|
Curculionidae, im. |
2,79 |
1,81 |
0,16 |
|
Hymenoptera, l. (неопред.) |
– |
– |
0,48 |
|
Hymenoptera, im. |
|||
|
Ichneumonidae |
3,40 |
0,36 |
1,11 |
|
Formicidae |
43,23 |
75,50 |
71,06 |
|
Rhaphidioptera, l. |
0,12 |
– |
– |
|
Lepidoptera, l. |
0,36 |
– |
0,63 |
Окончание таблицы 1
|
1 |
2 |
3 |
4 |
|
Diptera, im. |
|||
|
Tipulidae |
0,12 |
0,18 |
0,16 |
|
Brachycera |
0,24 |
0,18 |
0,48 |
|
Diptera, l. |
0,36 |
0,18 |
0,48 |
|
Обработано особей |
17 |
12 |
12 |
|
Количество объектов (общее) |
826 |
555 |
629 |
|
Количество объектов (на особь) |
48,6 |
46,3 |
52,4 |
Примечание: l. – личинки, im. – имаго.
Таблица 2. Спектр питания серой жабы различных размерных классов (относительное количество, %)
|
Таксон добычи |
Размерные классы по длине тела, мм |
||
|
17-30 |
31-50 |
51-70 |
|
|
1 |
2 |
3 |
4 |
|
Annelida |
|||
|
Oligochaeta |
– |
– |
0,21 |
|
Mollusca |
|||
|
Gastropoda |
1,08 |
0,53 |
0,48 |
|
Arthropoda |
|||
|
Crustacea |
|||
|
Isopoda |
– |
1,06 |
1,58 |
|
Arachnida |
|||
|
Opiliones |
0,92 |
1,59 |
2,41 |
|
Aranei |
2,77 |
2,20 |
1,93 |
|
Acarina |
15,23 |
1,15 |
0,96 |
|
Myriapoda |
|||
|
Diplopoda |
0,62 |
6,17 |
4,95 |
|
Chilopoda |
1,54 |
1,50 |
2,47 |
|
Insecta |
|||
|
Collembola |
16,00 |
2,47 |
0,83 |
|
Psocodea |
0,15 |
– |
– |
|
Homoptera |
|||
|
Auchenorrhyncha |
– |
0,09 |
0,34 |
|
Aphidodea |
0,77 |
– |
– |
|
Heteroptera |
1,84 |
0,53 |
1,31 |
|
Coleoptera, l. (неопред.) |
– |
0,09 |
0,14 |
|
Coleoptera, im. (неопред.) |
0,77 |
0,35 |
0,41 |
|
Carabidae, l. |
– |
0,09 |
0,07 |
|
Carabidae, im. |
4,77 |
4,59 |
10,45 |
|
Histeridae, im. |
– |
– |
0,07 |
|
Catopidae, im. |
1,54 |
0,89 |
0,07 |
|
Anisotomidae, im. |
2,31 |
1,06 |
1,10 |
|
Silphidae, l. |
– |
– |
0,41 |
|
Silphidae, im. |
– |
– |
0,96 |
|
Staphylinidae, l. |
– |
0,18 |
0,21 |
|
Staphylinidae, im. |
7,69 |
2,56 |
5,78 |
|
Scydmaenidae, im. |
0,31 |
– |
– |
|
Cantharidae, im. |
– |
– |
0,07 |
|
Scarabaeidae, l. |
– |
– |
0,07 |
|
Scarabaeidae, im. |
– |
– |
0,14 |
|
Phalacridae, im. |
– |
– |
– |
|
Byrrhidae, im. |
– |
– |
0,07 |
|
Buprestidae, im. |
– |
– |
0,07 |
|
Elateridae, l. |
0,31 |
0,18 |
0,14 |
|
Elateridae, im. |
– |
0,09 |
0,69 |
Окончание таблицы 2
|
1 |
2 |
3 |
4 |
|
Cryptophagidae, im. |
0,46 |
0,18 |
– |
|
Coccinellidae, im. |
– |
0,18 |
0,07 |
|
Mycetophagidae, im. |
– |
– |
0,07 |
|
Pyrochroidae, im. |
– |
– |
0,14 |
|
Tenebrionidae, im. |
– |
– |
0,14 |
|
Cerambycidae, im. |
– |
– |
0,14 |
|
Chrysomelidae, im. |
0,31 |
0,26 |
– |
|
Curculionidae, im. |
1,69 |
1,85 |
1,65 |
|
Hymenoptera, l. (неопред.) |
– |
– |
0,21 |
|
Hymenoptera, im. (неопред.) |
– |
0,18 |
– |
|
Ichneumonidae |
8,16 |
1,50 |
2,41 |
|
Formicidae |
|||
|
п/сем. Myrmicinae |
25,38 |
59,96 |
54,86 |
|
п/сем. Formicinae |
– |
7,63 |
0,41 |
|
Rhaphidioptera, l. |
– |
– |
0,07 |
|
Neuroptera, im. |
0,15 |
– |
0,07 |
|
Lepidoptera, l. |
0,77 |
0,09 |
0,48 |
|
Diptera, im. |
|||
|
Tipulidae |
0,31 |
0,09 |
0,14 |
|
Brachycera |
2,46 |
0,62 |
0,34 |
|
Diptera, l. |
1,69 |
0,09 |
0,41 |
|
Обработано особей |
42 |
22 |
29 |
|
Количество объектов (общее) |
650 |
1134 |
1454 |
|
Количество объектов (на особь) |
15,5 |
51,6 |
50,1 |
Примечание: l. – личинки, im. – имаго.
Наибольшую часть пищевого комка составляли бегающие формы, остальные – были представлены немногим числом. Основную часть объектов серая жаба съедает, схватывая с поверхности земли, в связи с чем в пище значителен процент беспозвоночных – обитателей герпетобия и стратобия (рис. 1).
Несмотря на условность выделенных размерных классов, установлены явные различия в их спектрах питания (табл. 2). Так, наблюдалось явное изменение в предпочитае-мости жертв с увеличением размеров. Более крупные особи потребляли значительно больше представителей отрядов Opiliones, Diplopoda, Chilopoda. У мелких особей в пище преобладали небольшие по размеру членистоногие из таких групп: Aranei, Acarina, Collembola и Ichneumonidae. В целом они составляли 42,16% от всех экземпляров добычи.
Из отряда Coleoptera в пищевом комке увеличивалось только количество жужелиц (от 4,77% до 10,45%). Одновременно изменялся и видовой состав этой группы. Мелкие осо- би потребляли в основном небольших или средних по размеру жужелиц следующих видов Trechus secalis Pk., Bembidion sp., Agonum fuliginosum Pz. и др. У средних особей видовой состав жужелиц менялся в сторону более крупных видов Leistus rufescens F., Patrobus excavatus Pk., Badister bullatus Schall., Stomis pumicatus Pz., Pterostichus oblongopynctatus F., Pt. strenuus Pz. и др. У крупных жаб в пище уже преобладали Carabus granulatus L., Pterostichus niger Schall., Pt. oblongopynctatus F., Pt. aethiops Pz., Agonum assimile Pk., A. obscurum Hbst., Synucfus nivalis Pk., Patrobus atrorufus Stroem. и др.
Кроме того, у крупных особей увеличивался видовой диапазон жертв из отряда жуков. Например, у мелких и средних жаб в пищевом комке эта группа была представлена 9 семействами, тогда как у крупных – 17. Причем в пище увеличивались представители с большими размерами из семейств Buprestidae, Cerambycidae, Scarabaeidae, Silphidae.
Как мы отмечали выше, в пище у серой
А
2,3
-
□ ползающие и бегающие ■ прыгающие ■ летающие
83,04
□ филлобий
0 стратобий
н дендробий
и геобий 0 герпетобий
Рис. 1. Соотношение групп организмов в пище серой жабы (n = 73, длина тела – 40-65 мм) от степени подвижности (А) и яруса обитания (Б)
жабы в разных биотопах в значительном количестве встречаются муравьи. Однако можно утверждать, что молодые животные не так охотно съедают муравьев. Эта группа насекомых у небольших жаб составляла всего 25,38% от пищевого комка, тогда как у более крупных особей она доходила до 67,59% и 55,27% (табл. 2). Не исключено, что мирме-кофагия развивается у серой жабы постепенно с увеличением размеров.
Таким образом, в различных типах биотопов в пищевом комке серой жабы преобладали различные группы беспозвоночных животных из типов Annelida, Mollusca, Arthropoda. Как и в других точках ареала, у жабы отмечается склонность к мирмекофагии. Основа пищи – бегающие формы, обитатели герпетобия и стратобия. Более крупные особи потребляли значительно больше представителей отрядов Opiliones, Diplopoda,
Chilopoda. У мелких особей в пище преобладали небольшие по размеру членистоногие из
Список литературы Особенности питания серой жабы Bufo bufo (Anura, Amphibia) в различных биотопах
- Велиева З.Д. О новых находках и экологии серой жабы в Азербайджане//Вопросы герпетологии. Л.: Наука, 1981.
- Гаранин В.И. Материалы по распространению и численности амфибий Волжско-Камского края//Природные ресурсы Волжско-Камского края. М.: Наука, 1964.
- Гаранин В.И. Земноводные и пресмыкающиеся Волжско-Камского края. М.: Наука, 1983.
- Горностаев Г.Н. Насекомые. М.: Изд-во ABF, 1998.
- Горностаев Г.Н. Определитель отрядов и семейств насекомых фауны России. М.: Издательская корпорация «Логос», 1999.
- Дунаев Е.А. Разнообразие земноводных. М.: Изд-во МГУ, 1999.
- Завгородний А.С. Земноводные и пресмыкающиеся (Amphibia et Reptilia) Козельского района Калужской области//Проблемы археологии, истории, культуры и природы Козельского края. Вып. 3-4. Калуга: «Полиграф-Информ», 2001.
- Завгородний А.С., Алексеев С.К., Шашков М.П. Спектры питания массовых видов земноводных (Amphibia) в широколиственных лесах юго-востока Калужской области//Вопросы археологии, истории, культуры и природы Верхнего Поочья. Калуга: Изд-во Н. Бочкаревой, 2001.
- Кузьмин С.Л. Трофология хвостатых земноводных: экологические и эволюционные аспекты. М.: Наука, 1992.
- Кузьмин С.Л. Земноводные бывшего СССР. М.: КМК, 1999.
- Кутенков А.П., Гурулева Е.Л. К экологии серой жабы (Bufo bufo L.) в южной Карелии//Фауна и экология наземных позвоночных. Петрозаводск: Карел. фил. АН СССР, 1988.
- Мамаев Б.М., Медведев Л.Н., Правдин Ф.Н. Определитель насекомых европейской части СССР. М.: Просвещение, 1976.
- Медведев С.И. Материалы к изучению пищи амфибий Северного Донца//Вестн. зоологии. 1974. № 1.
- Негробов О.П., Черненко Ю.И. Определитель семейств насекомых. Воронеж: Изд-во Воронеж. ун-та, 1989.
- Павлов А.В., Замалетдинов Р.И. Животный мир Республики Татарстан. Амфибии и рептилии. Методы их изучения. Казань, 2002.
- Пикулик М.М. Земноводные Белоруссии. Минск: Наука и техника, 1985.
- Писанец Е. Амфибии Украины (спарвочник -определитель земноводных Украины и сопредельных территорий). Киев, 2007.
- Ручин А.Б., Алексеев С.К. К изучению питания остромордой лягушки Rana arvalis в Калужской области//Актуальные проблемы герпетологии и токсинологии. Сб. науч. тр. Вып. 10. Тольятти, 2007.
- Ручин А.Б., Рыжов М.К. Амфибии и рептилии Мордовии: видовое разнообразие, распространение, численность. Саранск: Изд-во Мордов. ун-та, 2006.
- Фомичев С.Н., Коросов А.В. Специфика видовой структуры островного населения амфибий Заонежья//Биоразнообразие Европейского Севера: теоретические основы изучения, социально-правовые аспекты использования и охраны. Петрозаводск, 2001.
- Шкатулова А.П., Карасев Г.Л. Видовой состав и численность фоновых видов земноводных и пресмыкающихся Тюменской области//Безопасность жизнедеятельности в Сибири и на Крайнем Севере. Ч. 1. Тюмень, 1997.
- Antonelli M., Guidali F., Scali S. Alimentazione comparata di tre specie di Amuri in relazione all'habitat//Pianura. 2001. № 13.
