Особенности питания водорослей Nitzschia palea (Kutz.) W. Sm. (Bacillariophyta) и Mychonastes homosphaera (Skuja) Kalina et Punc. (Chlorophyta)
 W. Sm. (Bacillariophyta) и Mychonastes homosphaera (Skuja) Kalina et Punc. (Chlorophyta)")
Автор: Багмет Вероника Борисовна, Абдуллин Шамиль Раисович, Крупская Юлия Валерьевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология
Статья в выпуске: 3-4 т.15, 2013 года.
Бесплатный доступ
В статье рассмотрены особенности питания водорослей Nitzschia palea (Kütz.) W. Sm. и Mychonastes homosphaera (Skuja) Kalina et Punc., выделенных из цианобактериально-водорослевых ценозов пещеры Пропащая яма (Республика Башкортостан). Проведен сравнительный анализ роста штаммов данных видов в темноте и на свету на минеральной и органической среде. Установлено, что при наличии освещенности Nitzschia palea (Kütz.) W. Sm. и Mychonastes homosphaera (Skuja) Kalina et Punc. питаются автотрофно, при ее отсутствии переходят на гетеротрофный тип питания.
Тип питания, автотрофия, гетеротрофия
Короткий адрес: https://sciup.org/148201959
IDR: 148201959 | УДК: 581.1
The nutrition peculiarities of algae Nitzschia palea (Kutz.) W. Sm. (Bacillariophyta) and Mychonastes homosphaera (Skuja) Kalina et Punc. (Chlorophyta)
The nutrition peculiarities of algae Nitzschia palea (Kütz.) W. Sm. and Mychonastes homosphaera (Skuja) Kalina et Punc. from cyanobacterial-algal cenoses of Propashchaya Yama cave (Republic of Bashkortostan) are considered in the paper. Comparative analysis of these species strains growth in light and in darkness on mineral and organic medium was made. It was revealed, that Nitzschia palea (Kütz.) W. Sm. and Mychonastes homosphaera (Skuja) Kalina et Punc. in light feed on autotrophically, in darkness - heterotrophically.
Текст научной статьи Особенности питания водорослей Nitzschia palea (Kutz.) W. Sm. (Bacillariophyta) и Mychonastes homosphaera (Skuja) Kalina et Punc. (Chlorophyta)
В природных экосистемах концентрация минеральных и органических ресурсов, а также уровень освещенности изменяются как во временном, так и в пространственном масштабах. Большинство водорослей имеют автотрофный тип питания, хотя известны виды, которым свойственно миксотрофное и гетеротрофное питание.
Одним из наиболее легкоусвояемых водорослями органических субстратов являются свободные моносахариды. Способность к потреблению этих субстратов отмечена у представителей почти всех классов водорослей [3]. В исследованиях Н.С. Тух-ман с соавт. [4] было выявлено, что многие диатомовые водоросли, в том числе и Nitzschia palea , в условиях слабого освещения способны усваивать ацетат, лактат, этанол, насыщенные жирные кислоты, глицерин, аминокислоты, а также глюкозу. Некоторые виды рода Mychonastes также способны к росту в темноте, используя глюкозу в качестве единственного источника энергии и углерода [6].
Вопрос о питании водорослей в условиях отсутствия освещения, включая и пещеры, до настоящего времени остается полностью не выясненным, поэтому изучение данного вопроса является актуальным.
Целью данной работы было определить возможность диатомовой водоросли Nitzschia palea (Kütz.) W. Sm. и зеленой водоросли Mychonastes homosphaera (Skuja) Kalina et Punc. использовать экзогенные сахара (глюкозу) в качестве источника энергии на свету и в темноте, а также провести сравнительный анализ их скорости роста при разных условиях освещенности.
МАТЕРИАЛ И МЕТОДЫ
Для эксперимента использовали штамм Nitz-schia palea , выделенный из цианобактериальноводорослевого ценоза донных отложений озера
Брехунов пещеры Пропащая Яма (Бурзянский район, Республика Башкортостан) с начальной концентрацией клеток 1,24∙106 в см3 и штамм Mychonastes homosphaera , выделенный из цианобактериальноводорослевого ценоза стены недалеко от входного колодца на дне пещеры Пропащая яма (Бурзянский район, Республика Башкортостан) с начальной концентрацией клеток 7,02∙106 в см3. Предварительно, перед началом эксперимента штаммы были очищены от грибов и бактерий с помощью антибиотика нистатина [2]. Опытным путем была подобрана наиболее эффективная концентрация антибиотика: для Nitzschia palea – 20-25 ЕД /мл, для Mychonastes homosphaera – 80 ЕД /мл. Штаммы культивировали на среде Громова № 6 [1], диатомовую водоросль – с добавлением силиката натрия, в опытную среду дополнительно добавили 1% р-р глюкозы. Подсчет клеток осуществлялся при помощи камеры Горяева в течение десяти дней культивирования.
Полученные данные были обработаны с помощью пакета программ Statistica 8.0 и Microsoft Office Excelle 2007.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Было выявлено, что в начале опыта на свету в контроле (СК) наблюдается активное увеличение числа клеток Nitzschia palea , однако после восьмого дня размножение замедляется. Возможно, это связано с уменьшением концентрации минеральных веществ в среде. Различия в численности клеток между контролем на свету и опытом в темноте (ТГ) (р<0,014), а также с контролем в темноте (ТК) (р<0,007) на протяжения всего опыта были достоверными, различия с опытной средой на свету (СГ) стали достоверными с третьего дня (р=0,000) (рис. 1, табл. 1).
В СГ размножение идет крайне медленно, но сокращения численности клеток не наблюдается. Различия в численности клеток между СГ и ТГ стали достоверными лишь на восьмой день (р=0,000) (рис. 1, табл. 1).
Известия Самарского научного центра Российской академии наук. 2013. Т. 15, № 3 (4)
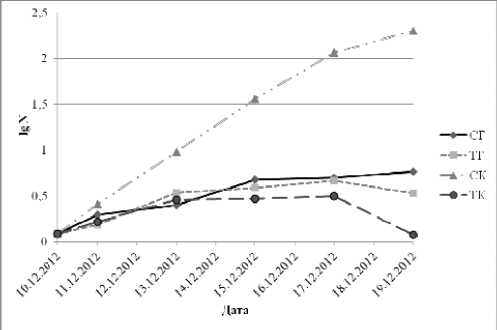
Рис. 1. Динамика изменений числа клеток Nitzschia palea (Kütz.) W. Sm. при разных условиях культивирования.
Прим.: N – число клеток; СК – свет, контроль; СГ – свет, глюкоза; ТК – темнота, контроль; ТГ – темнота, глюкоза
Таблица 1 . Скорость роста клеток Nitzschia palea (Kütz.) W. Sm. при разных условиях культивирования (кл./час)
|
10.12. 2012 |
11.12. 2012 |
13.12. 2012 |
15.12. 2012 |
17.12. 2012 |
19.12. 2012 |
|
|
СК |
0 |
0,056 |
0,143 |
0,568 |
1,629 |
1,806 |
|
СГ |
0 |
0,032 |
0,010 |
0,048 |
0,005 |
0,019 |
|
ТК |
0 |
0,016 |
0,026 |
0,001 |
0,005 |
-0,041 |
|
ТГ |
0 |
0,013 |
0,040 |
0,009 |
0,015 |
-0,026 |
Прим.: обозначения как на рис. 1
В ТК скорость размножения клеток самая низкая. На восьмой день численность клеток резко падает. Различия в численности клеток между ТК и СГ в первый день были достоверными (p<0,036), однако с третьего по шестой день показали недостоверность (p>0,05). Но в последующие дни различия стали более явными и достоверными (р<0,037) (рис. 1, табл. 1).
В ТГ численность клеток почти сравнима с СГ, и значительно выше, чем в ТК. Различия в численности клеток между ТГ и ТК не достоверны лишь в первый день (р=0,885), в остальные дни результаты были достоверными (р<0,016) (рис. 1, табл. 1).
За весь период проведения опыта в СК наблюдается активное увеличение числа клеток Mychonastes homosphaera , (ежедневно почти в 2 раза). Различия в численности клеток между СК и ТК на протяжения всего опыта были достоверными (р<0,001), различия с СГ стали достоверными с шестого дня (р=0,000),а между СК и ТГ стали достоверными на четвертый день (р<0,001) (рис. 2, табл. 2).
В ТК скорость размножения клеток Mychonastes homosphaera самая низкая. Различия в численности клеток между ТК и СГ были на протяжения всего опыта достоверными (p<0,001) (рис. 2, табл. 2).
В ТГ численность клеток выше, чем в ТК. Различия в численности клеток между ТГ и ТК были достоверными на протяжения всего опыта (р<0,001) (рис. 2, табл. 2).
Различия в численности клеток Mychonastes homosphaera между СГ и ТГ стали достоверными на четвертый день (р<0,001) (рис. 2, табл. 2).
Скорость потребления глюкозы, как правило, зависит от уровня освещенности. У некоторых видов свет инактивирует транспортную систему, обеспечивающую поступление глюкозы в клетки [5]. У Nitzschia palea , как и у Mychonastes homosphaera , в органической среде на свету размножение идет медленно, но сокращения численности клеток не наблюдается (рис. 1, 2, табл. 1, 2). Возможно, в нашем случае, у водорослей на свету также происходит инактивация транспортной системы.
Гетеротрофная утилизация органических субстратов у водорослей является важной стратегией выживания, когда уровень света слишком низок для фотосинтеза, или освещенность вообще отсутствует. Поэтому, на наш взгляд, можно говорить о том, что изученные нами штаммы водорослей из пещеры Пропащая яма в условиях темноты питаются гетеротрофно.
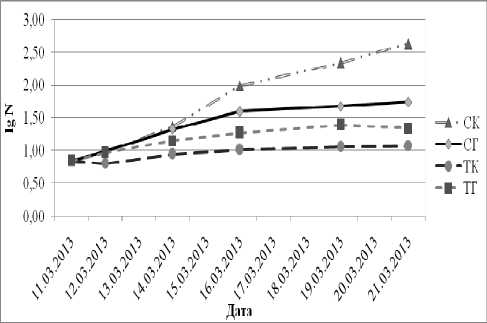
Рис. 2. Динамика изменений числа клеток Mychonastes homosphaera (Skuja) Kalina et Punc. при разных условиях культивирования.
Прим.: N – число клеток; СК – свет, контроль; СГ – свет, глюкоза; ТК – темнота, контроль; ТГ – темнота, глюкоза
Таблица 2 . Скорость роста клеток Mychonastes homosphaera (Skuja) Kalina et Punc. при разных условиях культивирования (кл./час)
|
11.03. 2013 |
12.03. 2013 |
14.03. 2013 |
16.03. 2013 |
19.03. 2013 |
21.03. 2013 |
|
|
СК |
0 |
0,105 |
0,289 |
1,561 |
2,503 |
4,546 |
|
СГ |
0 |
0,109 |
0,243 |
0,381 |
0,169 |
0,166 |
|
ТК |
0 |
-0,028 |
0,055 |
0,027 |
0,025 |
0,006 |
|
ТГ |
0 |
0,096 |
0,103 |
0,090 |
0,130 |
-0,060 |
Прим.: обозначения как на рис. 2
Таким образом, изученные штаммы водорослей Nitzschia palea (Kütz.) W. Sm. и Mychonastes homos-phaera (Skuja) Kalina et Punc. могут рассматриваться как миксотрофы, что является адаптацией к обитанию в пещерах.
Список литературы Особенности питания водорослей Nitzschia palea (Kutz.) W. Sm. (Bacillariophyta) и Mychonastes homosphaera (Skuja) Kalina et Punc. (Chlorophyta)
- Громов Б.В. Коллекция культур водорослей Биологического института Ленинградского университета//Труды Петергоф. биол. ин-та ЛГУ. Л., 1965. Т. 19. С. 125-139.
- Сиренко Л.А., Сакевич А.И., Осипов Л.Ф., Лукина Л.Ф., Кузьменко М.И., Козицкая В.Н., Величко И.М., Мыслович В.О., Гавриленко М.Я., Арендарчук В.В., Кирпенко Ю.А. Методы физиолого-биохимического исследования водорослей в гидробиологической практике. Киев: Наукова думка, 1975. 248 с.
- Успенская В.И. Экология и физиология питания пресноводных водорослей. М.: Изд-во МГУ, 1966. 123 с.
- Tuchman N.C., Schollett M.A., Rier S.T., Geddes P. Differential heterotrophic utilization of organic compounds by diatoms and bacteria under light and dark conditions//Hydrobiologia. 2006. V. 561. P. 167-177.
- Neilson A.H., Lewin R.A. The uptake and utilization of organic carbon by algae; an essay in comparative biochemistry//Phycologia. 1974. V. 13. P. 227-264.
- Yantao Li, Junchao Huang, Gerhard Sandmann, Feng Chen. Glucose sensing and the mitochondrial alternative pathway are involved in the regulation of astaxanthin biosynthesis in the dark-grown Chlorella zofingiensis (Chlorophyceae)//Planta. 2008. V. 228. Is. 5. P. 735-743.
