Особенности полиморфизма по признаку striata в популяциях озерной лягушки Rana ridibunda Pallas, 1771 (Anura, Amphibia) Республики Башкортостан
 Республики Башкортостан")
Автор: Файзулин Александр Ильдусович, Зарипова Фалия Фуатовна, Хусаинова Ильнара Миргалейтовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 3-1 т.15, 2013 года.
Бесплатный доступ
В 2006 – 2012 гг. исследован полиморфизм по признаку striata популяций озерной лягушки ( Rana ridibunda Pallas, 1771) на территории Республики Башкортостан для районов со средним – Предуралье и высоким – Зауралье природным фоновым содержанием тяжелых металлов в среде (меди, цинка, кадмия). Установлено, что в условиях техногенного загрязнения особи с признаком striata отличаются меньшим накоплением тяжелых металлов в Предуралье – меди в 1,9, кадмия в 1,4, цинка в 1,2 раза; для Зауралья – кадмия в 12,8, меди в 1,9, свинца в 1,6 раз.
Фенетическая структура, популяции, озерная лягушка, антропогенные воздействия, республика башкортостан
Короткий адрес: https://sciup.org/148201797
IDR: 148201797 | УДК: 591.53:597.6
Polymorphism on the basis of striata in the populations of a marsh frog Rana ridibunda Pallas, 1771 (Anura, Amphibia) in the Republic of Bashkortostan
Polymorphism on the basis of striata in the populations of a marsh frog Rana ridibunda Pallas, 1771 was investigated in 2006-2012 in the Republic of Bashkortostan in the areas with an average concentration of heavy metals – Cis-Urals, and Trans-Urals - with a high natural background concentration of heavy metals (copper, zinc, cadmium). It was found that in industrial pollution frogs with a sign striata accumulate less concentration of heavy metals in Cis-Urals – copper in 1,9, cadmium in 1,4, zinc in 1,2 times; in Trans-Urals - cadmium in 12,8, copper in 1,9, plumbum in 1,6 times.
Текст научной статьи Особенности полиморфизма по признаку striata в популяциях озерной лягушки Rana ridibunda Pallas, 1771 (Anura, Amphibia) Республики Башкортостан
У большого числа представителей рода Rana встречаются особи с дорсомедиальной полосой, обозначаемой как морфа или фенотип «striata» [27, 17, 43]. Наиболее изучены особенности полиморфизма по признаку striata у озерной Rana ridibunda Pallas, 1771 и остромордой R. arvalis Nilsson, 1842 лягушек. Наличие признака «striata» определяет доминантный аллель диаллельного аутосомного гена, что установлено для озерной [40] и остромордой [39] лягушек. Для полосатой морфы остромордой лягушки выявлены следующие физиологические особенности. С.С. Шварц и В.Г. Ищенко [36] указывают на высокую чувствительность особей морфы striata у R. arvalis к «за-морным» явлениям и их более высокую энергоемкость. Также у сеголетков R . arvalis установлено более высокое выделение СO 2 в единицу времени на 1 г массы у striata по сравнению с беспо-лосыми морфами выше в 1,5-2 раза [7]. Данная особенность обусловливает у них высокую миграционную способность и засухоустойчивость [17]. В другой работе [25] показано, что сеголетки полосатой морфы у остромордой лягушки обладают низкой чувствительностью к тироксину, связанную с исходно высоким уровнем окислительновосстановительных процессов. В целом отмечено более раннее половое созревание и короткая общая продолжительность жизни у полосатых особей [22]. Исследование натриевой проницаемости кожи R. arvalis [6] показало существенное ее сни-
жение (более чем в 3 раза) у полосатых особей по сравнению с бесполосыми. Данные физиологические особенности приводят к усилению легочного дыхания, что обуславливает высокое содержание железа в организме полосатых особей остромордой лягушки [4, 5]. Многочисленные исследования на разных видах рода Rana показали, что полосатые особи обладают большей массой тела и печени [36], также для них характерен повышенный обмен веществ [7], понижена проницаемость кожи [6]. Отмечаются также различия по накоплению тяжелых металлов для особей с различными фенотипами у остромордой [35] и у озерной [42] лягушек.
Распределение доли полосатых и бесполосых особей также связывают с особенностями местообитаний – наличием водно-воздушной растительности (где проявляется криптическое значение дорсомедиальной полосы), а также скорости течения водотока [20].
По литературным данным наблюдается зависимость проявления полиморфизма от степени антропогенного воздействия [3, 8–10, 16, 19, 23, 24, 26, 28–31, 34, 37, 38, 42]. Следует отметить, что для соотношения фенотипов striata и non-striata наблюдается сезонная изменчивость [20, 38]. При обитании земноводных в условиях загрязнения данные физиологические различия между полосатыми и бесполосыми особями могут быть причиной адаптивного преобладания фенотипа striata в популяциях озерной лягушки [2–5, 24].
Изучены особенности географического распределения встречаемости полосатых морф у зеленых лягушек [27, 41]. Полиморфизм по признаку «striata» и по комплексу признаков рисунка и окраски исследовался как в Республике Башкортостан [16], так и в сопредельных регионах: Республике Татарстан [10–12], Челябинской [34] и
Самарской [29, 31, 32, 42] областях. В других регионах России цветовой полиморфизм по данному признаку анализировался в Калужской [28], Тамбовской [20], Волгоградской [21] областях.
Цель работы – проанализировать особенности проявления полиморфизма по признаку striata в популяциях озерной лягушки в условиях Республики Башкортостан, а также выявить особенности аккумуляции тяжелых металлов полосатыми и бесполосыми морфами.
МАТЕРИАЛ И МЕТОДИКА
В качестве объекта исследования выбрана озерная лягушка – наиболее устойчивый к антропогенной трансформации местообитаний вид амфибий, населяющий в том числе и урбанизированные территории [14]. Анализ полиморфизма проводился по соотношению числа полосатых (морфа «striata») (рис. 1а) и бесполосых (морфа «non-striata») (рис. 1б) особей в популяции.


а)
б)
Рисунок 1. Варианты окраски спины озерной лягушки: а) полосатая; б) бесполосая.
Сбор материала проводился с 2006 по 2012 годы в Предуралье и Зауралье Республики Башкортостан (рис. 2) в 15 географических пунктах: 1. «Кама» – залив Курья р. Кама Краснокамского района; 2. «Затон» – парк «Волна», микрорайон Затон г. Уфа; 3. «Теплое» – оз. Теплое, Инорс, Калининский район, г. Уфа; 4. «Баязитово» – дер. Баязитово Альшеевского района, р. Тюлянь; 5. «Алкино» – пос. Алкино Чишминского р-на, р. Дема; 6. «Локотки» – с. Локотки, Уфимского района, оз. Большой Улукуль; 7. «Нагаево» – г. Уфа, дер. Нагаево, озера поймы р. Белая; 8. «Салават» – г. Салават, пруд в парке им. 50 лет Октября; 9. «Белая» – г. Салават, р. Белая (пляж у моста); 10. «Сабашево» – дер. Сабашево Мелеузовского района, р. Белая; 11. «Ишимбай» – г. Ишимбай, р. Белая (пляж); 12. «Исяново» – с. Исяново Баймак-ского р-на, пруд у оз. Талкас; 13. «Таналык» – г. Баймак, р. Таналык; 14. «Казанка» – г. Сибай, пос. Казанка, р. Худолаз; 15. «Худолаз» – г. Сибай, городская плотина, р. Худолаз.
Оценку антропогенного воздействия проводили по результатам химического анализа проб воды из мест обитания озерной лягушки по содержанию тяжелых металлов (Cu, Zn, Cd, Pb) и дру- гих показателей. Анализы проводились атомно– абсорбционным методом на аппарате Contrl A (Германия) в центральной лаборатории Сибай-ского филиала ОАО «Учалинский горнообогатительный комбинат», а также в испытательном лабораторном центре филиала ФГУЗ «Центр Гигиены и Эпидемиологии в РБ» г. Учалы. Анализ состояния биотопов озерной лягушки представлен в таблице 1.
По содержанию тяжелых металлов в воде обследованные местообитания озерных лягушек подразделены на 4 группы с критическим, высоким, средним и низким (относительно фона) содержанием тяжелых металлов.
Из таблицы 1 следует, что основными загрязнителями водоемов в регионе исследования являются медь, цинк и кадмий. По ранее опубликованным данным о накоплении тяжелых металлов в организме основным биотоксикантом является кадмий, аккумулирующийся в печени озерной лягушки [15].
Анализ накопления в коже меди, цинка, свинца, кадмия выполнен в центральной лаборатории СФ ОАО «УГОК» атомно-абсорбционным методом с расчетом по принятой методике .
Таблица 1. Характеристика местообитаний озерных лягушек по содержанию тяжелых металлов в водоемах для периода исследования
|
№ |
Географический пункт |
Содержания тяжелых металлов мг/л |
Степень загрязнения |
|||
|
по Cu |
по Zn |
по Pb |
по Cd |
|||
|
1 |
Кама |
- |
- |
- |
- |
низкий |
|
2 |
Затон |
0,076 1 |
0,123 1 |
0,000036 |
0,083 1,2,3 |
критический |
|
3 |
Теплое |
0,061 1 |
0,078 1 |
0,00024 |
0,018 1,2,3 |
критический |
|
4 |
Баязитово |
0,015 1 |
0,038 1 |
0,000012 |
0,0007 |
низкий |
|
5 |
Алкино |
0,007 1 |
0,023 1 |
0,000012 |
0,0002 |
низкий |
|
6 |
Локотки |
0,041 1 |
0,063 1 |
0,00003 |
0,022 1,2,3 |
высокий |
|
7 |
Нагаево |
0,006 1 |
0,037 1 |
0,00015 |
0,0025 2 |
средний |
|
8 |
Салават |
0,008 1 |
0,023 1 |
0,0005 |
0,0004 |
средний |
|
9 |
Белая |
0,018 1 |
0,039 1 |
0,00066 |
0,001 2 |
высокий |
|
10 |
Сабашево |
0,009 1 |
0,03 1 |
0,0005 |
0,0001 |
низкий |
|
11 |
Ишимбай |
0,02 1 |
0,05 1 |
0,00222 |
0,0016 2 |
высокий |
|
12 |
Исяново |
0,009 1 |
0,026 1 |
0,00001 |
0,0009 |
низкий |
|
13 |
Таналык |
0,0234 1 |
0,447 1,3 |
0,000366 |
0,0029 2 |
критический |
|
14 |
Казанка |
0,0026 1 |
0,032 1 |
0,00001 |
0,0003 |
средний |
|
15 |
Худолаз |
0,0213 1 |
0,379 1 |
0,00001 |
0,003 2 |
высокий |
Примечание: 1ПДК рыбохозяйственный (Россия); 2ПДУ рекреационных водоемов (Россия); 3ПДК рыбохозяйственный (Италия).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Соотношение морф striata и non-striata [1] в урбанизированных биотопах и естественных популяциях амфибий на территории Республики Башкортостан представлено на рис. 2.
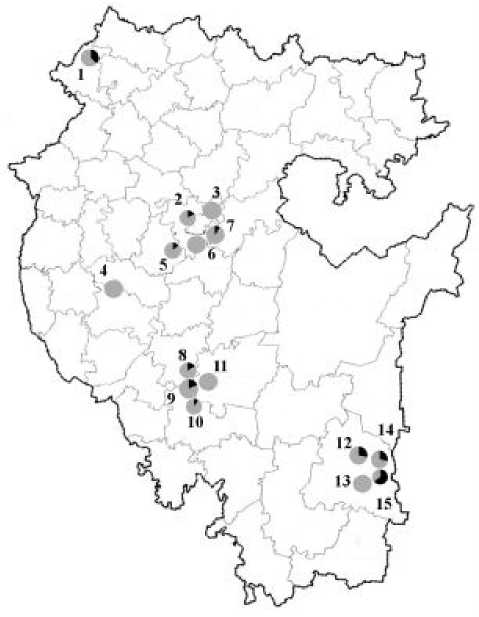
Рисунок 2. Доля полосатых и бесполосых особей озерных лягушек в антропогенных и контрольных участках.
Примечание: 1. Кама (n = 16); 2. Затон ( n = 30); 3. Теплое (n = 28); 4. Баязитово (n = 12); 5. Алкино (n =
40); 6. Локотки (n = 20); 7. Нагаево (n = 20); 8. Салават (n = 51); 9. Белая (n = 45); 10. Сабашево (n = 35); 11. Ишимбай (n = 19); 12. Исяново (n = 25); 13. Та-налык (n = 31); 14. Казанка (n = 48); 15. Худолаз (n = 121). Цветом выделены: черным – доля полосатых особей (striata %), серым – доля бесполосых особей.
Как видно из рисунка 2, морфа «striata» отсутствует в 5 популяциях. Редки с частотой до 20% полосатые особи в 6 популяциях. Средняя частота встречаемости от 25 до 40%% зарегистрирована в 3, а доминируют с частотой от 50 до 75%% только в 1 популяции. Анализ соотношения полосатых и бесполосых особей в двух зонах республики показал высокую долю особей с морфой striata в зоне Зауралья (20,6%), а в Предуралье их доля насчитывает 8,8%. На бесполосых особей приходится 17,6 и 53,0% соответственно. Распределение фенотипов среди озерных лягушек можно объяснить географической изменчивостью встречаемости признака «striata» в популяциях озерной лягушки.
По существующим данным доля особей с признаком «striata» снижается к востоку Республики Татарстан [11, 12] и северо-востоку Самарской области (наши данные) от центральной и западной части региона [29, 31-33]. По данным Е.А. Чибилева [34], доля полосатых особей превышает 50% только в юго-восточной части региона. Следует отметить, что восточнее Башкортостана в г. Челябинск доля морфы striata изменяется от 98% (очистные сооружения) и 46–57%% (пруд Коммунар) до 10% (Шершеневское водохранилище) [34].
В отличие от района исследования в других регионах отмечается доминирование особей с признаком striata. В частности для Центрально-
Черноземного региона, по данным Г.А. Лады [20], доля полосатых особей (по сборам в мае – июне) составляет – 97,2–98,6%%, для р. Северский Донец (Белгородской области, с. Новотаволжанка) – 92,3%, р. Усманка (Воронежский заповедник) – 100%, пойма р. Цна (окр. г. Тамбова) – 97,2%. В Калужской области также преобладают полосатые особи озерной лягушки – от 100% в большинстве исследованных популяций [28]. В частности в Калуге доля полосатых особей составляет от 95,46 до 95,24% в северо-западной, 95% в центральной и восточной части, до 90–84,62% для западной части города [28]. На Нижней Волге доля полосатых особей составляет от 52,1% левого берега Ахтубы (рисовые чеки) до 58,4% окр. оз. Хара (водоемы правобережья старосты Ахту-бы) [21].
Нами проведен анализ накопления тяжелых металлов полосатыми и бесполосыми особями озерной лягушки в районе исследования (табл. 2).
Таблица 2. Накопление тяжелых металлов в коже озерных лягушек (по сухой массе) полосатых и бесполосых морф
|
Популяции, морфа |
Медь |
Цинк |
Свинец |
Кадмий |
||
|
Предуралье |
||||||
|
Теплое |
Кожа (M) 1 бесполосые |
0,18 |
1240,46 |
11,83 |
11,27 |
|
|
Вода (мг/дм3) |
0,061 |
0,078 |
0,004 |
0,018 |
||
|
Затон |
Кожа (M) |
бесполосые |
1032,63 |
1823,70 |
20,49 |
199,07 |
|
Вода (мг/дм3) |
0,076 |
0,123 |
0,0006 |
0,083 |
||
|
Локотки |
Кожа (M) |
бесполосые |
0,25 |
837,62 |
7,43 |
21,48 |
|
Вода (мг/дм3) |
0,041 |
0,063 |
0,0005 |
0,022 |
||
|
Белая |
Кожа (M) |
полосатые |
5,83 |
94,09 |
0,15 |
1,32 |
|
бесполосые |
11,04 |
109,97 |
0,06 |
1,86 |
||
|
Вода (мг/дм3) |
0,018 |
0,039 |
0,011 |
0,001 |
||
|
Зауралье |
||||||
|
Худолаз |
Кожа (M) |
полосатые |
57,97 |
735,87 |
8,89 |
3,77 |
|
бесполосые |
81,34 |
736,32 |
14,31 |
48,42 |
||
|
Вода (мг/дм3) |
0,021 |
0,379 |
0,0005 |
0,003 |
||
|
Таналык |
Кожа (M) |
бесполосые |
7,51 |
862,85 |
5,01 |
14,98 |
|
Вода (мг/дм3) |
0,023 |
0,447 |
0,008 |
0,004 |
||
По данным таблицы 2 видно, у бесполосых происходит наибольшее, за исключение свинца, накопление кадмия, цинка, меди в местообитании со средним фоновым содержанием и высоким антропогенным воздействием (район г. Салават). В условиях высокого природного фона и техногенного загрязнения водоемов горнообогатительными предприятиями для популяции «Худолаз» также у бесполосых особей наблюдается более высокое накопление таких тяжелых металлов, как медь, свинец, кадмий (за исключением цинка). Следует отметить, что в условиях критического загрязнения – выборка из популяции «Таналык», накопление меди и свинца ниже, чем у полосатых особей из популяции «Худолаз».
Наши данные (табл. 2) по озерной лягушке из Предуралья показывают снижение накопления в ряду медь (в 1,9 раз)→кадмий (в 1,4 раза)→цинк (в 1,2 раза), при этом полосатые особи накапливают свинец в среднем в 2,5 раза больше, чем бесполосые. В Зауралье ряд снижения накопления имеет вид: кадмий (в 12,8 раза)→свинец (в 1,6 раз)→медь (в 1,9 раза), цинк накапливается у полосатых и бесполосых особей на одном уровне.
В литературе указывается, что «относительно слабая способность к биоаккумуляции и высокая скорость физиологических процессов у амфибий обусловили увеличение частоты встречаемости морфы striata в пределах естественных и искусственных геохимических аномалий [4] и «антропогенно дестабилизированных территорий» [5, c. 68].
По данным С.А. Шарыгина [35] полосатые особи striata остромордой лягушки накапливают, в отличие от бесполосых, в 5 раз меньше стронция–90, марганца в 3,5 раза, хрома в 5 раз, никеля в 4 раза, олова в 6 раз, цинка в 2,5 раза меньше, за исключением железа, которое полосатые особи накапливают в 2 раза больше. Более низкое накопление данных металлов в организме полосатых особей R . arvalis обуславливает высокую встречаемость полосатых особей в Уральском регионе [35].
Наши данные из таблицы 2 показывают, что только накопление кадмия ниже у особей озерной лягушки с морфой striata в Предуралье («Белая»), в отличие от Зауралья («Худолаз»).
Данные таблицы 2 подтверждают более низкое накопление морфами striata отдельных видов металлов. С другой стороны, в условиях высокого техногенного и природного уровня территории Республики Башкортостан преобладают популяции, где доля полосатых особей незначительна или они отсутствуют (рис. 1). Данный факт не согласуется с выводами А.С. Шарыгина [35] о влиянии геохимической провинции Урала на встречаемость в популяции особей с дорсомеди-альной полосой. По нашим данным, для Зауралья также отмечается доминирование полосатых особей остромордой лягушки по сравнению с Преду-ральем.
Таким образом, наши данные подтверждают выводы В. Л. Вершинина [5] о проявлении адаптационных свойств морфы striata у представителей рода Rana . При этом наблюдаются географические особенности в распределении полосатых морф озерной лягушки, не связанные с уровнем тяжелых металлов в местообитаниях.
В большинстве литературных данных отмечается увеличение доли особей морфы striata в условиях антропогенного воздействия городов Урала [5], Предкавказья [24], Самарской [29–31] и Ульяновской [26] областей. За пределами России, в Болгарии, увеличение доли полосатых особей озерной лягушки отмечается для популяций г. Пловдива [8]. Напротив, в целом или в отдельных популяциях, отмечают почти равное соотношение полосатых и бесполосых особей [37], снижение доли полосатых отмечается для Калужской области [28], Нижнего Поволжья [21, 37, 38] и даже практически полное отсутствие особей с признаком striata в Самарской области [31; наши данные] и Республики Башкортостан [16]. По данным А.А. Шиян [37, с. 20]: «В популяции лягушек с полей фильтрации сахарных заводов отмечено либо равное соотношение особей морф striata и maculata, либо преобладание (в 2,0 раза) особей морфы maculatа». Доля полосатых особей выше на 6,3% в контроле, по сравнению с рисовыми чеками [21].
При этом в условиях Республики Башкортостан – территории с высоким фоновым содержанием в среде меди, цинка и кадмия, в большинстве популяций озерной лягушки преобладают бес-полосые особи (рис. 2). Увеличение доли беспо-лосых особей данного вида характерно для сопредельных с Республикой Башкортостан регионов – в восточных районах Республики Татарстан [10, 12] и в Самарской области (окр. пос. Яблоня, Похвистневского района) (наши данные).
По нашему мнению, в условиях техногенного загрязнения меньшее накопление тяжелых металлов может свидетельствовать о проявление адаптивной реакции. Обитание в условиях высокого техногенного загрязнения водоемов тяжелыми металлами приводит к изменениям морфофизиологических показателей – относительной массы внутренних органов [13]. Следует отметить, что высокие концентрации тяжелых металлов могут приводить как к снижению массы органов в результате высоких энергетических затрат, направленных на детоксикацию, так и к гипертрофии ткани в результате патологических процессов. Отсутствие полосатых особей в большинстве популяций Республики Башкортостан, по нашему мнению, связано с особенностью локального распределения встречаемости морфы «striata», которая редка и в сопредельных регионах.
Таким образом, эколого-физиологические особенности фенотипов, маркируемых признаком «striata», имеют адаптивные преимущества, связанные как с высоким уровнем обмена, так и с более коротким личиночным периодом развития и повышенной проницаемостью кожных покровов в условиях техногенного загрязнения местообитаний.
Авторы благодарят инженера Сиражитдинову Э.М. (Сибай) за помощь при подготовке проб.
Часть материала собрана при поддержке ФЦП «Интеграция» Э-0121 «Изучение разнообразия амфибий Волжского бассейна (генетические и таксономические аспекты, географическая изменчивость, межвидовая гибридизация, биоиндикация антропогенного воздействия». В 2012 году исследования выполнены при поддержке Российского фонда фундаментальных исследований (проект № 12-04-31774).
Список литературы Особенности полиморфизма по признаку striata в популяциях озерной лягушки Rana ridibunda Pallas, 1771 (Anura, Amphibia) Республики Башкортостан
- Боркин Л.Я., Тихенко Н.Д. Некоторые аспекты морфологической изменчивости, полиморфизма окраски, роста, структуры популяции и суточной активности Rana esculenta на северной границе ареала//Экология и систематика амфибий и рептилий. Труды ЗИН АН СССР. Т. 89. Л., 1979. С. 18-54.
- Вершинин В.Л. О распространении озерной лягушки в городе Свердловске//Экология. 1990. № 2. С. 67-71.
- Вершинин В.Л. Экологические особенности популяций амфибий урбанизированных территорий: Автореф. дис. … д-ра биол. наук. Екатеринбург, 1997. 47 с.
- Вершинин В.Л. Морфа Striata и ее роль в путях адаптациогенеза рода Rana в современной биосфере//Доклады академии наук, 2004. Т. 396, № 2. С. 280-282.
- Вершинин В.Л. Морфа Striata у представителей рода Rana (Amphibia, Anura) -причины адаптивности к изменениям среды//Журнал общей биологии. 2008. Т. 69. № 1. С. 65-71.
- Вершинин В.Л., Терешин С.Ю. Физиологические показатели амфибий в экосистемах урбанизированных территорий//Экология. 1999. № 3. С. 283-287.
- Добринский Л.Н., Малафеев Ю.М. Методика изучения интенсивности выделения углекислого газа мелкими пойкилотермными животными с помощью оптико-акустического газоанализатора//Экология. 1974. № 1. С. 73-78.
- Желев Ж.М. Биоиндикационная оценка состояния двух биотопов в Южной Болгарии на основании флуктуирующей асимметрии и фенетического состава популяций озерной лягушки Rana ridibunda Pallas, 1771 (Anura, Amphibia, Ranidae) и краснобрюхой жерлянки Bombina bombina Linnaeus, 1761 (Amphibia, Anura, Discoglossidae) в условиях синтопического обитания//Перспективы науки. 2011. № 22. С. 7-18.
- Замалетдинов Р.И. Фенотипическая структура популяций зеленых лягушек на урбанизированных территориях//Поволжский экологический журнал. 2002. № 2. С. 163-165.
- Замалетдинов Р.И. Экология земноводных в условиях большого города (на примере г. Казани): Автореф. дис. … канд. биол. наук. Казань, 2003. 24 с.
- Замалетдинов Р.И. Изменчивость цветового полиморфизма озерной лягушки в Республике Татарстан//Сб. науч. тр. «Актуальные проблемы герпетологии и токсинологии». Вып. 8. Тольятти, 2005. С. 38-45.
- Замалетдинов Р.И. Географическая изменчивость цветового полиморфизма зеленых лягушек в Республике Татарстан//Актуальные проблемы герпетологии и токсинологии: Сб. науч. тр. Вып. 10. Тольятти, 2007. С. 61-65.
- Зарипова Ф.Ф., Файзулин А.И. Характеристика морфофизиологических показателей популяций озерной лягушки Rana ridibunda (Anura, Amphibia) урбанизированных территорий Республики Башкортостан//Известия Самарского научного центра Российской академии наук Т. 15, № 5. 2012. С. 145-149.
- Зарипова Ф.Ф., Файзулин А.И. Бесхвостые земноводные (Anura, Amphibia) урбанизированных территорий Республики Башкортостан: видовой состав, распространение, оценка встречаемости и состояние популяций//Экологический сборник 2. Труды молодых ученых Поволжья/под ред. проф. С.В. Саксонова. Тольятти: ИЭВБ РАН, 2009. С. 55-58.
- Зарипова Ф.Ф., Файзулин А.И., Юмагулова Г.Р. Содержание тяжелых металлов в печени озерной лягушки Rana ridibunda Pallas, 1771 Башкирского Зауралья//Вестник Оренбургского государственного университета. № 6. Оренбург, 2009. С. 145-146.
- Зарипова Ф.Ф., Юмагулова Г.Р., Файзулин А.И. Характеристика состояния популяции озерной лягушки Rana ridibunda Pallas, 1771 (Anura, Amphibia) в Республике Башкортостан по полиморфизму рисунка окраски спины//Известия Самарского научного центра Российской академии наук Т. 1, № 1. 2009. С. 78-82.
- Ищенко В.Г. Динамический полиморфизм бурых лягушек фауны СССР. М., «Наука», 1978. 148 с.
- Колякин Н.Н. Озерная лягушка в условиях промышленного города/Н.Н. Колякин//Экологическая и морфологическая изменчивость животных под влиянием антропических факторов. Волгоград, 1994. С. 83-92.
- Кубанцев Б.С., Жукова Т.И. Антропогенные воздействия на среду обитания земноводных и половая структура их популяций//Экологическая и морфологическая изменчивость животных под влиянием антропических факторов. Волгоград, 1994. С. 124.
- Лада Г.А. О генетическом полиморфизме озерной лягушки в Центральном Черноземье//Материалы 4 всесоюз. совещ. «Фенетика популяций». М., 1990. С. 151-152.
- Лебединский А.А. Гельминтофауна озерных лягушек в условиях антропогенного воздействия и связь инвазии с полиморфизмом//Животные в природных экосистемах. Н. Новгород, 1994. С. 25-32.
- Леденцов А.В. Динамика возрастной структуры и численности репродуктивной части популяции остромордой лягушки (Rana arvalis Nilss.): Автореф. дисс.. канд. биол. наук. Свердловск, 1990. 18 с.
- Никашин И.А. Эколого-морфологические признаки популяций озерной лягушки (Rana ridibunda Pall.) как средство оценки антропогенного воздействия на водные экосистемы (на примере Липецкой области): Автореф. дис. … канд. биол. наук. Липецк, 2007. 17 с.
- Пескова Т.Ю. Адаптационная изменчивость земноводных в антропогенно загрязненной среде: Автореф. дисс. … д-ра биол. наук. Тольятти, 2004. 36 с.
- Рункова Г.Г. Опыт применения некоторых методов математического планирования эксперимента в эколого-биологических исследованиях//Математическое планирование эксперимента в биологических исследованиях. Свердловск, 1975. Вып. 97. С. 18-104.
- Спирина Е.В. Амфибии как биоиндикационная тест-система для экологической оценки водной среды обитания: Автореф. дисс.. канд. биол. наук. Ульяновск, 2007. 23 с.
- Терентьев П.В. Характер географической изменчивости зеленых лягушек//Труды Петергофского биологического института ЛГУ. 1962. № 19. С. 98-121.
- Устюжанина О.А., Стрельцов А.Б. Изменчивость и встречаемость морфы striana у Rana ridibunda, R. lessonae, R. esculenta в Калужской области//Зоол. ж. 2005. Т. 84, № 6. С. 699-706.
- Файзулин А.И. Эколого-фаунистический анализ земноводных Среднего Поволжья и проблемы их охраны: Автореф. дисс.. канд. биол. наук. Тольятти, 2004. 20 с.
- Файзулин А.И. Антропогенные воздействия -микроэволюционный фактор?//В мире научных открытий, 2010, №4 (10), Часть 1. С. 77-80.
- Файзулин А.И., Кузовенко А.Е. Использование амфибий в мониторинге состояния окружающей среды в условиях Самарской области: фенетическая структура популяций//Известия Самарского научного центра Российской академии наук Т. 1 (3), № 1. 2012. С. 829-833.
- Файзулин А.И., Чихляев И.В. Возрастная динамика полиморфизма озерной лягушки (Rana ridibunda) из района Мордовинской поймы (Национальный парк «Самарская Лука»)//Актуальные проблемы герпетологии и токсинологии: Сб. науч. тр. Вып. 4. Тольятти, 2000. С. 75-78.
- Файзулин А.И., Чихляев И.В. Характеристика полиморфизма озерной лягушки (Rana ridibunda) Самарской Луки//Бюлл. «Самарская Лука». № 11-01. Самара, 2001. С. 314-318.
- Чибилёв Е.А. Биология и экология зеленых и бурых лягушек Челябинской городской агломерации//Материалы 1 Международной научно-практической конференции «Животные в антропогенном ландшафте». Астрахань: Изд-во Астраханского гос. ун-та, 2003. С. 73-76.
- Шарыгин С.А. Микроэлементы в организме некоторых амфибий и рептилий и их динамика под влиянием антропогенных факторов: Автореф. дис. … канд. биол. наук. Свердловск, 1980. 24 с.
- Шварц С.С., Ищенко В.Г. Динамика генетического состава популяций остромордой лягушки//Бюлл. Моск. об-ва испыт. природы. Отд. биол. 1968. Т. 73. № 4. С. 127-134.
- Шиян А.А. Экологическая характеристика озерной лягушки (Rana ridibunda Pall.) при обитании на полях фильтрации сахарных заводов: Автореф. дис. … канд. биол. наук. Саратов, 2011. 21 с.
- Шиян А.А. Изменения популяционных характеристик озерной лягушки (Rana ridibunda Pall.) при обитании в прудах-испарителях сахарных заводов//Научный журнал Кубанского государственного аграрного университета [Электронный ресурс]. Краснодар: КубГАУ, 2011. № 67 (3). URL: http://ej.kubagro.ru/2011/03/pdf/27.pdf (дата обращения: 20.08.2012).
- Щупак Е.Л. Наследование спинной полосы особями остромордой лягушки//Информационные материалы института экологии растений и животных. Свердловск: ИЭРиЖ УрО АН СССР, 1977. С. 36.
- Berger L., Smielowski J. Inheritance of vertebral stripe in Rana ridibunda Pall. (Amphibia, Ranidae)//Amphibia-Reptilia. 1982. Vol. 3. P. 145-151.
- Borkin L.Ja., Garanin W.I., Tichenko N.T., Zaune I.A. Some results in the green frogs survay in the USSR//Mitt. zool. Mus. Berlin. 1979. Pl. 1. № 55. P. 153-170.
- Faizulin A.I. Adaptive strategies and assessment of tolerance to anthropogenic impacts on the territory of the Volga River Basin//Types of Strategy and not only (Materials of the Fourth Russian-Polish School of Young Ecologists. Togliatti: Kassandra, 2010. Р. 11-14.
- Hoffman E.A., Blouin M.S. A revier of color and pattern polymorphisms in anurans//Biological Journal of Linnean Society 2000. 70. № 4. P. 633-665.
