Особенности поведения полевок (Microtus, Rodentia)
")
Автор: Лучникова Екатерина Михайловна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Статья в выпуске: 4, 2014 года.
Бесплатный доступ
На основании проведенного исследования биоритмов и экспериментального поведения были выявлены этолого-экологические особенности полевок, обитающих на территории Западной Сибири. Выявленные различия могут быть расценены как один их механизмов дифференциации экологических ниш и как способ снижения межвидовой конкуренции у совместно обитающих видов.
Грызуны, полевки, суточная активность, экспериментальное поведение, межвидовая конкуренция
Короткий адрес: https://sciup.org/146116531
IDR: 146116531 | УДК: 599.363
Peculiarities of behavior in the voles (Microtus, Rodentia)
The study of biorhythms and experimental behavior revealed the ethological and ecological peculiarities of the voles, inhabiting the Western Siberia. Discovered peculiarities can be treated as the mechanisms in differentiation of the ecological niches, as well as a factor, lowering the interspecific competition in closely living species.
Текст научной статьи Особенности поведения полевок (Microtus, Rodentia)
Введение. В настоящее время активно ведутся исследования популяций совместно существующих видов, относящихся к одной таксономической группе. Изучение поведения близкородственных видов - потенциальных конкурентов направлено, в первую очередь, на выявление этологических и биоритмологических путей дифференциации их экологических ниш (Шенброт, 1986).
Удобным объектом для исследования эколого-этологических аспектов межвидовых отношений являются серые полевки (р. Microtus) В условиях Западной Сибири они сосуществуют между собой и с лесными полевками (р. Myodes^ на достаточно большой территории. Изучение этологических особенностей серых полевок помогает составить наиболее полную картину адаптаций этой группы к условиям обитания и сосуществования с лесными полевками.
Методика. Исследования проводились на биостанции КемГУ «Ажендарово» (Кемеровская обл.) в июле-августе 2010-2013 гг. Для сравнительного анализа использованы данные по поведению и биоритмологии лесных полевок, полученные автором, а также материалы кафедры зоологии и экологии КемГУ, в том числе биоритмологические наблюдения, проведенные в Томской обл.
Полевки, участвующие в экспериментах, были отловлены при помощи стандартных ловчих канавок и живоловок (Онищенко и др., 2010) и содержались в индивидуальных вольерах в режиме естественного освещения, влажности и температуры. Для ночных наблюдений использовали красный монохроматический свет. Биоритмологические данные были получены с использованием методики изучения суточной активности, предложенной Л.Н. Ердаковым (1972). В целях изучения поведения полевок в острой стрессовой ситуации проводились тесты «Открытое поле» и «Экстраполяционное избавление». Процессы ориентации и запоминания были исследованы при помощи 8-рукавного радиального лабиринта и водного теста Мориса. Объем наблюдений представлен в таблице 1.
Таблица 1
Видовой состав полевок, участвовавших в экспериментах
|
Виды |
Суточная активность |
Тест «Открытое поле» |
Тест «Экстраполяционн ое избавление» |
Водный тест Морриса |
|
Полевка-экономка Microtus oeconomus |
12 |
22 |
18 |
15 |
|
Пашенная полевка М. agrestis |
3 |
14 |
11 |
16 |
|
Обыкновенная полевка М. arvalis |
4 |
7 |
8 |
7 |
|
Рыжая полевка Myodes glareolus |
11 |
29 |
29 |
28 |
|
Красная полевка М. rutilus |
5 |
12 |
11 |
11 |
|
Красно-серая полевка М. г и foe anus |
7 |
8 |
7 |
7 |
Полученные данные были обработаны при помощи стандартных статистических методов с использованием пакета прикладных программ «Морфолог». Достоверность различий оценивалась при помощи Т-критерия Стьюдента. Для анализа биоримологических данных были построены хронограммы.
Результаты и обсуждение. Разные виды полевок р. Microtus различаются по биоритмологическим показателям. Все исследованные виды характеризуются 24-часовым полифазным ритмом суточной активности, однако показатели общей и двигательной активности, их пики и спады у разных видов не совпадают.
У полевки-экономки в суточном ритме преобладает ночная и утренняя активность с длительным пиком между 24 и б часами. В дневные часы активность падает, достигая минимума в 15 и 19 часов. Общая и двигательная активность колеблются синхронно, а в вечерние часы вклад двигательной компоненты достигает 100%. При сопоставлении хронограмм суточной активности полевок-экономок Кемеровской и Томской обл. можно отметить, в целом, совпадение их ритмов суточной активности, что свидетельствует о достаточной консервативности и позволяет экстраполировать полученную информацию для Западной Сибири.
У пашенной полевки преобладает ночная и сумеречная активность с пиком между 23 и 24 часами. Спад активности - 134- наблюдается после восхода Солнца и вплоть до сумерек. Полифазность ритма выражена в наибольшей степени среди всех исследованных нами видов - в каждом часе наблюдения мы регистрировали как сон, так и бодрствование, в том числе питание, передвижение, благоустройство.
У обыкновенной полевки максимумы активного состояния приурочены к ночным и ранним утренним часам с пиками между 3 и б часами. В 23 и 7 часов наблюдаются наиболее выраженные спады активности.
При сопоставлении хронограмм трех видов серых полевок можно отметить, что ритмы активности пашенной полевки и полевки-экономки практически совпадают, в то же время, суточный ритм обыкновенной полевки значительно отличается от ритмов вышеуказанных видов; на протяжении длительного промежутка наблюдается полная инверсия римтов, которая может быть расценена как разделение экологической ниши по временной оси.
Анализ экспериментальных данных позволяет утверждать, что поведение полевок различается в зависимости от их родовой и видовой принадлежности. В незнакомой обстановке, в том числе, при острой стрессовой ситуации, серые полевки в большей степени сохраняют способность к решению задач, чем рыжие полевки. Так, в тесте «Экстраполяционное избавление» 88% серых полевок нашли выход из экспериментальной установки. В эксперименте они сначала пытались найти выход неверным способом - выпрыгнуть; тем не менее, именно представители этого вида продемонстрировали лучшие скоростные показатели в решении задачи. Лесные полевки сразу же выбирали безпрыжковую стратегию выхода, но оказались наиболее медлительными, затрачивая на избавление вдвое больше времени (р<0,05).
В водном тесте Морриса серые полевки запоминали расположение подводной платформы и целенаправленно искали ее, о чем свидетельствует резкое уменьшения времени, затраченного на решение задачи уже при второй попытке. Запоминание положения невидимой платформы происходило уже в первой фазе опыта, а на следующих стадиях полевки активно использовали свои экстраполяционные способности, о чем свидетельствовала правильная траектория движения.
В тесте «Открытое поле» все исследованные виды полевок (как рода Microtus, так и род Myodes) стремятся к передвижению по периферическим (пристеночным) секторам, что отражает скрытный образ жизни этих грызунов и их стремление найти убежище. У представителей рода серых полевок отмечена повышенная вертикальная активность, зверьки пытаются покинуть экспериментальную установку, совершая прыжки и вертикальные стойки. Представители рода серых полевок, в целом, более активны, нежели лесные полевки. Поведение
Вестник ТвГУ. Серия "Биология и экология". 2014. № 4 полевки-экономки и темной (пашенной) полевки не показало достоверных различий (р>0,05).
В тесте «8-рукавный лабиринт» все серые полевки проявляют способности к пространственному обучению и формированию «образа» лабиринта, однако скорость этих процессов у разных видов различается. У полевки экономки практически сразу формируется вся картина лабиринта, они склонны к последовательному обследованию рукавов лабиринта и уже после первого прохождения делают в среднем по одной ошибке, в связи этим и время, затраченное на попытки у них невелико.
Пашенная и обыкновенная полевки обследуют рукава лабиринта без четкой последовательности и ошибаются чаще экономки (р<0,05), но в конечном итоге также формируют устойчивую картину лабиринта. На ранних стадиях пашенная полевка склонна подолгу задерживаться (затаиваться) в рукавах лабиринта, что существенно увеличивает время его прохождения. Тем не менее, на последних попытках у всех трех видов результаты практически совпадают. Таким образом, все исследуемые виды демонстрируют способности к обучению и когнитивным процессам - формированию образа лабиринта.
Самые интересные данные были получены при сравнительном анализе поведения в лабиринте наиболее многочисленных представителей родов серых и лесных полевок (полевка-экономка и рыжая полевка). Они были подвергнуты анализу с использованием метода главных компонент (рис. 1, табл. 2).
Таблица 2 Наиболее значимые вклады признаков в распределение выборок полевок в пространстве первых двух главных компонент (рис.1)
|
Компонента |
1 |
2 |
|
лямбда |
3,588 |
2,748 |
|
% дисперсии |
30,631 |
23,467 |
|
Вклад признака |
вес |
вес |
|
1 попытка |
+0,028 |
+0,328 |
|
2 попытка |
+0,067 |
+0,472 |
|
3 попытка |
-0,064 |
+0,315 |
|
4 попытка |
+0,022 |
+0,522 |
|
8 попытка |
+0,290 |
-0,038 |
|
9 попытка |
+0,654 |
-0,086 |
|
10 попытка |
+0,667 |
-0,039 |
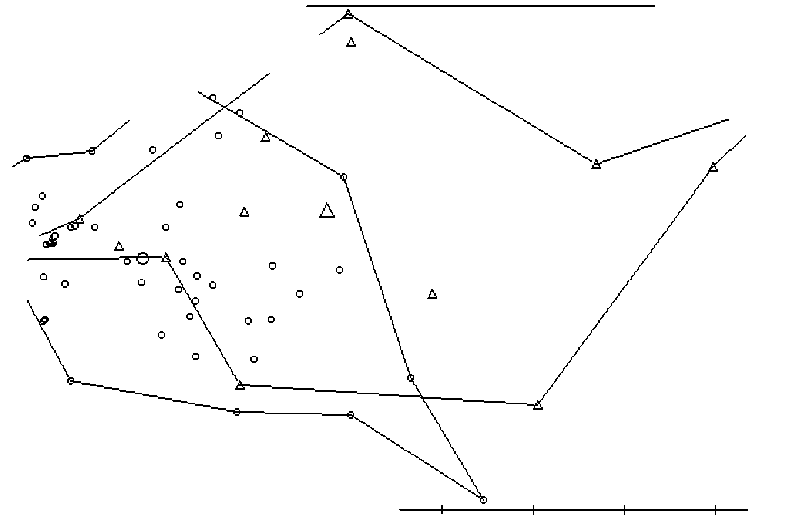
Рис. 1 . Скаттер-диаграмма распределения выборок полевок в пространстве первых двух главных компонент (вклады признаков см. в табл. 3), построенная на основе количества ошибок.
Кружки - полевка-экономка, треугольники - рыжая полевка.
Как видно на скаттер-диаграмме, выборки изучаемых видов различаются по первой компоненте (хотя и с большим перекрыванием). Анализ вкладов исходных признаков в эту компоненту позволяет её интерпретировать как описывающую различия по количеству ошибок в последних трёх попытках прохождения лабиринта. Действительно, полёвки-экономки совершали меньше ошибок, чем рыжие (табл. 3).
Различия по второй компоненте менее значимы и в ней наиболее весомыми оказываются ошибки, связанные с первыми четырьмя прохождениями. С позиций этологии это можно объяснить сменой мотивацией при прохождении лабиринта (исследовательская/пищевая).
Выявлены достоверные различия по количеству ошибок, совершенных полёвками-экономками и рыжими полёвками в 1 -4, б и 10 попытке (табл. 4).
Таким образом, оказавшись в лабиринте, полёвка-экономка начинает искать выход из лабиринта и практически не реагирует на приманку; в середине опыта грызун уже знает, что выхода из лабиринта нет, и начинает искать пищу; к последним попыткам полёвка-экономка уже хорошо ориентируется в лабиринте и во время его прохождения допускает минимальное количество ошибок. Полёвки-экономки живут, как правило, на сырых лугах, полянах, травяных болотах, также часто встречаются в редколесьях около водоёмов, в поймах рек, реже обитают в лесах. Чтобы выжить в таких непостоянных условиях среды, грызуну необходимо хорошо ориентироваться на местности. Рыжая же полёвка обитает преимущественно в смешанных лесах и на лугах с высокотравной растительностью, отличающихся более стабильными условиями; от нее не требуется столь чётко ориентироваться на местности. Это наблюдается во время опытов. Рыжая полёвка спокойно обследует рукава лабиринта и ищет пищу. Посему, поведение грызунов в лабиринте можно объяснить различием мотиваций: полёвка-экономка ищет выход из лабиринта, а рыжая полёвка - пищу.
Таблица 3
Статистические показатели этологических параметров (количество ошибок)
|
№ попытки |
М±т |
lim (min - max) |
CV, % |
|
|
Полевки -экономки |
||||
|
1 попытка |
1,52 ±0,162 |
0,00 - 6,00 |
1,193 |
78,59 |
|
2 попытка |
1,11 ±0,137 |
0,00 - 4,00 |
1,003 |
90,28 |
|
3 попытка |
1,20 ±0,133 |
0,00 - 4,00 |
0,979 |
81,30 |
|
4 попытка |
1,11 ±0,131 |
0,00 - 5,00 |
0,965 |
86,83 |
|
5 попытка |
1,09 ±0,168 |
0,00 - 7,00 |
1,233 |
112,82 |
|
6 попытка |
1,00 ±0,112 |
0,00 - 4,00 |
0,824 |
82,42 |
|
7 попытка |
0,94 ±0,131 |
0,00 - 5,00 |
0,960 |
101,64 |
|
8 попытка |
0,91 ±0,113 |
0,00 - 3,00 |
0,830 |
91,50 |
|
9 попытка |
0,55 ±0,126 |
0,00 - 4,00 |
0,925 |
166,47 |
|
10 попытка |
0,44 ± 0,098 |
0,00 - 2,00 |
0,718 |
161,58 |
|
Рыжие полевки |
||||
|
1 попытка |
2,43 ±0,500 |
0,00 - 7,00 |
1,869 |
76,97 |
|
2 попытка |
2,42 ± 0,562 |
0,00 - 7,00 |
2,102 |
86,54 |
|
3 попытка |
2,28 ±0,518 |
0,00 - 7,00 |
1,939 |
84,81 |
|
4 попытка |
2,21 ±0,547 |
0,00 - 7,00 |
2,045 |
92,35 |
|
5 попытка |
1,57 ±0,453 |
0,00 - 5,00 |
1,697 |
107,98 |
|
6 попытка |
2,00 ± 0,457 |
0,00 - 7,00 |
1,710 |
85,49 |
|
7 попытка |
1,50 ±0,344 |
0,00 - 4,00 |
1,286 |
85,73 |
|
8 попытка |
1,28 ±0,266 |
0,00 - 3,00 |
0,994 |
77,35 |
|
9 попытка |
0,92 ± 0,245 |
0,00 - 3,00 |
0,917 |
98,74 |
|
10 попытка |
0,93 ± 0,267 |
0,00 - 3,00 |
0,997 |
107,40 |
Таблица 4
Достоверные статистические различия в поведении рыжей полёвки и полёвки-экономки во время прохождения 8-рукавного лабиринта (значения Т-критерия Стьюдента ниже 95 % не приведены)
|
№ попытки |
T - критерий Стьюдента |
Уровень различий |
|
1 |
2.242 |
> 95% |
|
2 |
3.391 |
> 99% |
|
3 |
2.937 |
> 99% |
|
4 |
2.935 |
> 99% |
|
6 |
3.149 |
> 99% |
|
10 |
2.067 |
> 95% |
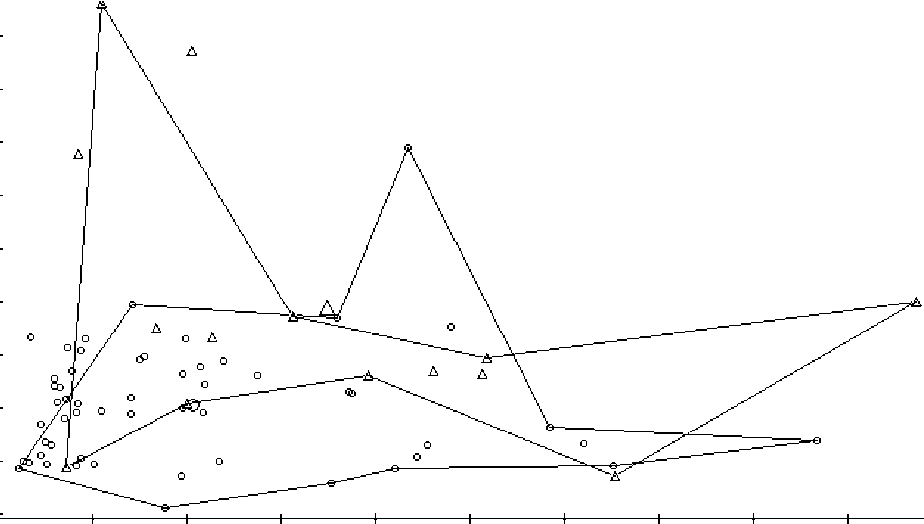
Рис. 2. Скаттер-диаграмма распределения выборок полевок в пространстве первых двух главных компонент (вклады признаков см. в табл.5) построенная на основе времени прохождения лабиринта.
Кружки - полевка-экономка, треугольники - рыжая полевка
Таблица 5
Вклады признаков в распределение выборок полевок в пространстве первых двух главных компонент (рис. 2)
|
компонента |
1 |
2 |
|
лямбда |
1,881 |
0,616 |
|
% дис, |
44,179 |
14,475 |
|
вес |
вес |
|
|
1 попытка |
+0,233 |
-0,084 |
|
2 попытка |
+0,341 |
-0,080 |
|
3 попытка |
+0,313 |
+0,591 |
|
4 попытка |
+0,304 |
+0,308 |
|
5 попытка |
+0,386 |
+0,165 |
|
6 попытка |
+0,286 |
+0,259 |
|
7 попытка |
+0,294 |
+0,009 |
|
8 попытка |
+0,280 |
-0,409 |
|
9 попытка |
+0,301 |
-0,383 |
|
10 попытка |
+0,391 |
-0,365 |
Скаттер-диаграмма, иллюстрирующая распределение выборок полёвок по времени прохождения лабиринта, представлена на рис. 2. Очевидно, что выборки изучаемых видов различаются по первой компоненте с небольшим перекрыванием. Анализ вкладов исходных признаков в эту компоненту позволяет её интерпретировать как описывающую различия по времени прохождения лабиринта, причём примерно по всем попыткам в равной степени - экономки в целом быстрее, чем рыжие справлялись с заданием на протяжении всего цикла опыта (табл. 5, 6).
Выявлены достоверные различия (табл. 7) между полёвками-экономками и рыжими полёвками по времени прохождения лабиринта (2-6 и 10 попытки).
Таблица 6 Статистические показатели этологических параметров
(время прохождения лабиринта) у полевок-экономок
|
№ попытки |
М±т |
lim (min - max) |
CV, % |
|
|
Полевки-экономки |
||||
|
1 попытка |
3,24 ±0,288 |
0,00 - 10,00 |
2,093 |
64,50 |
|
2 попытка |
2,22 ±0,195 |
0,00 - 6,00 |
1,423 |
63,91 |
|
3 попытка |
2,24 ±0,183 |
0,00 - 7,00 |
1,329 |
59,18 |
|
4 попытка |
2,01 ±0,130 |
0,00 - 4,00 |
0,943 |
46,93 |
|
5 попытка |
1,98 ±0,165 |
0,00 - 6,00 |
1,201 |
60,61 |
|
6 попытка |
1,97 ±0,164 |
0,00 - 5,00 |
1,195 |
60,59 |
|
7 попытка |
2,28 ±0,169 |
1,00-6,00 |
1,231 |
53,90 |
|
8 попытка |
1,93 ±0,159 |
0,50 - 7,00 |
1,160 |
60,00 |
|
9 попытка |
1,93 ±0,185 |
0,00 - 6,00 |
1,345 |
69,53 |
|
10 попытка |
1,55 ±0,150 |
0,00 - 5,00 |
1,090 |
70,05 |
|
Рыжие полевки |
||||
|
1 попытка |
4,20 ±0,587 |
1,00-7,00 |
2,274 |
54,14 |
|
2 попытка |
3,46 ± 0,524 |
1,00-7,00 |
2,031 |
58,58 |
|
3 попытка |
4,10 ±0,707 |
0,50 - 9,00 |
2,740 |
66,83 |
|
4 попытка |
3,16 ±0,620 |
0,50 - 8,00 |
2,403 |
75,88 |
|
5 попытка |
3,93 ±0,565 |
1,00-7,00 |
2,187 |
55,59 |
|
6 попытка |
3,00 ±0,390 |
1,00-7,00 |
1,512 |
50,40 |
|
7 попытка |
3,00 ±0,488 |
1,00-7,00 |
1,890 |
62,99 |
|
8 попытка |
2,33 ± 0,444 |
1,00-6,00 |
1,718 |
73,64 |
|
9 попытка |
2,53 ± 0,307 |
1,00-5,00 |
1,187 |
46,86 |
|
10 попытка |
2,33 ± 0,398 |
1,00-5,00 |
1,543 |
66,13 |
Таблица 7
Достоверные статистические различия в поведении рыжей полёвки и полёвки-экономки во время прохождения 8-рукавного лабиринта (значения Т-критерия Стьюдента ниже 95 % не приведены)
|
№ попытки |
Т - критерий Стьюдента |
Уровень различий |
|
2 |
2.698 |
> 99 % |
|
3 |
3.671 |
> 99.9% |
|
4 |
2.852 |
> 99% |
|
5 |
4.552 |
> 99.9% |
|
6 |
2.772 |
> 99% |
|
10 |
2.212 |
> 95% |
Заключение. Все исследованные виды серых полевок относятся к полифазным видам, с преобладанием активности в темное время суток. Суточный ритм обыкновенной полевки значительно отличается от ритмов пашенной плевки и полевки-экономки. Выявленные биоритмологические различия могут быть расценены как один их механизмов дифференциации экологических ниш и как способ снижения межвидовой конкуренции у совместно обитающих видов. Все полевки демонстрируют способности к обучению и когнитивным процессам - формированию образа лабиринта, однако успешнее и быстрее всего эти процессы протекают у полевок-экономок. В острой стрессовой ситуации они демонстрируют активную поведенческую стратегию и эффективнее других серых полевок справляются с экспериментальной задачей. Представители рода Microtus показывали наилучшие результаты обучения в радиальном 8-рукавном лабиринте. Они быстрее других справлялись с поставленной задачей и допускали минимальное количество ошибок. Такое поведение может обусловлено образом жизни серых полёвок. Эти виды обитают преимущественно в пойме, где условия среды постоянно меняются, вынуждая животных быстро приспосабливаться к любым изменениям обстановки и быстрее других видов реагировать на стрессовые ситуации (в частности, паводки).
Лучникова Е.М. Особенности поведения полевок (Microtus, Rodentia) /
Е.М. Лучникова // Вести. ТвГУ. Сер. Биология и экология. 2014. № 4. С. 133-142.
Список литературы Особенности поведения полевок (Microtus, Rodentia)
- Ердаков Л.Н. 1978. Простая методика регистрации суточной активности//Известия СО АН СССР. С. 135-138.
- Онищенко С.С., Ильяшенко В.Б., Лучникова Е.М., Бибик Е.В., Скалон Н.В. 2010. Практические рекомендации по изучению мелких млекопитающих. Кемерово: КГУ. 96 с.
- Шенброт Г.И. 1986. Экологические ниши, межвидовая конкуренция и структура сообществ наземных позвоночных//Экологические, этологические и эволюционные аспекты организации многовидовых сообществ позвоночных: Итоги науки и техники. Серия «Зоология позвоночных». М.: ВИНИТИ. Т. 14. С.5-70.
