Особенности распространения, биологии, экологии и морфологии ужа обыкновенного Natrix natrix (Linneaus, 1758) Волгоградской области
 Волгоградской области")
Автор: Гордеев Д.А.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 1-1 т.14, 2012 года.
Бесплатный доступ
В настоящей работе обсуждаются региональные особенности биологии, экологии и морфологии ужа обыкновенного, показано распространение вида на территории Волгоградской области, дана характеристика подвидов на основе метрических и меристических признаков. Проанализированы основные показатели, применяемые в систематике вида.
Уж обыкновенный, пространственное распределение, сезонная и суточная динамика, популяционные особенности, морфология вида
Короткий адрес: https://sciup.org/148200541
IDR: 148200541 | УДК: 598.126.3
Features of biology, ecology and morphology snake ordinary Natrix natrix (Lineaus, 1758), Volgograd region
This paper discusses the regional differences of biology, ecology and morphology of the ordinary snake, species distribution is shown in the Volgograd region, the characteristic of subspecies on the basis of metric and meristic traits. Analyzes the main indicators used in the taxonomy of the species.
Текст научной статьи Особенности распространения, биологии, экологии и морфологии ужа обыкновенного Natrix natrix (Linneaus, 1758) Волгоградской области
Обыкновенный уж ( Natrix natrix Linnaeus, 1758) распространен в Европе (кроме Ирландии), северной части Великобритании, северной части Скандинавии, северо-западной Африке и западной Азии до северно-западной Монголии, юга Восточной Сибири и прилежащих районов севера Китая на востоке и юго-западного Ирана на юге[1-3]. Из девяти подвидов в России обитает три: [1, 2, 4] N. n. natrix (Linnaeus, 1758 ) – на большей территории европейской части СНГ, исключая Заволжье, крайних юговосточных районов и восточного Предкавказья; N. n.scutata (Pallas, 1771) – в Заволжье, на Урале, в Западной Сибири, Казахстане, Бурятии и на юге Восточной Сибири; N. n. persa (Pallas, 1814) известен из восточного Предкавказья, Закавказья и югозападного Туркменистана, из Крыма по единичным находкам. Таким образом, обыкновенный уж – наиболее широко распространенный и многочисленный вид семейства Colubridae , обитающий в европейской части России, однако сведения о его экологии в Волгоградской области до настоящего времени неполны, что определило актуальность данного исследования.
Нами учитывались стандартные метрические и меристические показатели, а также количество че-шуй вокруг глаза, число предглазничных и заглаз-ничных щитков. Плотность микропопуляций определялась маршрутным методом и учетом на пробных площадках [5]. Статистическая обработка полученных данных производилась по общепринятым методикам и включала расчет средних значений для каждого показателя, ошибку, а также коэффициент вариации признаков и расчет достоверности различий (коэффициент Стьюдента).
В условиях Волгоградской области, как и на остальной территории ареала [6-8], обитание ужа обыкновенного приурочено к увлажненным участкам преимущественно интразонального характера: обычны в зарослях кустарника по берегам водоемов и рек, поймах, при этом избегает открытых пространств (для Волгограда известна микропопуляция,
часть ареала которой занимает степной биотоп, однако распространение вида от водоема не превышает 100 м).
Установлено, что в условиях Волгоградской области встречается два подвида: N. n. natrix (Linnaeus, 1758) и N. n.scutata (Pallas, 1771) , отмеченных повсеместно (рис.).
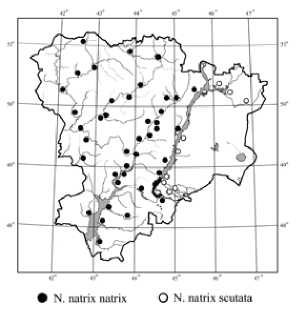
Рис. Обитание ужа обыкновенного на территории Волгоградской области
Ареал N. n. natrix (Linnaeus, 1758) занимает Правобережье Волги вдоль крупных рек: Волги, Дона и их притоков: Арчеды, Медведицы, Иловли, Хопра, озер и прудов. Плотность микропопуляций подвида варьирует от 2,0-4,1 ос./га (р. Иловля) до 10 ос./га (р. Дон). Для Волгограда плотность популяции номинативного подвида изменяется от 1,3 до 3,3 ос./га. В среднем на 1 км маршрута приходится 20 особей. Ареал N. n.scutata (Pallas, 1771) занимает Левобережье Волги. Среди исследованных биотопов, наибольшая плотность микропопуляции отмечена для Волго-Ахтубинской пойме, где может достигать 12 ос/га (ерик Каширский). Причем, в зависимости от характера местности, данный показатель варьирует от 3 до 7 ос./га (ерик Судомойки), а на испарителе Большой Лиман (к востоку от г. Волжский) достигает значения 5,2 ос./га.
Выход с мест зимовки начинается в марте (20-е числа), однако, массовое появление приходится на вторую половину апреля. Брачный период начинается вскоре после выхода с зимовки, как правило, в начале мая. В это время отмечается максимум активности змей и плотности популяций. Откладка яиц приходится на конец июня, в кладке их насчитывается от 10 до 25. Сеголетки появляются в конце августа и имеют следующие размерные характеристики: длина тела 150,2-189,3 мм, хвоста – 35,8-53,1 мм. Максимальная длина тела животного в условиях области была зафиксирована в г. Волгограде и составила 840,0 мм, а хвоста – 170,2 мм.
Весной змеи активны в течение всего дня с 9:00 до 19:00 ч, с увеличением дневной температуры в летний период первый пик длится с 7:00 до 12:00 ч, второй с 16:00 до 20:00 ч, осенью подвижны в течение всего дня. Таким образом, суточная активность носит сезонный одновершинный и двухвершинный характер.
Убежищем служат заросли тростника, корни деревьев, расщелины субстрата, обычны под камнями, пнями и корягами.
По результатам наших наблюдений (которые согласуются с данными других исследователей) в пищевом рационе ужа преобладают бесхвостые амфибии, значительно реже мальки рыб и насекомые. То есть, в пищевом отношении обыкновенный уж – хорошо выраженный батрахофаг.
Змеи хорошо плавают и ныряют. Они могут находиться под водой довольно продолжительное время: охотясь или спасаясь от врагов. Пойманное животное некоторое время шипит, активно извивается, пытаясь скрыться, если это не помогает – выделяет из клоаки жидкость, обладающую резким неприятным запахом. Если и это не помогает, то долгое время остается неподвижным, ложась на спинную сторону и широко раскрывая рот высовывая при этом язык, притворяясь мертвой.
За сезон рептилии совершают две-три миграции: весенняя связана с выходом из зимних убежищ, поиском партнера и мест для спаривания, распределению по прилегающим биотопам; летняя - поиск самкой мест для откладки яиц, перемещение в места с наибольшей плотностью жертв; осенняя – поиск мест зимовки. Первый этап миграции начинается вскоре после выхода с зимовки, характеризуется незначительными расстояниями в виду низкой активности рептилий, летний более продолжительный и связан с засухой или сменой питания, третий этап самый продолжительный по времени и расстоянию перемещения особей. Могут мигрировать на небольшие расстояния в поисках корма. На зимовку уходят в конце сентября – начале октября, период активности длится около 210 дней.
Врагами являются серая цапля (Ardea cinerea), большая белая цапля (Ardea alba) , обыкновенная лисица (Vulpes vulpes) , медянка обыкновенная (Coronella austriaca) .
Основным отличительным признаком вида является наличие двух пятен по бокам головы: белых (2,6%), желтых (7,9%), желто-оранжевых (50,0%), ярко-оранжевых (36,8%) или красных (2,7%), которые могут быть слитными (10,5%), но чаще раздельные (89,5%). Окраска спинной стороны большинства змей оливковая (55,6%), также может быть темно-серой (19,4%) или черной (25,0%). 7,1% рептилий на дорсальной стороне без рисунка, тогда как для 85,7% особей характерен рисунок в виде белых (57,1%) или желтых (42,9%) вкраплений. Окраска брюшной стороны представлена темно-серым (36,4%) или черным (63,6%) фоном с рисунком в виде пятен или полос желтого (22,7%) или белого (77,3%) цвета. Внешние морфологические подвидовые различия проявляются в степени выраженности и цвета височного пятна, вариации рисунка и окраски тела, количестве брюшных и подхвостовых щитков. Анализ окраски затылочных пятен змей в зависимости от подвида показал, что доминирующим для номинативного подвида является желтооранжевое окрашивание (54,8%), а редким проявлением – беловатое и красное (3,2%), на остальные морфы (желтые, ярко-оранжевые пятна) приходится 9,7% и 29,1% соответственно. Диапазон вариаций рассматриваемого признака восточного подвида (N. n. scutata) заметно меньше и преобладают яркие тона: ярко-оранжевый (71,4%) и желто-оранжевый (28,6%), при этом белая желтая и красная вариации окраски отсутствуют. Все отмеченные нами змеи второго подвида были с раздельными пятнами. В окраске спинной стороны тела западной формы преобладают оливковые или темно-серые тона (65,5% и 24,1% соответственно), а черная морфа является редкой (10,4%). Для N. n. scutata характерно преобладание темных форм (черная окраска) – 85,7%, а редкой оливковая (14,3%), темно-серая окраска спинной стороны не обнаружена. Рисунок дорсальной стороны также показывает некоторые различия. Так 90,4% N. n. natrix с рисунком в виде белых (71,4%) или желтых (28,6%) вкраплений, 9,6% змей лишены рисунка. Для N. n. scutata количество особей в выборке без рисунка увеличивается почти в 3 раза: 28,6%, доля ужей с рисунком составляет 71,4%, однако доминируют желтые вкрапления (57,1%). 85,7% рептилий номинативного подвида имеют черную окраску вентральной стороны тела и рисунок из белых полос и вкраплений, а 14,3% темно серый фон с желтым рисунком, тогда как черная окраска брюха у восточного подвида (N. n. scutata) отмечена у 53,3% особей, а темно-серая в 52,7% случаев; желтый рисунок характерен для 26,7%, а белый – для 73,3% змей.
Максимальная длина тела ужа обыкновенного на изучаемой территории составила 840мм (табл.) при среднем значении 428±14,3 мм, при этом в зависимости от пола этот показатель изменяется в пределах 184-840 мм для самок и 241-675 мм для самцов. Достоверных отличий самок от самцов по данному показателю не выявлено, но приближается к таковому у западных микропопуляций.
Длина хвоста в 3-4 раза меньше длины тела и варьирует от 40-170 мм у самок и 87-171 мм у самцов. По данным [5] половой диморфизм проявляется в соотношении длины тела и хвоста змеи: L/Lcd, Lcd/L, L/(L-Lcd), что характерно и для волгоградской популяции независимо от подвида. Вокруг середины тела у ужа обыкновенного 15-20 чешуй, у самцов значение данного показателя несколько смещено в сторону максимального значения (16-20 чешуй). Вдоль брюха располагается 165-186 чешуй и 53-73 пары подхвостовых. Вокруг глаза ужа находится 4-5 щитков, при этом 4 щитка отмечено у 11,8%, а 5 у 88,2% особей, верхнегубных 5-8, образуют следующие сочетания: 5/5, 6/6, 6/7, 7/6, 7/7, 7/8, 8/8, из которых наиболее распространенной является 7/7 (46,7%), реже встречаются 6/7 и 6/6
(15,0%), 5/5 (10,0%), 7/6, 7/8 (5,0%), 8/8 (3,3%). Таким образом, асимметричные проявления в выборке отмечены в 25,0% случаев, что говорит о высокой лабильности показателя, однако коэффициент вариации составляет 9,28±0,01.
Таблица. Морфометрические показатели N. n. natrix и N. n. scutata Волгоградской области
|
Признак |
пол |
N.n. natrix min-max / M±m, мм |
T st |
N.n. scutata min-max / M±m, мм |
||
|
N.n.n. (самки/ самцы) |
N.n.n./ N.n. s. |
N.n. s. (самки/ самцы) |
||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
L |
самки |
442-840/ 430,1±15,62 |
2,97 |
1,52 |
0,73 |
184-556/ 392,7±19,1 |
|
самцы |
395-639/ 380,1±6,27 |
0,25 |
241-675/ 377,5±8,2 |
|||
|
L cd |
самки |
40-170/ 126,4±1,67 |
2,40 |
2,63 |
1,95 |
41,1-135,4/ 118,0±2,74 |
|
самцы |
108,5-171,1/ 140,9±5,8 |
2,22 |
87-149/ 126,2±3,20 |
|||
|
L cd /L |
самки |
0,16-0,25/ 0,23±0,001 |
3,57 |
0,00 |
3,57 |
0,20-0,25/ 0,23±0,01 |
|
самцы |
0,27-0,36/ 0,28±0,01 |
0,00 |
0,27-0,32/ 0,28±0,01 |
|||
|
Sq. |
самки |
15-20/ 18,7±0,24 |
1,70 |
5,52 |
1,44 |
16-19/ 17,1±0,12 |
|
самцы |
18-20/ 19,2±0,17 |
5,67 |
16-20 17,5±0,25 |
|||
|
Ventr. |
самки |
165-180/ 179,0±1,73 |
0,97 |
0,25 |
0,67 |
173-186/ 178,5±1,01 |
|
самцы |
171-182/ 180,9±0,92 |
1,00 |
172-185 179,5±1,10 |
|||
|
Scd. |
самки |
53-73/ 62,9±1,09 |
3,92 |
3,48 |
4,15 |
55-62/ 57,5±1,1 |
|
самцы |
65-70/ 68,1±0,80 |
3,90 |
54-68 63,4±0,90 |
|||
|
Scd./Ventr |
самки |
0,32-0,41/ 0,33±0,01 |
4,26 |
0,71 |
3,57 |
0,31-0,40/ 0,34±0,01 |
|
самцы |
0,37-0,40/ 0,39±0,01 |
0,00 |
0,36-0,40/ 0,39±0,01 |
|||
|
Количество чешуй вокруг глаза |
самки |
4-5/ 5,0±0,01 |
- |
6,33 |
2,00 |
4-5/ 4,8±0,03 |
|
самцы |
5 |
- |
4-5/ 4,9±0,04 |
|||
|
Lab |
самки |
5-8/ 6,3±0,13 |
3,16 |
3,76 |
4,81 |
6-7/ 6,8±0,03 |
|
самцы |
5-7/ 6,8±0,09 |
3,70 |
5-7/ 6,3±0,10 |
|||
|
Lab |
самки |
5-8/ 6,3±0,13 |
3,16 |
3,76 |
4,81 |
6-7/ 6,8±0,03 |
|
самцы |
5-7/ 6,8±0,09 |
3,70 |
5-7/ 6,3±0,10 |
|||
|
Sub. Lab. |
самки |
7-10/ 8,3±0,18 |
1,94 |
0,97 |
0,63 |
7-10/ 8,6±0,25 |
|
самцы |
7-9/ 8,7±0,10 |
0,45 |
7-10/ 8,8±0,20 |
|||
|
Предглазничные |
самки |
1 |
- |
- |
2,00 |
1-2/ 1,3±0,03 |
|
самцы |
1 |
- |
1-2/ 1,2±0,04 |
|||
|
Заглазничные |
самки |
2-3/ 2,8±0,03 |
- |
2,38 |
2,38 |
2-3/ 2,7±0,03 |
|
самцы |
3 |
- |
2-3/ 2,8±0,03 |
|||
Нижнегубные щитки отмечены в следующих комбинациях: 7/7, 8/8, 8/9, 9/7, 9/9, 10/9, 9/10, 10/8, 10/10, из которых наиболее распространены 9/9
(30,5%) и 10/10(16,9%), остальные же 6,8%, 8,5%, 10,2%, 5,1%, 10,2%, 8,5%, 3,4% соответственно.
В типичном случае (92,3%) предглазничных щитков насчитывается 1 с правой и левой стороны, однако в ряде случаев их может быть 2 (7,7%) и свойственны для заволжских популяций. Число щитков относительно стабильно (CV = 8,43±0,01), но по данному признаку возможна асимметрия, отмеченная в комбинациях 1/2 и характерная для 15,8% ужей подвида N. n. scutata. Заглазничных щитков больше: от 2 до 3 и они более стабильны (CV = 4,84 ±0,01). 2 щитка отмечено у 18,3%, а 3 – у 81,7%, асимметрия встречается реже, чем среди предыдущих (по 2,5%, приходящиеся на вариации 2/3 и 3/2).
Учет подвидовых особенностей размерных характеристик и признаков фолидоза показал достоверные отличия для числа чешуй вокруг середины тела, подхвостовых, верхнегубных и щитков вокруг глаза, из которых наиболее надежным отличием характеризуется первый и последний показатели (T st = 3,71 и 6,33 соответственно).
Анализ традиционных метрических и меристических показателей позволил выявить по ряду признаков половой диморфизм: по отношению длины хвоста к длине тела (L cd /L) выявлено достоверное отличие между самками и самцами, для первых он составляет 0,16-0,25, а для последних – 0,27-0,36 (Tst=3,57), число подхвостовых чешуй у самок имеет более выраженный диапазон изменчивости, но в среднем их больше у самцов как у N. n. natrix так и N. n. scutata . Достоверных отличий по отношению подхвостовых щитков к брюшным между западными и восточными популяциями не выявлено, но для тех и других свойственен половой диморфизм, который в большей степени проявляется у N. n. natrix : самки отличаются меньшей величиной (0,320,41/0,33±0,01 – N. n. natrix и 0,31-0,40/0,34±0,01 N. n. scutata ), чем самцы (0,37-0,40/0,39±0,01– N. n. natrix и 0,36-0,40/0,39±0,01 N. n. scutata ). Верхнегубных щитков больше у самок заволжских популяций (6,8±0,03 – самки и 6,3±0,10 самцы), у N. n. na-trix наблюдается обратное – самцы несут больше верхнегубных щитков (6,3±0,13– самки и 6,8±0,09 самцы).
Таким образом, в биологическом и экологическом отношении уж обыкновенный, обитающего на территории Волгоградской области, сходен с популяциями других регионов России. Однако морфологические в, зависимости от широты обитания, признаки претерпевают некоторые изменения: так среднее число брюшных щитков у самцов волгоградской популяции меньше, чем у саратовской, но больше калмыцкой [9], обратная картина наблюдается у самок; диапазон изменчивости чешуй вокруг середины тела на изучаемой территории выше, чем в северных регионах. Вместе с тем, особенности метрических и меристических признаков не выходят за пределы видовой изменчивости.
Список литературы Особенности распространения, биологии, экологии и морфологии ужа обыкновенного Natrix natrix (Linneaus, 1758) Волгоградской области
- Ананьева Н.Б., и др. Земноводные и пресмыкающиеся. Энциклопедия природы России. М.: Изд-во «ABF», 1998. С. 494-496.
- Ананьева Н.Б., и др. Атлас пресмыкающихся Северной Евразии (таксономическое разнообразие, географическое распространение и природоохранный статус)/Зоол. ин-т РАН. СПб., 2004. С. 164-165.
- Банников А.Г., и др. Определитель земноводных и пресмыкающихся фауны СССР. М.: Просвещение, 1977. 414 c.
- Морозенко Н.В. Эколого-морфологическая структура и фенетический анализ популяций обыкновенного ужа (Reptilia; colubridae, natrix) Нижнего Поволжья: Дисс.... канд. биол. наук. Саратов, 2003. 199 с.
- Шляхтин Г.В., Голикова В.Л. Методика полевых исследований экологии амфибий и рептилий. Саратов: Изд-во Сарат. ун-та, 1986. 78 с.
- Табачишин В.Г., Табачишина И.Е. Распространение и особенности экологии обыкновенного ужа (Natrix natrix) на севере Нижнего Поволжья//Поволж. экол. журн. 2002. № 2. С. 179-183.
- Табачишина И.Е. Эколого-морфологический анализ фауны рептилий севера Нижнего Поволжья. Дис.... канд. биол. наук: Саратов, 2004 182 с.
- Вершинин В.Л. Амфибии и рептилии Урала. Екатеринбург: УрО РАН, 2007. 170 с.
- Ждокова М.К. Эколого-морфологический анализ фауны амфибий и рептилий Калмыкии. Дисс.... канд. биол. наук. Саратов, 2003. 284 с.
