Особенности размножения и структуры популяции лесной мышовки (Sicista betulina Pall.) на северном пределе ареала
 на северном пределе ареала")
Автор: Ивантер Эрнест Викторович, Кухарева Анна Вячеславовна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 6 (111), 2010 года.
Бесплатный доступ
Мышовка, популяция, репродукция, плодовитость, структура популяции
Короткий адрес: https://sciup.org/14749775
IDR: 14749775
Текст статьи Особенности размножения и структуры популяции лесной мышовки (Sicista betulina Pall.) на северном пределе ареала
Будучи единственным представителем надсемейства тушканчикообразных, обитающим преимущественно в таежной зоне и, в отличие от всех других мелких млекопитающих этого региона, впадающим в зимнюю спячку, лесная мышовка, тем более в условиях северной границы ареала, представляет для исследователей особый интерес. В основу настоящей статьи, посвященной особенностям размножения и экологической структуры популяции этого вида на северном пределе распространения, положены материалы, собранные в 1958–2009 годах в процессе многолетних стационарных (заповедник «Кивач», Приладожский стационар Карку, окрестности Кончезерской биостанции) и экспедиционных исследований, охвативших всю территорию южной половины Карелии.
Для изучения размножения лесных мышовок популяцию этих зверьков мы подразделяли на возрастные группы. Для их выделения послужил основанный на рекомендациях Г. А. Клевезаль [8] метод возрастной диагностики мышовок [4], [5], заключающийся в подсчете годовых слоев (линий склеивания) на окрашенных гематоксилином Эрлиха поперечных срезах нижней челюсти в периостальной зоне кости (рис. 1).
Точное определение возраста зверьков и анализ состояния их половой системы показывают, что в условиях таежного Северо-Запада России лесные мышовки впервые начинают размножаться в возрасте около года, после первой зимовки. Никаких изменений в гонадах сеголеток, указывающих на возможность их созревания и участия в размножении в год своего рождения, не обнаружено. Размеры семенников не превышают у них 5,0 х 3,3 мм, а вес – 57 мг (табл. 1). Инфантильная матка имеет рога до 17, влагалище – до 10,5 мм.
У взрослых мышовок размеры половых органов в несколько раз больше. Вес двух семенников – 33– 250 мг, длина семенника – 3,2–12,1 мм, длина рогов матки – 9,0–32, влагалища – 2,5–18,7 мм. Семенные пузырьки выражены только у зимовавших особей. О позднем половом созревании лесных мышовок (отсутствии размножения молодых, созревающих лишь на следующий год после рождения) сообщают и другие исследователи [13], [15], [21], [1], [11], [3], [20].
О сроках и интенсивности размножения лесной мышовки в Карелии можно судить по результатам специального гистологического исследования половых циклов самцов и самок. В начале июня, когда у этого вида начинается сезон репродукции, функциональное состояние половых органов самцов характеризуется активным сперматогенезом. Внешне это выражается в значительном увеличении веса семенников (к июлю – до 177 мг) и их размеров (до 7,2–7,9 мм к концу июня), а также в развитии придаточных желез. В семенных канальцах (средний диаметр – 192 мкм) присутствуют все клеточные элементы от клеток Сертоли и сперматогониев до зрелых сперматозоидов, собирающихся в просветах канальцев.
Гистологическая картина сперматогенеза и сезонные изменения размеров гонад (рис. 2 и 3) показывают его стабильную активность в период с начала – середины июня и до конца июля, то есть в течение большей части периода бодрствования, отсутствие каких-либо подъемов и спадов, соответствующих периодам спаривания и паузам между ними. Постепенное затухание сперматогенеза и дегенерация зародышевого эпителия до однорядного слоя сперматогониев и клеток Сертоли начинаются в августе, но в массе происходят, очевидно, только в сентябре, перед впадением в спячку.
У молодых самцов гонады находятся в состоянии полного покоя, генеративный эпителий содержит лишь клетки Сертоли и сперматогонии (рис. 3).
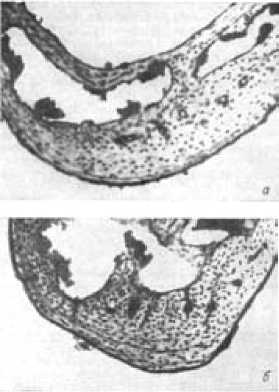
Рис. 1. Поперечные срезы нижней челюсти мышовок разного возраста: а – в возрасте 1 года; б – 2 лет; в – 4 лет.
Окраска гематоксилином Эрлиха, × 60
Таблица 1
|
Характеристика половой системы лесной мышовки |
||
|
Показатели |
Зимовавшие |
Прибылые, август |
|
июнь июль август |
||
Самцы
|
число исследованных зверьков длина семенника, мм |
208 6,7 (5,0 – 11,0) |
89 6,4 (4,8 – 12,1) |
21 4,8 (3,2 – 6,0) |
55 3,9 (3,0 – 5,0) |
|
число исследованных зверьков |
202 |
85 |
19 |
7 |
|
длина семенных пузырьков, мм |
8,4 (3,5 – 11,7) |
7,8 (4,5 – 11,0) |
3,0 (2,5 – 4,5) |
2,5 (1,0 – 3,6) |
|
число исследованных зверьков |
78 |
61 |
17 |
21 |
|
вес двух семенников, мг |
176 (114 – 250) |
150 (69 – 205) |
68 (33 – 130) |
36,1 (22 – 57) |
Самки
|
число исследованных зверьков |
38 |
33 |
14 |
28 |
|
длина рогов матки, |
18,2 (11,0 – |
19,4 (9,0 – |
17,8 (12,3 – |
11,2 (6,0 – |
|
мм |
27,6) |
32,0) |
23,6) |
17,0) |
|
число исследованных |
19 |
26 |
14 |
19 |
|
зверьков |
5,4 (2,7 – |
8,8 (3,8 – |
8,9 (2,5 – |
4,0 (1,0 – |
|
длина влагалища, мм |
11,2) |
17,4) |
18,7) |
10,5) |
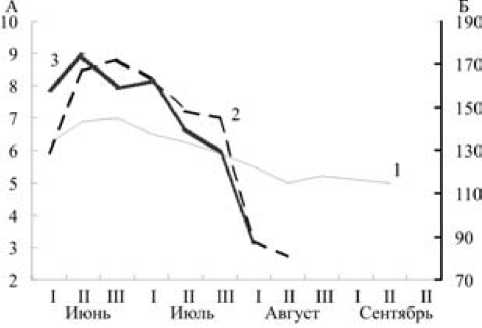
Рис. 2. Средняя длина семенника (1), семенных пузырьков (2) и средний вес двух семенников (3) у лесных мышовок по декадам. По оси абсцисс – декады (римские цифры);
по оси ординат А – длина, мм; по оси ординат Б – вес, мг
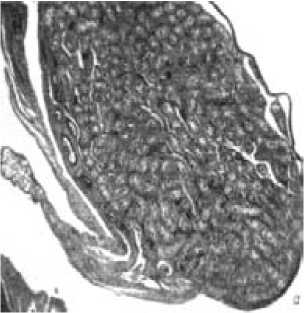
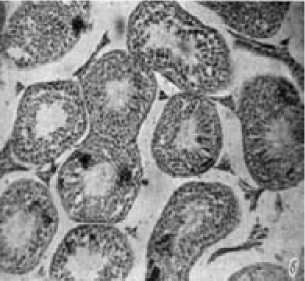
Рис. 3. Поперечные срезы извитых канальцев семенника молодого самца лесной мышовки на стадии покоя (а) и взрослого в период сперматогенеза (б). Микрофото, × 400
Изучение эстрального цикла самок (рис. 4) показало, что лесная мышовка – моноцикличе-ский вид, лишь однажды за лето приходящий в состояние эструса, причем по времени эструсы самок соответствуют активному сперматогенезу самцов. В июне половые органы самок увеличены, а в июле достигают максимальных размеров (табл. 2). Особенно резко возрастает вес яичников (до 10–12 мг). В яичниках много фолликулов на разных стадиях развития, от примордиальных до граафовых пузырьков, диаметром от 130 до 300 мкм. Ткань яичника богата кровеносными сосудами. Некоторые полостные фолликулы близки к разрыву. Эпителий слизистой влагалища многорядный, в верхних слоях хорошо заметно ороговение клеток, происходит их усиленная дисквамация. У беременных самок (в июне – июле) яичник содержит желтые тела беременности (их максимальный диаметр – 258 мкм) и атрезии. Матка гиперемирована и на поперечном разрезе имеет звездчатую форму, в просвете – секрет маточных желез. После родов рост фолликулов замедляется, прекращается он на стадии образования многослойного зачаткового эпителия; одновременно увеличивается число атрезирующих фолликулов.
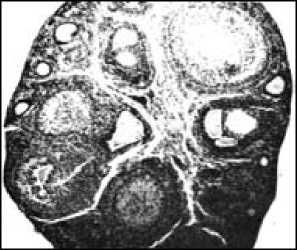
а
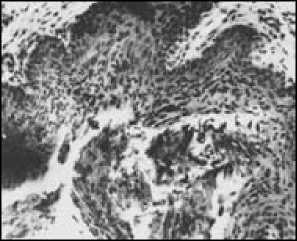
б
Рис. 4. Срезы половых органов взрослой самки лесной мы-шовки: а – яичник в период интенсивной фолликулярной деятельности; б – влагалище во время эструса
Общий ход и интенсивность размножения самок лесной мышовки по декадам представлены в табл. 2. Согласно этим данным, во второй декаде июня (до этого времени взрослые самки нам не попадались) около трети взрослых самок были беременны, остальные или еще не размножались (64 %), или уже завершили кормление недавно появившегося выводка (7 %). К концу июня число беременных (42 %), лакти-рующих (5 %) и рожавших (26 %) самок значительно возрастает, а без следов размножения, напротив, сокращается (26 %). В первой декаде июля более 90 % особей принимают участие в размножении, при этом процент беременных достигает максимума (60 %), затем их доля по- степенно снижается. К концу месяца большинство самок уже обзаводятся потомством (78 %) и либо приступают к лактации (43 %), либо уже откормили выводок (36 %). В августе отлавливались только кормящие самки (32 %), остальные уже закончили размножение. Все это говорит о том, что в условиях Карелии лесные полевки приносят за лето только один выводок (об этом говорит и одновершинный характер кривой уловов молодых), причем массовое рождение детенышей приходится на первую половину июля.
Если учесть, что беременность длится у этого вида 25–30 дней [22], то общая картина размножения лесной мышовки в Карелии представляется следующим образом. Массовый гон и спаривание происходят в первой половине июня, рождение детенышей – в первой-второй декадах июля, а их массовое расселение (переход к самостоятельной жизни и выход из гнезд) – в августе, то есть через 30–40 дней после рождения. Это соответствует продолжительности гнездового развития (35–37 дней), установленной в результате прямых наблюдений за жизнью новорожденных [1], [17].
Характерные сезонные изменения численности мышовок [4], [6], [7] свидетельствуют о сравнительно позднем их размножении в Карелии. Вследствие этого здесь наблюдаются и более поздние выход и расселение молодых (не ранее последней декады июля), тогда как в более южных районах их начинают ловить почти на месяц раньше [23], [13], [10], [19]. Лишь в средней Сибири размножение этого вида проходит в близкие с Карелией сроки [14], [3], [11].
Таблица 2
Интенсивность размножения взрослых самок лесной мышовки
|
Месяц и декада |
Общее число исследованных самок |
Относительное количество самок (% от общего числа) |
|||
|
беременных |
кормящих |
рожавших |
без следов размножения |
||
|
Июнь |
|||||
|
II |
14 |
28,6 |
– |
7,1 |
64,3 |
|
III |
19 |
42,1 |
5,3 |
26,3 |
26,3 |
|
II–III |
33 |
36,4 |
3,0 |
18,2 |
42,4 |
|
Июль |
|||||
|
I |
25 |
60 |
4 |
28 |
8 |
|
II |
28 |
39,3 |
17,9 |
14,3 |
58,6 |
|
III |
14 |
7,1 |
42,9 |
35,7 |
14,3 |
|
I–III |
67 |
40,3 |
17,9 |
23,9 |
17,9 |
|
Август |
|||||
|
I |
9 |
– |
55,6 |
33,3 |
11,1 |
|
II |
8 |
– |
25 |
50 |
25 |
|
III |
5 |
– |
– |
– |
100 |
|
I–III Сен- |
22 |
– |
31,8 |
31,8 |
36,4 |
|
тябрь (I–III) |
8 |
– |
– |
– |
100 |
Таблица 3
Вариации величины выводка у лесной мышовки в Карелии
|
Месяц |
Общее число беременных самок |
Количество самок (экз.) с числом эмбрионов |
Среднее число эмбрионов на самку |
|
3 4"1 5 "1 6"1 7 8 9 10 11 |
По всему материалу
|
Июнь |
12 |
2 |
3 |
5 |
– |
1 |
– |
– |
– |
1 |
5,1 (2,2 + 2,9) |
|
Июль |
35 |
2 |
13 |
7 |
9 |
3 |
– |
1 |
– |
– |
5,1 (2,6 + 2,5) |
|
Август |
6 |
– |
1 |
3 |
2 |
– |
– |
– |
– |
– |
5,2 (2,8 + 2,4) |
|
Всего |
53 |
8 |
21 |
20 |
11 |
4 |
– |
1 |
– |
1 |
5,1 (2,6 + 2,5) |
Из них: самки в возрасте 1 года
Июль 21 21 I 1 ] 4 ] 8 i 6 ] 1 I –1 ] – – 5,3 (2,9 + 2,4)
самки в возрасте 2 лет и старше
Июль i 8 П 2 1 51– 11– – – – – 4,0 (2,0 + 2,0)
Примечание. В скобках указано число эмбрионов в правом и левом рогах матки.
Сроки размножения лесной мышовки меняются по годам в зависимости от метеорологической обстановки, и прежде всего от хода весны. При раннем наступлении весенних явлений, теплой и сухой погоде в мае – июне размножение зверьков начинается раньше и проходит в более сжатые сроки. Напротив, в годы с поздней, затяжной и холодной весной размножение сильно задерживается и растягивается. Наиболее раннее начало размножения лесной мышовки наблюдалось в Карелии в 1959 году, когда переход среднесуточной температуры через 0 и 5º был отмечен соответственно 13 и 26 апреля. Гон у мышовок проходил в тот год в начале июня, первые беременные самки встречены 7 и 9 июня, а первые молодые – в конце июля (26.07). Почти на полмесяца позднее началось размножение в 1951 и 1958 годах. Переход среднесуточной температуры через 0 и 5º в 1951 году произошел 25 апреля и 19 мая, в 1958 году – 28 апреля и 22 мая. В связи с этим задержалось и размножение. Первые беременные самки были добыты в самом конце июня (27.07.1951 и 25.07.1958), первые молодые – во второй декаде августа (12.08.1951, 18.08.1958).
Гораздо стабильнее интенсивность размножения. Доля участвующих в нем перезимовавших самок почти не меняется по годам и составляет 98–100 %. Несущественны различия и в величине разовой плодовитости. Таким образом, в отличие от ряда других видов мелких млекопитающих Карелии, годовые изменения численности лесной мышовки обусловлены в основном экзогенными факторами. Размножение этого вида характеризуется невысоким темпом (размножаются только взрослые особи, и к тому же всего 1 раз в год), и поэтому не может служить эффективным регулятором его численности.
С другой стороны, популяция мышовок в Карелии отличается общей невысокой плотностью и даже в годы пиков численности не достигает критической величины. В этих условиях вряд ли есть необходимость в выработке внутрипопуляцион- ных механизмов автоконтроля. Их отсутствие является следствием общей невысокой численности вида у северной границы ареала и одновременно причиной резких колебаний его плотности на этом фоне. Не исключено, что в отдельных участках ареала с оптимальными условиями, где средний многолетний уровень численности вида в несколько раз выше, чем на Северо-Западе России, механизмы эндогенной саморегуляции все же имеют место. Иначе трудно объяснить cтабильность численности популяции при высоком уровне ее плотности и отсутствие этой стабильности при низкой плотности популяции. В первом случае численность может регулироваться за счет изменения числа участвующих в размножении взрослых самок (в Татарии и Кировской области в некоторые годы отмечалось до 10 и даже до 70 % яловых самок) [13], [10], а во втором – с этой задачей вполне справляются внешние факторы (например, климатические условия), и интенсивность размножения не меняется по годам.
Средний размер выводка у лесных мышовок Карелии на основании подсчета эмбрионов у 53 беременных самок составил 5,1 + 0,2 с колебаниями от 3 до 11 (табл. 3). Это несколько ниже, чем в Кировской области, Республике Коми и Татарии, где средний показатель плодовитости равнялся соответственно 5,5, 5,9 и 5,5 (в Волжско-Камском заповеднике, по данным И. В. Назаровой и В. В. Мовчаренко [13], эта величина составила 6,4), но выше, чем в предгорьях Алтая (4,3 эмбриона на самку), долине р. Верхняя Ангара (3,7) и Ивановской области (4,0) [13], [10], [11], [3], [18].
Среднее число эмбрионов, приходящихся на 1 самку, в течение репродуктивного периода практически не меняется. Наибольшей плодовитостью отличаются первородящие годовалые самки, с возрастом она снижается (табл. 3). Различий между средним числом эмбрионов и плацентарных пятен, на что указывал для Татарии В. А. Попов [13], на нашем материале не обнаружено: на 1 беременную самку приходится в среднем 5,11 + 0,30 эмбриона, а на 1 рожавшую – 5,16 + 0,32 плацентарных пятна. Разница статистически недостоверна (р < 0,01). Это не значит, что в наших условиях у мышовок вообще нет рассасывания эмбрионов. Просто автолиз может проходить на более ранних этапах эмбриогенеза и данным методом не фиксироваться. Кроме того, мы имеем и прямое подтверждение существования у них автолиза: у одной из беременных самок обнаружен явно отстающий в росте и рассасывающийся эмбрион. Но одно бесспорно: масштабы эмбриональной гибели у мышовок Карелии незначительны, во всяком случае, в последние 2–3 недели беременности. Что же касается распределения эмбрионов по рогам матки, то оно практически равномерное (различия в пределах ошибки).
Обращает внимание весьма значительная асимметричность кривой распределения самок по числу эмбрионов в выводке (показатель «косо-сти» А равен +1,5; m = 0,39; t = 4,4). Это указыва- ет на существование в популяции лесных мышо-вок Карелии отчетливой тенденции к увеличению разовой плодовитости и свидетельствует о том, что процесс эволюционного отбора на повышенную плодовитость (призванную компенсировать повышенную в периферических популяциях смертность) еще не завершен: уровень плодовитости еще не достиг оптимального для данных условий уровня. Мышовка проникла на север сравнительно недавно, и адаптация ее к местным условиям, по-видимому, еще продолжается.
Половой состав отловленных взрослых мы-шовок отражает изменение активности зверьков разного пола в связи с размножением (табл. 4, рис. 3 и 4). В первой декаде июня ловятся исключительно самцы, во второй половине месяца они составляют около 90 %, а затем, обычно ко второй половине июля, по мере снижения половой активности самцов и освобождения самок от забот о выводке, соотношение полов выравнивается. В августе оно снова уклоняется от 1:1, но уже в пользу самок (70,6 %). К этому времени они уже откормили выводок и активность их, а вместе с ней и отлов, увеличивается.
Общая доля самцов в уловах взрослых зверьков составила по всему материалу 74,1 %, причем сопоставление полученных величин хи-квадрат с критической (3,84) говорит о большой статистической достоверности отмеченной диспропорции. Приблизительно такая же картина наблюдается и в других частях ареала [16], [23], [13], [12], [3].
В уловах молодых мышовок с июля по сентябрь также преобладали самцы, но их количественное доминирование не столь существенно (табл. 5). Тем не менее высказанное нами ранее [4], [6], [7] предположение о том, что это связано не с большей их активностью, а с количественным преобладанием в популяции вследствие более частого рождения особей мужского пола, остается в силе. К близкому выводу приходят и другие исследователи [13], [15], [11]. Это позволяет отнести лесную мышовку к видам со стойким численным преобладанием самцов [9], [2].
Возрастной состав популяции мышовок меняется по сезонам (табл. 5, рис. 5 и 6). До конца июля в ловушки и канавки попадают почти исключительно взрослые перезимовавшие особи, а начиная с первой декады августа все больший удельный вес в уловах занимают молодые зверьки. В начале месяца их не более 40 %, в середине и конце месяца – 84 %, а в сентябре – 90–100 %. Ко второй декаде сентября взрослые мышовки почти полностью исчезают и ловятся только прибылые зверьки, родившиеся в текущем году. При этом сокращается не только относительная (доля в уловах), но и абсолютная численно сть взрослых животных. Такое резкое изменение возрастной структуры популяции мышовок в конце лета связано как с процессами естественной смертности взрослых особей, так и с более ранним уходом их на зимовку. Последнее характерно, по-видимому, и для зверьков других популяций [10], [17]. Сроки появления молодых мышовок в отловах варьируют в разных частях ареала. В долине р. Верхняя Ангара они попадаются с конца июля, в тайге Сосьвинского Приобья – несколько позже (в начале августа), причем особенно резкое увеличение их количества в отловах произошло в третьей декаде этого месяца [11]. В южных же частях ареала (Татария, Оренбургская область, предгорья Алтая) прибылые зверьки начинают встречаться почти на месяц раньше, с первых чисел июля [13], [10], [19].
Большой интерес представляет анализ возрастного состава взрослой части популяции (табл. 5). Благодаря применению специальной методики определения возраста мышовок (по слоистым структурам кости нижней челюсти) удалось установить, что основу группы зимовавших зверьков составляют особи, родившиеся в прошлом году (80 %), далее следуют мышовки, пережившие две зимы (16 %), и совсем редки особи трех лет и старше (4 %). В августе – сентябре, перед уходом на зимовку, популяция имеет следующую структуру: прибылые составляют около 80 %, годовики – 16,4, двухлетки – 2,7, трехлетние и более старые – 0,9 %. Таким образом, численность каждой следующей возрастной группы сокращается по сравнению с предыдущей в 5–6 раз, или в среднем на 80 %. Эти 80 % и составляют величину годовой смертности населения мышовок, причем,
|
судя по тому, что количественное соотношение между соседними возрастными группами довольно стойко сохраняется, темп смертности у них приблизительно одинаков. |
||||||||
|
Возрастной и |
Таблица 4 половой состав популяции |
|||||||
|
лесной |
мышовки |
в Карелии (по |
данным отлова) |
|||||
|
Месяц |
Число зимовав- |
Число прибы- |
Общее число |
|||||
|
ших |
лых |
зверьков |
||||||
|
абс. |
из них |
χ2 |
абс. |
из них |
χ 2 |
абс. |
из них зимо- |
|
|
самцы, |
самцы, |
вавшие, % |
||||||
|
% |
% |
|||||||
|
Июнь |
275 |
88,4 |
162,0 |
– |
– |
– |
275 |
100 |
|
Июль |
171 |
58,5 |
4,9 |
12 |
58,3 |
0,3 |
183 |
93,4 |
|
Август |
33 |
36,4 |
2,5 |
145 |
53,1 |
0,6 |
178 |
18,5 |
|
Сентябрь |
3 |
66,6 |
0,3 |
30 |
63,3 |
2,1 |
33 |
9,1 |
|
Всего |
482 |
74,1 |
111,7 |
187 |
55,1 |
1,9 |
669 |
72,0 |
Таблица 5
Возрастная структура популяции лесной мышовки (по исследованию черепов)
|
Возрастные группы |
По слоистым структурам |
По стертости зубов |
||
|
n |
% |
n |
% |
|
|
Сеголетки |
49 |
32,9 |
76 |
45,5 |
|
Годовики |
80 |
53,0 |
66 |
39,5 |
|
Двухлетки |
16 |
10,7 |
24 |
14,4 |
|
Трехлетки |
3 |
2,0 |
1 |
0,6 |
|
Четыре года и старше |
1 |
0,7 |
0 |
0 |
|
Всего |
149 |
100 |
167 |
100 |
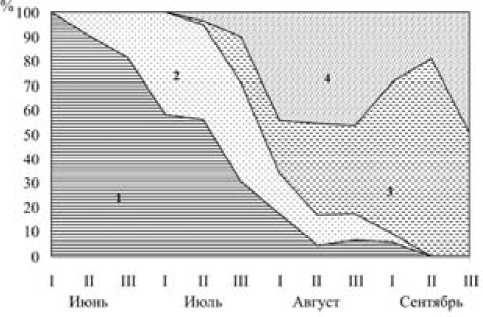
Рис. 5. Изменения половой и возрастной структуры популяции лесной мышовки по данным отлова:
1 – зимовавшие самцы; 2 – зимовавшие самки; 3 – прибылые самцы; 4 – прибылые самки. По оси абсцисс – декады; по оси ординат – доля улова, %
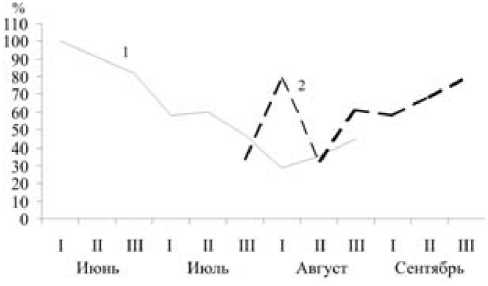
Рис. 6. Изменение доли (%) самцов у взрослых (1) и молодых (2) лесных мышовок по декадам
Список литературы Особенности размножения и структуры популяции лесной мышовки (Sicista betulina Pall.) на северном пределе ареала
- Айрапетьянц А. Э. К биологии лесной мышовки в Ленинградской области//Вопросы экологии и биоценологии. Вып. 9. Л.: ЛГУ, 1969. С. 117-124.
- Большаков В. Н., Кубанцев Б. С. Половая структура популяций млекопитающих и ее динамика. М.: Наука, 1984. 232 с.
- Буйдалина Ф. Р. Лесная мышовка (Sicista betulina Pall.) в тайге Сосьвинского Приобья//Фауна, таксономия, экология млекопитающих и птиц. Новосибирск: Наука. СО АН СССР, 1987. С. 41-43.
- Ивантер Э. В. К экологии лесной мышовки (Sicista betulina Pall.)//Aquilo. Ser. Zool. 13: 1972. С. 103-108.
- Ивантер Э. В. Методика определения возраста лесной мышовки, Sicista betulina (Rodentia, Dipodoidea)//Зоол. журн. 1973. Т. 52. № 2. С. 255-257.
- Иванте р Э. В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.: Наука. ЛО, 1975. 246 с.
- Ивантер Э. В. К экологии лесной мышовки (Sicista betulina Pall.)//Зоол. журн. 2007. Т. 86. С. 476-493.
- Клевезаль Г. А. Регистрирующие структуры млекопитающих в зоологических исследованиях. М.: Наука, 1988. 286 с.
- Кубанцев Б. С. О половом составе популяций у млекопитающих//Журн. общ. биол. 1972. Т. 33. № 2.
- Кулик И. Л., Тупикова Н. В., Никитина Н. А., Карасева Е. В., Суворова Л. Г. Материалы по экологии лесной мышовки (Sicista betulina Pall.)//Сб. тр. зоол. музея МГУ. Вып. 10. М.: МГУ, 1968. С. 146-160.
- Лямкин В. Ф., Малышев Ю. С., Пузанов В. М. К экологии лесной мышовки на Северо-Восточной оконечности ареала//Грызуны: Материалы VI Всесоюзного совещ. Л.: Наука, 1983. С. 328-329.
- Плешак Т. В., Козловский И. С. К экологии лесной мышовки//Биология и промысел охотничьих животных. Пермь, 1983. С. 57-61.
- Попов В. А. Млекопитающие Волжско-Камского края. Казань: Казанский филиал АН СССР, 1960. 467 с.
- Реймерс Н. Ф. Птицы и млекопитающие южной тайги Средней Сибири. М.; Л.: Наука, 1966. 420 с.
- Реймерс Н. Ф., Воронов Г. А. Насекомоядные и грызуны Верхней Лены. Иркутск: Иркутское книжн. изд-во, 1963. 191 с.
- Снигиревская Е. М. Грызуны Башкирского заповедника//Тр. Башк. гос. заповедника. Вып. 1. Уфа, 1947. С. 5-16.
- Фокин И. М. Тушканчики. Л.: ЛГУ, 1978. 181 с.
- Хелевина С. А., Окулова Н. М. Экология и морфология лесной мышовки (Sicista betulina Pall.) Ивановской области//Тушканчики фауны СССР: Тез. Всесоюзного совещ. Ташкент: ФАН, 1988. С. 116-119.
- Цветкова А. А. Особенности размножения степной и лесной мышовок на Южном Урале//Экология. 1978. № 3. С. 90-92.
- Шенброт Г. И., Соколов В. Е., Гептнер В. Г., Ковальская Ю. М. Тушканчикообразные. Млекопитающие России и сопредельных регионов. М.: Наука, 1995. 573 с.
- Штильмарк Ф. Р. Основные черты экологии мышевидных грызунов в кедровых лесах Западного Саяна//Фауна кедровых лесов Сибири и ее использование. М.: Наука, 1965. С. 5-52.
- Godfrey G. K. Observation on the nature of the decline in numbers of two Microtus populations//J. Mammal. 1955. Vol. 36. № 2.
- Kubik J. Badania nad morfologia i biologia smuzki (Sicista betulina Pall.) Bialoweskiege Parku Narodowego//Ann. Univ.M. Curie-Sklod. 1952. C. 7.1.
