Особенности размножения птиц-дуплогнездников на северной границе распространения сосновых лесов

Автор: Зацаринный И.В., Константинов В.М.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 353 т.16, 2007 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140150539
IDR: 140150539
Breeding biology of hole-nesting birds in northern limit of pine range
Текст статьи Особенности размножения птиц-дуплогнездников на северной границе распространения сосновых лесов
И. В. Зацаринный1), В. М. Константинов2)
-
1) Рязанский государственный университет, Рязань, 390000, Россия
-
2) Московский педагогический государственный университет, Москва, 129278, Россия
Поступила в редакцию 3 апреля 2007
Изучение гнездовой биологии птиц в северных экосистемах представляет собой важное направление в изучении их экологии. Изучение процессов, протекающих в популяциях у северных границ распространения видов, способствует выявлению адаптаций, позволяющих птицам проникать на север, расширяя ареал. Ранее было выяснено, что существует ряд закономерностей приспособления птиц к обитанию у северных границ распространения. Основные из них касаются фенологии и эффективности размножения. Теперь хорошо известно, что начало размножения птиц при продвижении с юга на север запаздывает на 3-4 суток на каждый градус широты.
Птицы, обитая в экстремальных условиях, в подходящих ситуациях увеличивают продуктивность размножения. Известно, что в северных частях ареалов происходит последовательное увеличение средней величины кладки по сравнению с южными районами.
Дуплогнездники представляют собой экологически обособленную группу птиц, для которых необходимым условием размножения является наличие подходящих укрытий для гнёзд. Так, для факультативных дуплогнездников важно наличие готовых дупел, для облигатных – деревьев с рыхлой древесиной, позволяющей птицам самостоятельно выдолбить дупло. Многочисленными исследованиями установлено, что размещение искусственных гнездовий в лесных стациях с недостатком естественных дупел приводит к увеличению численности птиц, испытывающих в них дефицит, и появлению новых видов дуплогнездников (Зимин 1966, 1968, 1976; Иноземцев 1976; Drent 1987; и др.). Так, отсутствие подходящих укрытий для гнёзд ограничивает плотность населения птиц в гнездовой период даже при обилии доступной пищи (Тима 1957; Бубличенко 2006; и др.). В этом случае проблему несоответствия распределений пищевых и гнездовых ресурсов птицы решают разными способами. Так, неравномерное распределение подходящих для помещения гнёзд укрытий в молодых и средневозрастных лесах приводит к мозаичному, а иногда и «полуколониальному» гнездованию дуплогнездников (Севастьянов 1962; Зимин 1976; Dhondt, Schillemans 1983). Отсутствие необходимых для постройки гнезда условий заставляет этих птиц использовать нехарактерные для данного вида укрытия, а конкуренция за места, пригодные для устройства гнёзд, приводит к откладыванию яиц в одно и то же гнездо или вытеснению одного вида другим (Клитин 1972; Ушакова, Ушаков 1976; Järvinen, Pryl 1977; Бучко, Скільський 1995; Головань 1997; Kucharski 1998; Lesinski 2000). Однако в целом в природе редко возникает ситуация недостатка естественных мест гнездования (Carlson et al. 1998). Поэтому возможно, что увеличение плотности населения птиц при развешивании искусственных гнездовий повышает только локальную плотность гнездования, а гнездящиеся здесь птицы охраняют индивидуальные территории на разном расстоянии от такой «искусственной полуколонии».
Основные исследования по изучению биологии птиц у северных границ распространения лесной зоны Фенноскандии (69º с.ш.) были выполнены в своё время в северной Финляндии (Järvinen 1978, 1983; и др.). Работы по экологии дуплогнездников в северо-западной части Кольского полуострова практически отсутствуют. Целью настоящей работы стало изучение особенностей гнездовой биологии дуплогнездников у северной границы распространения сосновых лесов и выяснение влияния размещения искусственных укрытий на плотность гнездования этих птиц.
Район исследования, материал и методика
Изучение гнездовой биологии дуплогнездников проводили в мае-июле 2003-2006 годов в окрестностях заповедника «Пасвик», расположенного на северо-западе Кольского полуострова, на стыке государственных границ России и Норвегии. Растительность здесь представлена сосновыми лесами, произрастающими у северных пределов распространения сосны Pinus sylvestris . К числу важных особенностей этих лесов относятся: сильная разреженность, небольшая высота деревьев, высокая степень их повреждения.
Основной материал собран при обследованиях искусственных гнездовий, который дополнен данными по гнёздам, найденным в естественных дуплах. В целях привлечения птиц в 2003-2004 годах развесили 133 синичника (диаметр летка около 3 см), которые располагались на 4 площадках. За период работы утеряно 13 синичников, которые были сняты или разбиты людьми. Две площадки с синичниками находились в окрестностях посёлка Янискоски и две – посёлка Никель. Синичники размещали 2 способами: 1) 14 синичников развесили в ряд («транссектой») длиною в 6 км в окрестностях пос. Янискоски; 2) остальные синичники размещены на площадках. Одна из них (RUS 1; 68º56´ с.ш., 28º45´ в.д.) находилась в 4 км юго-западнее Яниско-ски в бруснично-воронично-багульниково-лишайниковом сосняке с примесью берёзы пушистой. Здесь гнездовья были размещены на площади в 2.85 га в виде двух параллельных линий по 20 гнездовий в каждой. Расстояние между соседними синичниками одной линии было 30 м, между линиями – 50 м. По такой же схеме были заложены еще 2 площадки в окрестностях Никеля. Первая из них (RUS 2; 69º24´ с.ш., 30º06´ в.д.) находилась в 5.4 км северозападнее посёлка, в смешанном сосново-берёзовом лесу с примесью ивы, где напочвенный растительный покров представлен брусникой, вороникой, зелёными мхами, пятнами кустистых лишайников, куртинами злаков и осок. Другая площадка (RUS 3; 69º24´ с.ш., 30º11´ в.д.) располагалась в 2.6 км северо-западнее Никеля вдоль доминирующего направления розы ветров от металлургического комбината. Здесь участки типичного сосново-берёзового леса перемежаются открытыми полянами и участками, пройденными верховыми и низовыми пожарами, зарастающими сосной, берёзой и осиной.
Результаты
На исследуемых территориях в окрестностях заповедника «Пасвик» обитает 5 видов дуплогнездников из воробьиных: мухоловка-пеструшка Ficedula hypoleuca , сероголовая гаичка Parus cinctus , большая синица P. major , пухляк P. montanus и обыкновенная горихвостка Phoenicurus phoenicurus . Искусственные гнездовья почти исключительно заселяли пеструшка (14.8%), сероголовая гаичка (4.7%) и большая синица (3.4%) (табл. 1). За весь период наблюдений установлен только один случай гнездования в синичнике горихвостки. Пухляк вообще не использует искусственные гнездовья. Общая заселяемость синичников птицами за весь период наблюдений возрастала от 9.0 % в первый, 2004 год до 38.3% в третий год наблюдений, в 2006 (табл. 1). Рассмотрим последовательно особенности гнездования каждого из модельных видов.
Таблица 1. Заселённость искусственных гнездовий в заповеднике «Пасвик» в 2004-2006гг.
|
Год |
Кол-во дуплянок |
Количество занятых дуплянок |
|||||||||
|
Птицами |
Другими животными |
||||||||||
|
всего |
в том числе |
всего |
в том числе |
||||||||
|
абс. |
% |
Fh |
Pma |
Pci |
Phoe |
абс. |
% |
оса |
шмель |
||
|
2004 |
133 |
12 |
9.0 |
6 |
2 |
3 |
1 |
— |
— |
— |
— |
|
2005 |
126 |
30 |
23.8 |
17 |
5 |
8 |
— |
2 |
1.6 |
2 |
— |
|
2006 |
120 |
46 |
38.3 |
33 |
6 |
7 |
— |
1 |
0.8 |
— |
1 |
|
Итого |
379 |
88 |
23.2 |
56 |
13 |
18 |
1 |
3 |
0.8 |
2 |
1 |
Fh – Ficedula hypoleuca , Pma – Parus major , Pci – P. cinctus , Phoe – Phoenicurus phoenicurus
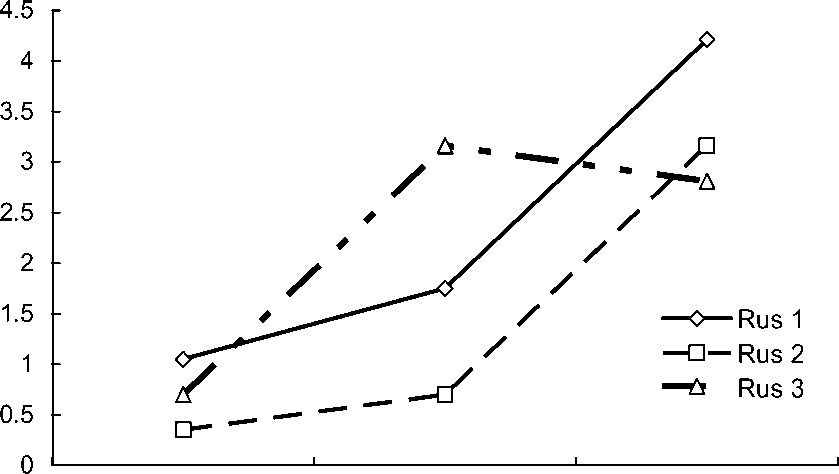
Ficedula hypoleuca – редкий нерегулярно гнездящийся вид сосновых и смешанных лесов заповедника «Пасвик» (Макарова и др. 2003). В 1999-2002 гг. пеструшек не отмечали на учётных маршрутах. В 2003 г был встречен только один поющий самец (Летопись природы… 2005). После размещения в 2004 г. 119 синичников численность F. hypoleuca на этих участках начала возрастать (рис. 1). Сначала плотность гнездования пеструшки на модельной площадке в сосняке была выше (в
2004 – 1.05 , в 2006 – 4.21 пар/га), чем в смешанном лесу (соответственно, 0.35 и 3.16). На площадке в антропогенно-трансформированных лесных сообществах численность этого вида была подвержена колебаниям: в 2004 г. численность составляла 0.7 пар/га, в 2005 г. – 3.16, а в 2006 г. – 2.81 (рис. 1).
2004 2005 2006
Год
Рис. 1. Межгодовая динамика изменения плотности гнездования мухоловки-пеструшки в искусственных гнездовьях.
Rus 1 – площадка, расположенная в сосняке; Rus 2 – в смешанном сосново-берёзовом лесу;
Rus 3 – в трансформированных лесных стациях. Различия по динамике численности птиц в выделенных стациях статистически не значимы (χ2 = 0.25-1.0; df = 2; 9; Р > 0.25-0.75).
Первые яйца в кладках пеструшки появились с 25 мая по 10 июня. В среднем начало кладок приходится на 2 июня ( n = 25). Зарегистрирован единственный случай начала откладки яиц в июле (3 июля 2006) (табл. 2). Птенцы вылуплялись с 14 по 29 июня (в среднем 22 июня), а оставляли гнёзда с 1 по 15 июля (в среднем 8 июля). Величина полной кладки мухоловки-пеструшки в районе исследований составляла от 3 до 8 яиц. Средняя величина кладки 5.85 ± 1.20 яйца (табл. 3). На одну пару в среднем приходилось 4.92 ± 2.10 птенца и 4.70 ± 2.20 слётка. Успешность размножения мухоловки-пеструшки была сравнительно высокой и составляла 79.4% от числа отложенных яиц (табл. 4). Часть погибших яиц была неоплодотворённой или содержала погибшие эмбрионы. Одна кладка была оставлена птицами в период насиживания (табл. 4). Отход птенцов у мухоловки-пеструшки был в 4.5 раза меньше, чем отход яиц. Зафиксирован только один случай гибели птенца в жилом гнезде и один случай оставления птенцов родителями (вероятно, из-за гибели последних).
Таблица 2. Сроки размножения дуплогнездников в лесных сообществах заповедника «Пасвик» в 2003-2006 гг.
|
Вид |
Стадия гнездования |
N |
Май |
Июнь |
Июль |
||||||
|
І |
ІІ |
ІІІ |
І |
ІІ |
ІІІ |
І |
ІІ |
ІІІ |
|||
|
Откладка 1-го яйца |
26 |
– |
– |
9 |
16 |
– |
– |
1 |
– |
– |
|
|
F. hypoleuca |
Вылупление |
28 |
– |
– |
– |
– |
10 |
18 |
– |
– |
|
|
Вылет |
37 |
– |
– |
– |
– |
– |
– |
20 |
17 |
– |
|
|
Откладка 1-го яйца |
10 |
– |
7 |
3 |
– |
– |
– |
– |
– |
– |
|
|
P. cinctus |
Вылупление |
28 |
– |
– |
10 |
15 |
3 |
– |
– |
– |
– |
|
Вылет |
28 |
– |
– |
– |
2 |
9 |
16 |
1 |
– |
– |
|
|
Откладка 1-го яйца |
10 |
– |
– |
7 |
2 |
1 |
– |
– |
– |
– |
|
|
P. major |
Вылупление |
10 |
– |
– |
– |
– |
5 |
4 |
1 |
– |
– |
|
Вылет |
9 |
– |
– |
– |
– |
– |
3 |
5 |
1 |
– |
|
|
Откладка 1-го яйца |
1 |
– |
– |
1 |
– |
– |
– |
– |
– |
– |
|
|
Ph. phoenicurus |
Вылупление |
1 |
– |
– |
– |
– |
– |
1 |
– |
– |
– |
|
Вылет |
1 |
– |
– |
– |
– |
– |
– |
1 |
– |
– |
|
|
P. montanus |
Вылет |
5 |
– |
– |
– |
– |
– |
1 |
3 |
1 |
|
|
Таблица 3. Величина кладок дуплогнездников в лесных сообществах заповедника «Пасвик» в 2003-2006 гг. |
||||||||||
|
Вид |
N |
Количество кладок с числом яиц |
В среднем |
|||||||
|
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|||
|
F. hypoleuca |
39 |
2 |
2 |
11 |
11 |
11 |
2 |
- |
- |
5.85±1.20. m = ±0.19 |
|
P. cinctus |
28 |
- |
- |
3 |
4 |
7 |
11 |
3 |
- |
7.25±1.17. m = ±0.22 |
|
P. major |
10 |
- |
- |
1 |
1 |
3 |
2 |
2 |
1 |
7.60±1.50. m = ±0.47 |
Примечание: В графе «в среднем» указаны показатели: среднее значение ± стандартное отклонение признака, m – ошибка средней.
Таблица 4. Успешность размножения дуплогнездников в лесных сообществах заповедника «Пасвик» в 2003 – 2006 гг.
|
Характеристика |
Ficedula hypoleuca (37 кладок) |
Parus cinctus (28 кладок) |
Parus major (10 кладок) |
|||
|
абс. |
% |
абс. |
% |
абс. |
% |
|
|
Число отложенных яиц |
219 |
100 |
203 |
100 |
76 |
100 |
|
Погибло яиц (всего): |
37 |
16.9 |
17 |
8.4 |
6 |
7.9 |
|
оставление кладки |
16 |
7.3 |
6 |
3.0 |
— |
— |
|
прочие причины |
21 |
9.6 |
11 |
5.4 |
6 |
7.9 |
|
Вылупилось птенцов |
182 |
83.1 |
186 |
91.6 |
70 |
92.1 |
|
Погибло гнездовых птенцов (все- |
8 |
3.7 |
14 |
6.9 |
22 |
28.9 |
|
го): |
7 |
3.2 |
— |
— |
8 |
10.5 |
|
оставлены родителями |
— |
— |
— |
— |
6 |
7.9 |
|
разорение гнезда |
1 |
0.5 |
14 |
6.9 |
8 |
10.5 |
|
прочие причины |
||||||
|
Вылетело птенцов |
174 |
79.4 |
172 |
84.7 |
48 |
63.2 |
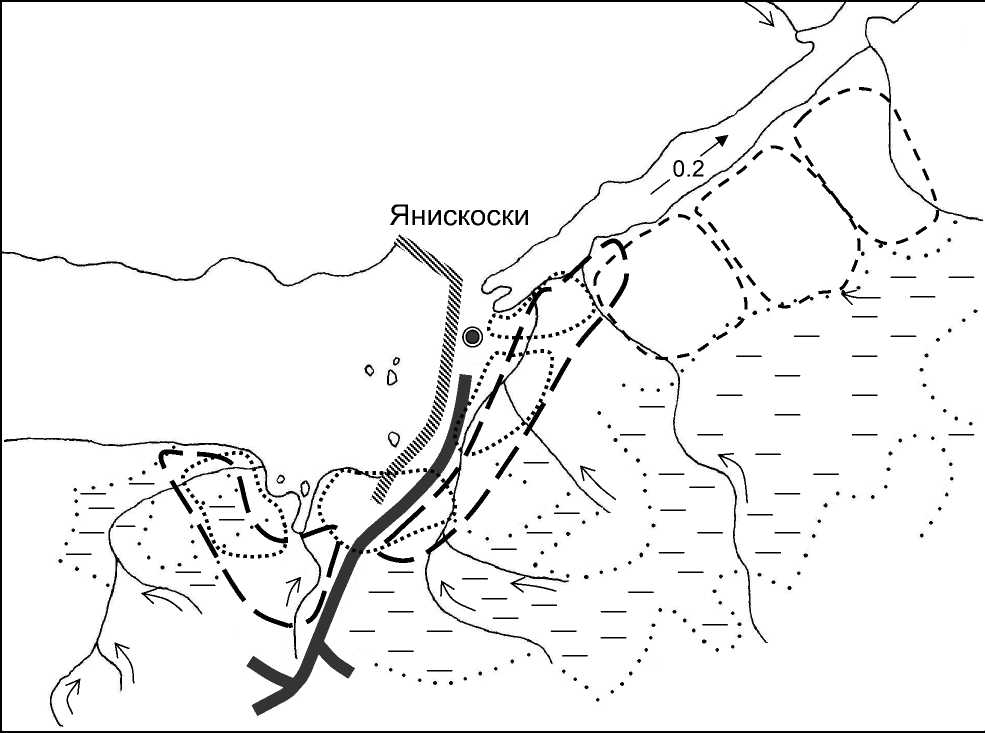
Условные обозначения:
Гнездовые территории:
|
- Parus major |
@ Янискоски – населённые пункты SSSSSSSSS - плотина ГЭС-5 |
|
<'> - Parus cinctus |
^^Н - грунтовые дороги - ручьи |
|
- Parus montanus |
0.2 - реки - болота 300 м |
♦----♦
Рис. 2. Размещение гнездовых территорий синиц в окрестностях заповедника «Пасвик» в 2004 году.
Parus cinctus – наиболее массовый гнездящийся и зимующий вид синиц рассматриваемого района (Макарова и др. 2003). В гнездовой период сероголовые гаички держатся преимущественно в смешанных сосново-берёзовых лесах, растущих на надпойменных террасах. Плотность гнездования в естественных гнездовьях составляет 2.3 пар/км2 (Зацаринный 2005). Площадь гнездовых территорий гаичек в смешанных лесах заповедника «Пасвик» составляет около 43 га (рис. 2). Расстояние между соседними жилыми дуплами было 700-750 м. На модельном участке, где проводили картирование территорий, сосновоберёзовые леса очень однородны по составу. Поэтому территории гаичек имели округлую форму и почти одинаковую площадь.
1.2
0.8
0.6
0.4
0.2
л
/
Rus 1
—□ - Rus 2
—л - Rus 3
----о----
2004 2005 2006 Год
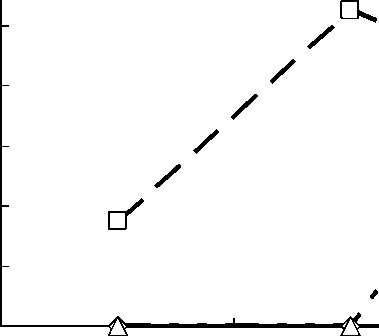
Рис. 3. Межгодовая динамика изменения плотности гнездования сероголовой гаички в искусственных гнездовьях на площадках.
Rus 1 – площадка, расположенная в сосняке, Rus 2 – в смешанном сосно-берёзовом лесу, Rus 3 – в трансформированных лесных стациях.
Сероголовые гаички в основном заселяли синичники, расположенные в смешанных лесах (рис. 3). Ежегодно здесь гнездилось 1-3 пары (0.35-1.05 пар/га). Эти птицы совсем не использовали синичники в чистых сосняках. На площадке в антропогенно-трансформированных лесных стациях (старые пожарища, выбросы металлургического комбината) синицы гнездились только в 2006 году (рис. 2).
Расстояние между соседними жилыми гнёздами в синичниках на трансекте было в среднем 510 м (205-1940 м, n = 6), а между жилыми гнёздами на площадках – 235 м (50-510 м, n = 8).
Первые яйца в гнездах P. cinctus появлялись с 10 по 30 мая. В среднем начало кладок приходилось на 14 мая ( n = 10). Птенцы вылуплялись с 20 мая по 13 июня (в среднем 7 июня) и покидали гнезда с 9 июня по 5 июля (в среднем 23 июня) (табл.2).
Величина полной кладки P. cinctus варьировала от 5 до 9 яиц и в среднем составила 7.25 ± 1.17 яйца (табл. 3). На одну пару в среднем приходилось 6.64 ± 1.74 птенца и 6.14 ± 1.90 слётка. Основные причины отхода были связаны с неоплодотворённостью яиц или гибелью эмбрионов, на что приходится 65% от числа всех погибших яиц ( n = 17). Достоверно установлен только один случай оставления самкой полной кладки. Отход птенцов примерно равен отходу яиц: 6.9% против 8.4% ( n = 203). Успешность размножения гаички была сравнительно высокой – 84.7% от числа отложенных яиц (табл. 4).
Parus major – малочисленный гнездящийся вид заповедника «Пас-вик». Она встречается обычно вблизи населённых пунктов, преимущественно в берёзовых насаждениях (Макарова и др. 2003). Судя по ли- тературным данным, сто лет назад на Кольском полуострове большой синицы не было. Позднее она заселила полосу вдоль железной дороги до побережья Баренцева моря, хотя в восточной, тундровой части полуострова по-прежнему отсутствует (Семенов-Тян-Шанский, Гилязов 1991). В северной Финляндии (69°03´ с.ш., 20°50´ в.д) большую синицу на гнездовании впервые отметили в 1966 году (Valanne et al. 1968 – цит. по: Järvinen 1978). На территории заповедника «Пасвик» и в прилегающих к нему районах большая синица населяет различные типы лесов с преобладанием лиственных деревьев: вдоль дорог, по берегам ручьёв, озёр, рек и болот. Максимальной плотности населения в гнездовое время –0.29 пар/га – она достигает в окрестностях населённых пунктов (рис. 2). Плотность гнездования большой синицы на участках с искусственными гнездовьями была выше. В частности, на площадке в антропогенно-трансформированных лесных стациях она составила в 2004 г. – 0.35, в 2006 – 1.05 пар/га (рис. 4). На площадке в смешанном лесу в 2005-2006 гг. гнездилось только по одной паре P. major, а на модельном участке в сосновом лесу этот вид не встречался (рис. 4).
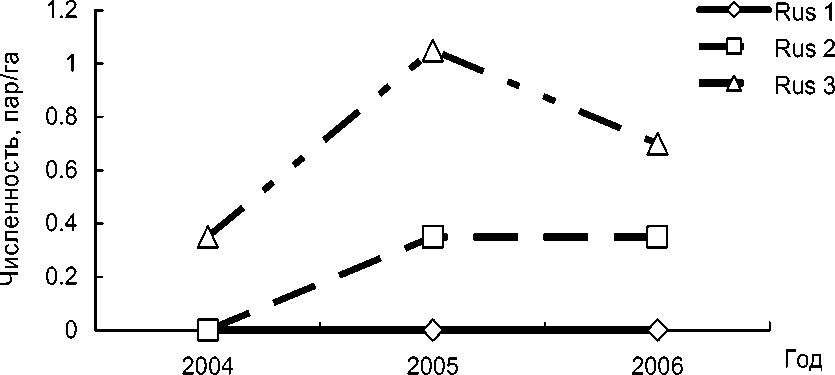
Рис. 4. Межгодовая динамика изменения плотности гнездования большой синицы в искусственных гнездовьях.
Rus 1 – площадка, расположенная в сосняке, Rus 2 – в смешанном сосново-берёзовом лесу, Rus 3 – в трансформированных лесных стациях. Различия по динамике численности птиц в выделенных стациях статистически не значимы ( χ 2 = 0.10, df = 2, Р = 0.95).
Первые яйца в кладках большой синицы появлялись с 21 мая по 3 июня, в среднем 27 мая ( n = 9). Зарегистрирован только один случай откладки яиц во вторую декаду июня (15 июня 2005). Птенцы вылуплялись в разных гнёздах с 11 по 24 июня (в среднем 18 июня), лишь в одном гнезде они появились 10 июля 2005. Оставление гнёзд слётками происходило с 1 по 12 июля (в среднем 5 июля) (табл. 2).
Величина полной кладки варьировала от 5 до 10 яиц и в среднем составила 7.60 ± 1.50 яйца (табл. 2). На одну пару в среднем приходится
7.00 ± 1.56 птенца и 4.80 ± 2.82 слётка. Гибель птенцов превышает отход яиц в 3.6 раза (табл. 4). Зарегистрированы случаи оставления родителями птенцов и один случай разорения гнезда. Успешность размножения составила 63.2% от числа отложенных яиц (табл. 4). Вторые кладки у большой синицы в районе заповедника «Пасвик» нами не отмечены.
Parus montanus – гнездящийся и зимующий вид на территории и в окрестностях заповедника «Пасвик» (Макарова и др. 2003). Пухляк придерживается лиственных и смешанных лесов по долинам рек и крупных озёр. Плотность населения этого вида в период размножения в этих условиях достигала 0.9 пар/км2 (рис. 2). Синичники пухляк обычно не заселяет, как было и в наших условиях. За период работы (2003-2006) было найдено 5 гнёзд пухляка. Все дупла располагались в стволах старых берёз в сосново-берёзовых лесах по долинам ручьёв, впадающих в реку Паз.
Вылет птенцов пухляка регистрировался с 25 июня по 12 июля (в среднем 6 июля) (табл. 2). На одну пару в среднем приходится 5.4 яйца и столько же слётков. Наиболее часто в гнездовой период в районе исследований встречаются пары пухляков без выводков (65%) ( n = 26). Реже отмечали одиночных птиц (15%), которые поют до середины июня, но на их территориях ни гнёзд, ни выводков мы не встречали.
Phoenicurus phoenicurus – обычный гнездящийся вид, населяющий разреженные высокоствольные сосновые и смешанные сосново-берёзовые леса заповедника «Пасвик» (Макарова и др. 2003). В синичниках за весь период наблюдений обнаружено только 1 гнездо горихвостки. Первое яйцо в полной кладке из 7 яиц было отложено 28 мая, 20 июня вылупились 6 птенцов, а 3 июля покинули гнездо лишь 4 слётка.
Обсуждение
На модельных площадках после размещения искусственных гнездовий типа «синичник» стала постепенно возрастать численность мухоловки-пеструшки (рис. 1). Птицы заселяли синичники во всех выделенных нами типах стаций, предпочитая гнездовья, развешенные в сосновых лесах. Здесь за период наблюдений плотность гнездования пеструшек увеличилась с 1.05 (2004 г.) до 4.21 пар/га (2006 г.). Это может свидетельствовать о том, что в хвойных и смешанных лесах северо-запада Кольского полуострова мухоловкам хватает пищи для выкармливания птенцов, но недостаток мест для гнездования ограничивает возможности размножения птиц. Для подтверждения этого мнения требуется дополнительно выяснить, насколько возросла общая численность пеструшек в этом районе. Возможно, что на модельных площадках загнездились птицы с соседних территорий и образовали что-то вроде полуколониального поселения, создающего впечат- ление повышения общей численности птиц. Наши данные подтверждают мнение других исследователей о том, что некоторые виды одиночно гнездящихся птиц в условиях обилия пищи и недостатка мест для гнездования могут гнездиться компактно, образуя своеобразные ассоциации, или полуколонии (Pleasants 1979; Stasey, Bock 1979).
В случае большой синицы развешивание синичников в целом не привело к заметному увеличению численности. Максимальная плотность гнездования этого вида отмечена на площадке в трансформированных человеком лесных стациях. В смешанном лесу этот показатель был ниже (рис. 2). Большие синицы не заселяли гнездовья в сосняках. Как известно, распределение больших синиц по гнездовым биотопам связано с требованиями, предъявляемыми этим видом к кормовым стациям. В районе исследования большие синицы в основном кормились в стациях с преобладанием лиственных деревьев с низкой сомкнутостью крон и наличием открытой (свободной от мхов и лишайников) поверхности земли (Зацаринный 2006), что и отмечено в наших условиях, где подходящих для поисков корма стаций было мало.
У сероголовой гаички и пухляка никаких изменений в плотности населения в гнездовой период на участках с синичниками, по сравнению с участками без них, не отмечено. Вероятно, это связано с тем, что эти виды строго территориальны в разные периоды годового жизненного цикла, и плотность их гнездования не ограничивается наличием подходящих для размножения мест, а определяется запасом пищевых ресурсов на охраняемых территориях и уровнем зимней смертности (Бардин 1975; Cederholm, Ekman 1976; Ekman et al . 1981; Jansson et al . 1981; Карелин 1985; Правосудов 1987; и др.). Биотопическое распределение этих видов в заповеднике связано с выбором этими птицами подходящих кормовых стаций (Зацаринный 2005, 2006).
Подводя итог изложенному выше, можно отметить, что на северной границе распространения сосновых лесов в Фенноскандии не все виды дуплогнездников испытывают недостаток в укрытиях для гнёзд и лимитирующим фактором в большей мере является трофический.
Анализ сроков начала размножения птиц в заповеднике «Пасвик» показал, что мухоловки-пеструшки здесь начинают откладку яиц позже, чем в более южных частях северо-запада России. Так, в заповеднике «Пасвик» пеструшки приступают к размножению в среднем 2 июня (табл. 2), а в Карелии большинство кладок этой мухоловки появляется в конце мая – начале июня, в среднем 31 мая (Зимин 1988; Артемьев 1998). На той же широте, но западнее района наших исследований мухоловки-пеструшки начинают откладку яиц позже, чем в заповеднике «Пасвик» (Järvinen 1983).
Сероголовые гаички в заповеднике «Пасвик» начинают откладку яиц несколько раньше, чем в более южных районах Кольского полу-
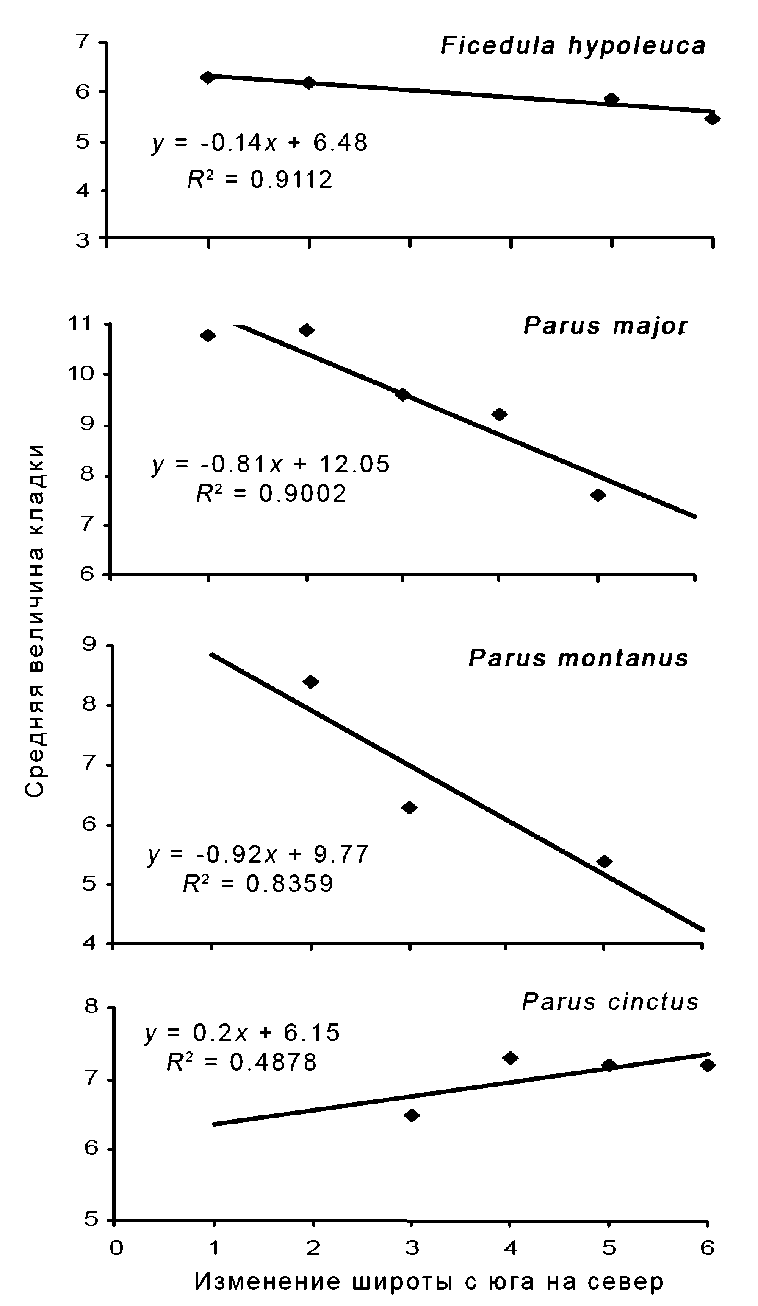
Рис. 5. Географическая изменчивость величины кладки мухоловки-пеструшки, сероголовой гаички, большой синицы и пухляка.
Обозначения: 1 – Окский заповедник (Нумеров 1988), 2 – Карелия (Зимин 1988; Артемьев, 1998), 3 – Кандалакшский заповедник (Правосудов 1987; Шутова 2001), 4 – Лапландский заповедник (Семёнов-Тян-Шанский, Гилязов 1991), 5 – заповедник «Пасвик» (наши данные), 6 – северная Финляндия (Järvinen 1978; 1983).
острова, Финляндии и Норвегии (Järvinen 1978; Правосудов 1987; Се-мёнов-Тян-Шанский, Гилязов 1991).
Большие синицы в заповеднике «Пасвик» начинают откладку яиц немного позднее, чем в южных частях Кольского полуострова (Шутова 2001). Запаздывание сроков начала размножения большой синицы начинается с 64° с.ш., и в заповеднике «Пасвик», по сравнению с южной Карелией, составляет около 10-14 дней (Зимин 1988).
Наши исследования фенологии размножения модельных видов дуплогнездников, полученные на сравнительно небольших выборках, показывают, что сроки начала гнездования у изученных видов птиц в заповеднике «Пасвик», по сравнению с более южными районами, происходит не в строгом соответствии с правилом, согласно которому начало размножения запаздывает на 3-4 сут на каждый градус широты. Это согласуется с мнением других исследователей, что сроки гнездования птиц более тесно связаны с ходом конкретных весенних температур и конкретными экологическими условиями (Järvinen, Linden 1980; Järvinen 1983; Соколов 2006).
Анализ географических особенностей изменчивости величины кладки у птиц-дуплогнездников показал, что не у всех видов, обитающих в заповеднике «Пасвик», средняя величина кладки больше, чем в более южных частях гнездового ареала (рис. 5). У F. hypoleuca , P. major и P. montanus в направлении с юга на север происходит даже уменьшение средней величины кладки.
Так, средняя величина кладки у мухоловки-пеструшки в районе исследований составляет 5.9 яйца ( n = 39). Величина кладки в более западной части ареала, в северной Финляндии, практически не отличается – 5.5 яйца (Järvinen 1983). Южнее, в Карелии, средняя величина кладки немного выше и составляет 6.2 яйца (Артемьев 1998), а в центре европейской части России – 6.3 (Нумеров 1988).
Средняя величина кладки большой синицы в заповеднике «Пас-вик» составляет 7.6 яйца ( n = 10). Этот показатель здесь ниже, чем в более южных частях Кольского полуострова и в Карелии. Так, величина первой кладки большой синицы в Лапландском заповеднике – 9.2, второй – 7.4 яйца (Семёнов-Тян-Шанский, Гилязов, 1991), в Кан-далакшком, соответственно, 9.6 и 8.0 (Шутова 2001). В Карелии средняя величина кладки возрастает до 10.9 яиц (Зимин 1988). Возможно, эти различия обусловлены сравнительно небольшой выборкой.
Величина кладки пухляка в районе исследований составляет 5.4 яйца ( n = 5), что ниже, чем в южной части Кольского полуострова – 6.3 (Правосудов 1987) и Карелии – 8.4 яйца (Зимин 1988).
У сероголовой гаички в европейской части России средняя величина кладки увеличивается с увеличением широты. В заповеднике «Пасвик» (69º с.ш.) она составляет 7.2 яйца (n = 28) и не отличается от более западных районов северной Финляндии – 7.2 (Järvinen 1978). Южнее, в центральной части Кольского полуострова, этот показатель практически такой же – 7.3 (Семенов-Тян-Шанский, Гилязов, 1991). На самом юге Кольского полуострова, на широте Полярного круга, величина кладки гаички ниже, чем в районе наших исследований – 6.5 яйца (Правосудов 1987). Наши данные по сероголовой гаичке согласуются с результатами исследований, проведенных в Якутии, где у этого вида была установлена линейная зависимость увеличения средней величины кладки при переходе с 59-63º до 68º с.ш (Секов, Гермогенов 2006).
Описанные тенденции позволяют заключить, что не у всех перелётных видов птиц-дуплогнездников происходит увеличение средней величины кладки с увеличением широты местности. В частности, это относиться к мухоловке-пеструшке. У оседлых и кочующих птиц, обитающих на северных границах своего распространения и активно расселяющихся далее на север, например, у пухляка и большой синицы, происходит уменьшение средней величины кладки в направлении с юга на север. У оседлых северотаёжных видов, например, у сероголовой гаички, размер кладки увеличивается при переходе от южных районов к более северным. Эти данные свидетельствуют в пользу гипотезы о том, что максимальная величина кладки характерна для оптимальных, а минимальная – для субоптимальных условий в пределах ареала (Пославский 1977; Slagsvold 1981).
Успешность размножения мухоловки-пеструшки и большой синицы в заповеднике «Пасвик» не отличается от этого показателя в более южных районах северо-запада России (Семёнов-Тян-Шанский, Гилязов 1991, Шутова 2001; Артемьев 1998). Успех размножения сероголовой гаички в районе исследования был выше, чем в южных частях Кольского полуострова и в северной Финляндии, но ниже, чем в центральной Якутии (Семёнов-Тян-Шанский, Гилязов 1991; Järvinen 1978; Се-ков, Гермогенов 2006).
Авторы выражают искреннюю благодарность всем сотрудникам заповедника «Пасвик» и экологического центра «Сванховд» (Норвегия), без активного участия и помощи которых выполнение данной работы было бы невозможно. Работа выполнена при финансовой поддержке Государственного природного заповедника «Пасвик», Рязанского государственного университета и экологического центра «Сванховд».
