Особенности репродуктивной биологии жимолости синей Lonicera caerulea L

Автор: Боярских И.Г.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Плодовые и ягодные культуры: физиология, морфология
Статья в выпуске: 1 т.52, 2017 года.
Бесплатный доступ
Жимолость синяя ( Lonicera caerulea L. s. l., сем. Caprifoliaceae ) - высокоценное ягодное растение, культура которого активно развивается в последние годы в странах с умеренным климатом. Ценность этого вида обусловлена сверхранним сроком созревания плодов, высоким содержанием витамина С и биологически активных фенольных соединений, которые широко используются в медицине, косметологии, пищевой промышленности и сельском хозяйстве. Репродуктивная способность сортов и отборных форм L. caerulea сильно варьирует в различных условиях интродукции как в России, так и за рубежом, не обеспечивая реализацию потенциальной урожайности промышленных насаждений. С целью раскрытия возможных причин снижения завязываемости плодов и их массы мы провели оценку морфологической полноценности пыльцы, самоплодности и взаимоопыляемости, а также исследовали проявления несовместимости у сортов и отборных форм этого вида. Исследовали наиболее распространенные в Западной Сибири и на северо-востоке Китая сорта жимолости синей разного эколого-географического и генетического происхождения: L. caerulea subsp. altaica (Pall.) Gladkova (syn. L. altaica Pall.), L. caerulea subsp. venulosa (Maxim.) Worosh. (syn. L. edulis Turcz. ex Freyn), L. caerulea subsp. kamtschatica (Sevast.) Gladkova (syn. L. kamtschatica (Sevast.) Pojark., L. caerulea var. kamtschatica (Sevast.) Pojark.). В полевых опытах в 1999-2002 годах на экспериментальном участке (г. Новосибирск) оценивали завязываемость плодов, их массу, семенную продуктивность, самоплодность и взаимоопыляемость у 8 сортов жимолости (Салют, Берель, Парабельская, Томичка, Памяти Гидзюка, Голубое веретено, Золушка, Камчадалка). Цитоэмбриологические исследования выполняли в 1999-2014 годах на 19 сортах, изучали продукты микроспорогенеза, выявляли нарушения мейоза. Было показано снижение фертильности пыльцевых зерен (до 0,8 %), связанное с аномалиями в мейозе у сортобразцов жимолости алтайского происхождения. Значительное варьирование завязываемости плодов (0,9-64,0 %), их массы и количества семян обусловливалось генетическим контролем реакции несовместимости при автогамии и в различных вариантах перекрестного опыления сортов жимолости синей. В качестве морфологических проявлений реакции несовместимости были выявлены аномалии роста пыльцевых трубок в тканях пестика при переопылении близкородственных сортов и автогамии. Низкое качество пыльцы у отдельных сортообразцов L. caerulea subsp. altaica и частичная несовместимость при перекрестном опылении сортов, вызванная их близкородственным происхождением, могут быть основными причинами снижения продуктивности культурных насаждений жимолости синей. Подбор сортов для совместного выращивания с учетом их генетического происхождения (из разных селекционных семей) позволит обеспечить высокую урожайность насаждений L. caerulea. Для сортов, полученных от родительской формы Старт (Голубое веретено, Синяя птица, Лазурная, Золушка, Герда, Берель), лучшими опылителями могут быть производные исходной формы Дельфин - Томичка, Парабельская, Памяти Гидзюка, Васюганская, Нарымская. При использовании в посадках высокопродуктивных сортов Бархат, Салют, Берель и гибридов с их участием, характеризующихся плохим качеством пыльцы, необходимо включение дополнительных сортов-опылителей.
Стерильность пыльцы, самоплодность, самонесовместимость, взаимоопыляемость сортов, завязываемость плодов, семенная продуктивность, пыльцевые трубки
Короткий адрес: https://sciup.org/142214011
IDR: 142214011 | УДК: 634.74:631.522 | DOI: 10.15389/agrobiology.2017.1.200rus
Features of Lonicera caerulea L. reproductive biology
Blue-berried honeysuckle ( Lonicera caerulea L. s. L.) of the Honeysuckle family ( Caprifoliaceae ) is a high-quality berry plant which has been actively cultivated in recent years in various countries with temperate climate. Blue-berried honeysuckle is valued for ultraearly fruit ripening, as well as high content of antiscorbutic vitamin and biologically active phenolic compounds with antioxdidant, immunomodulating, antibacterial, antiviral, antifungal, antiallergic and other activities widely used in medicine, cosmetic surgery, food industry and agriculture. Variation of reproductive ability of L. caerulea cultivars and selections in different conditions of introduction in Russia and abroad prevents realization of potential fruitfulness of industrial plantations. For the purpose of finding out possible causes of decrease in fruit set and weight, morphological full-value of pollen, self-fertilization and interpollination were assessed, self-incompatibility of this species cultivars was studied. Blue-berried honeysuckle cultivars of various ecological-geographical and genetic origin widespread in West Siberia and north-east China were studied, which belong to the subspecies L. caerulea subsp. altaica (Pall.) Gladkova (syn. L. altaica Pall.), L. caerulea subsp. venulosa (Maxim.) Worosh. (syn. L. edulis Turcz. ex Freyn), L. caerulea subsp. kamtschatica (Sevast.) Gladkova [syn. L. kamtschatica (Sevast.) Pojark., L. caerulea var. kamtschatica (Sevast.) Pojark.]. Self-pollination and interpollination of 8 honeysuckle cultivars (Salyut, Berel', Goluboe vereteno, Zolushka, Tomichka, Parabel'skaya, Pamyati Gidzyuka and Kamchadalka) were studied by data of field research in 1999-2002 on the experimental plot in Novosibirsk. Cytoembryologic research, including that of microsporogenesis products and possible meiosis disturbance of 19 honeysuckle cultivars, was conducted from 1999 to 2014. The results obtained showed a decrease in pollen fertility to 0.8 % connected with anomalies in meiosis. Significant variations in fruit set (0.9-64.0 %), weight and seed quantity were determined by the genetic control of self-incompatibility under self-pollination and in different variants of cross-pollination of blue-berried honeysuckle cultivars. As morphological manifestations of self-incompatibility, anomalies in growth of pollen tubes in style tissues after transpollination of closely related cultivars and autogamy were determined. The main causes of loss cultural plantations productivity of blue honeysuckle may be low pollen fertility in some cultivars L. caerulea subsp. altaica and partial self-incompatibility with the cross-pollination varieties due to their closely related origin. The cultivars for effective pollination of blue-berried honeysuckle plantations were selected (selection from different families). For varieties that are derived from the parent form Start - Goluboe vereteno, Sinyaya ptica, Lazurnaya, Zolushka, Gerda, Berel' best pollinators may be cultivars derived from the initial form of Del'fin - Tomichka, Parabel'skaya, Pamyati Gidzyuka, Vasyuganskaya, Narymskaya. It is necessary to include additional cultivars of pollinators when planting high-yielding varieties Barhat, Salyut, Berel' or new cultivars produced with their involvement which are characterized by low pollen fertility
Текст научной статьи Особенности репродуктивной биологии жимолости синей Lonicera caerulea L
В последние десятилетия в странах с умеренным климатом интенсивно развивается культура жимолости синей (Lonicera caerulea L. s. l., сем. Caprifoliaceae). Ценность этого вида обусловлена сверхранним сроком созревания плодов, высоким содержанием витамина С (1, 2) и биологически активных фенольных соединений (3-5), которые, проявляя антиоксидантную (6-8), иммуномодулирующую (1), антибактериальную (8), противовирусную (1), противогрибковую (2), антиаллергическую и другие виды активности (1), широко используются в медицине (9), косметологии, пищевой промышленности и сельском хозяйстве. В России до- стигнуты серьезные успехи в селекции жимолости синей. В Государственный реестр селекционных достижений РФ, допущенных к использованию, включено более 100 сортов L. caerulea.
В университете Саскачевана (University of Saskatchewan, Канада) сформирована уникальная коллекция жимолости синей, включающая 32 сорта российской селекции, 50 видообразцов из питомников и научных учреждений Северной Америки и России и оригинальный материал дикорастущих форм, собранный в ходе экспедиционных обследований в Японии (о. Хокайдо), Канаде, США. На основе этой коллекции в Канаде создана группа выдающихся сортов, в том числе пригодных для механизированной уборки (10). Товарные плантации жимолости синей ( L. caerulea subsp. emphyllocalyx ) начали закладывать в Японии на острове Хоккайдо с 1970 года (11). В настоящее время собственное производство жимолости в Японии составляет около 200 т/год (85 га), активно ведутся исследования биологии этого подвида (12). Успешная интродукция жимолости синей проводится в США (13). Многочисленные исследования ведутся в европейский странах — Польше (14-17), Чехии, Словакии (9, 18), Румынии (19), Эстонии (20), Литве (21). Достигнуты заметные успехи в промышленном выращивании ягод и их переработке в Китае (22, 23). Для закладки промышленных плантаций жимолости синей в провинции Хей-лунцзян в основном используют сорта российской селекции, а также сеянцы дикорастущих форм: L. boczkarnikowae Plekh. (syn. L. regelyana Boc-zkarn.), произрастающей на северо-востоке Китая (22), и L. caerulea subsp. venulosa (Maxim.) Worosh. (syn. L. edulis Turcz.ex Freyn.) из природных популяций хребта Малый Хинган.
Несмотря на интерес исследователей к L. caerulea , работы по репродуктивной биологии этого вида практически не ведутся. Отсутствие знаний об особенностях биологии опыления и оплодотворения приводит к серьезным ошибкам при реализации программ по широкому введению новой культуры в производство. Обследование питомников в провинции Хейлунцзян, проведенное нами в 2014 году, показало, что в промышленных масштабах размножается только один сорт российской селекции — Берель, выделенный по комплексу хозяйственно ценных признаков в коллекционных насаждениях нескольких хозяйств на северо-востоке Китая. Анализ данных по продуктивности L. caerulea в провинции Хейлунцзян, а также в различных пунктах интродукции в России (23, 24) выявил широкую амплитуду изменчивости по показателям репродуктивной способности сортов и отборных форм жимолости синей.
Исследование биологических особенностей формирования продуктивности у сортов L. caerulea проводилось в 1994-2004 годах в Центральном сибирском ботаническом саду СО РАН (ЦСБС, г. Новосибирск, Россия). Полученные данные частично опубликованы (25, 26), однако круг специалистов, работающих с жимолостью синей в России и за рубежом, продолжает расширяться. Поэтому рассмотрение данных по биологии опыления L. caerulea , полученных ранее и дополненных в последние годы, остается актуальным.
Мы впервые установили, что причинами снижения продуктивности культурных насаждений жимолости синей становятся частичная несовместимость при перекрестном опылении сортов, вызванная их близкородственным происхождением, и низкое качество пыльцы у некоторых сорто-образцов L. caerulea subsp. altaica .
Целью нашей работы было выявление возможных причин снижения потенциальной продуктивности жимолости синей и подбор сортов для совместного выращивания, для чего мы оценивали фертильность мужского гаметофита, самостерильность, взаимоопыляемость, массу плодов и семенную продуктивность, а также изучали характер проявления несовместимости у сортов Lonicera caerulea L. разного географического и генетического происхождения.
Методика. Исследования проводили в Центральном сибирском ботаническом саду СО РАН, в лесостепной зоне юга Западной Сибири, в условиях континентального климата с умеренной обеспеченностью теплом и влагой. Изучали сорта и отборные формы жимолости синей разного эколого-географического и генетического происхождения.
Полевые опыты проводили в 1999-2002 годах. Фертильность пыльцы, завязываемость плодов, их массу и семенную продуктивность оценивали при само- и перекрестном опылении у 8 сортов (Салют, Берель, Парабельская, Томичка, Памяти Гидзюка, Голубое веретено, Золушка, Камчадалка) трех подвидов с тетраплоидным набором хромосом (2 n = 36): L. caerulea subsp. altaica (Pall.) Gladkova (syn. L. altaica Pall.), L. caerulea subsp. venulosa (Maxim.) Worosh. (syn. L. edulis Turcz. ex Freyn), L. caerulea subsp. kamtschatica (Sevast.) Gladkova [syn. L. kamtschatica (Sevast.) Pojark., L. caerulea var. kamtschatica (Sevast.) Pojark.] (27). Контролем служили варианты при естественном свободном опылении. Все варианты опыления исследовали в 3-кратной повторности на разных растениях одного сорта, выбирая одновозрастные ветви.
Жимолость образует компактные соцветия, состоящие из двух тесно сближенных цветков (двуцветник), после опыления которых образуется одно соплодие, поэтому с помощью марлевых мешков изолировали не отдельные цветки, а не менее 100 двуцветников в каждой повторности (под каждым изолятором). Цветки изолировали за 3-5 сут до начала цветения. Опыление выполняли в фазу массового цветения 2-3 раза под каждым изолятором с интервалом 1-2 сут. В период полного созревания учитывали число завязавшихся плодов, среднюю массу одного плода в каждом варианте опыления, число выполненных семян и неразвившихся семязачатков. Результаты учитывали отдельно по каждому изолятору. Пыльцу заготавливали из окрашенных желтых бутонов, подсушивали при рассеянном свете, проверяли фертильность и хранили в эксикаторе в стеклянных пузырьках до опыления (28).
В 1999-2014 годах у 19 сортов жимолости синей оценивали фертильность пыльцы посредством окрашивания ацетокармином (29, 30). Результаты регистрировали с помощью светового микроскопа Primo Star («Carl Zeiss», Германия). Определяли долю нормальных пыльцевых зерен от общего числа изученных.
Лучшими опылителями считались сорта, которые обеспечили долю завязавшихся плодов выше контроля, равный или близкий к нему показатель. Сорта, обеспечившие завязывание 50-70 % плодов по сравнению с контролем, выделялись в группу допустимых опылителей. Сорта, для которых завязывание плодов составляло ниже 50 % к контролю, относили к плохим опылителям (28).
Наблюдения за ростом пыльцевых трубок в тканях пестиков в каждом варианте опыления проводили стандартными методами (32). Столбики пестиков фиксировали через 6, 12, 24, 36 ч после опыления. Препараты прорастающих пыльцевых трубок жимолости окрашивали флуорохромом (анилиновый синий). Препараты анализировали с помощью люминесцентного микроскопа МЛ-2Б с фотонасадкой МФН-10 («ЛОМО», Россия).
Методами математической статистики (31) с применением пакета прикладных программ Microsoft Excel рассчитывали среднюю ( X ) и ошибку средней (S x ).
Результаты. Информация о сортообазцах, изученных в работе, представлена в таблице 1.
1. Географическое и генетическое происхождение сортообразцов Lonicera caeru-lea L., использованных в работе
|
Название |
Происхождение |
Таксон |
|
Голубое веретено |
Сеянцы от свободного опыления отборной формы |
L. caerulea subsp . kamtschatica |
|
Синяя птица Золушка Лазурная Герда Камчадалка |
Старт из камчатской популяции Сеянец от опыления сорта Синяя птица смесью пыльцы сеянца дикорастущей жимолости синей из камчатской популяции Сеянец от свободного опыления отборной формы из камчатской популяции |
2 n = 36 |
|
Салют |
Сеянцы дикорастущей жимолости из Рудного Алтая |
L. caerulea subsp. Altaica |
|
Галочка Сириус Бархат Огненный опал Селена |
(Восточный Казахстан, Лениногорский район) Сеянцы от свободного опыления отборной формы жимолости из Рудного Алтая |
2 n = 36 |
|
Берель |
Сеянец от опыления отборной формы ¹ 12-19 (Сириус) из Рудного Алтая смесью пыльцы сортообразцов Голубое веретено, Синяя птица, Лазурная |
L. caerulea subsp. altaica ½ L. ca-erulea subsp. Kamtschatica 2 n = 36 |
|
Бакчарская Томичка Памяти Гидзюка Нарымская Васюганская Парабельская |
Сеянец от свободного опыления отборной формы ¹ 15-63 из Приморского края Сеянцы от свободного опыления отборной формы ¹ 68-2 (Дельфин) сеянца дикорастущей жимолости из Дальнегорского района Приморского края |
L. caerulea subsp. Venulosa 2 n = 36 |
Успех оплодотворения в значительной степени зависит от жизнеспособности пыльцевых зерен. Анализ фертильности пыльцы (19992014 годы) показал ее высокую морфологическую полноценность у сорто-образцов камчатского и приморского происхождения. При окрашивании ацетокармином фертильность пыльцы у образцов L. caerulea subsp. kam-tschatica в среднем составляла 96,2±1,5 % с коэффициентом вариации ( Сv ) 4,1 %. Для образцов L. caerulea subsp. venulosa фертильность пыльцевых зерен была 91,2±3,4 %, но отмечались более выраженные колебания по годам ( Сv = 9,2 %).
Значительной изменчивостью фертильности пыльцы (от 0,8 до 98,6 %, Сv = 67,2 %) характеризовались сорта алтайского происхождения и гибрид, полученный с их участием (Берель). Большое количество дефектных пыльцевых зерен было обнаружено при исследовании пыльцы у сортов Галочка, Сириус, Бархат, Салют и Берель. Высокое качество пыльцы, не зависевшее от условий выращивания, имели сорта Огненный Опал (91,6-97,5 %) и Селена (87,3-96,6 %). У сортов Берель, Салют, Сириус, Галочка и Бархат также наблюдалась очень слабая продуктивность пыльцы: формировалось низкое число микроспор. У сортообразцов Сириус и Бархат большая часть пыльников не содержала микроспор. Как на цветках, готовых к раскрытию, так и на полностью раскрытых большинство пыльников были зеленого цвета и вялыми.
В наших дополнительных исследованиях (33) у этих образцов были выявлены многочисленные нарушения на разных стадиях мейоза, которые могли становиться причиной низкой фертильности пыльцевых зерен. Снижение фертильности пыльцевых зерен наблюдалось и у гибридов, полученных с участием этих сортов (34). Среди гибридов, у которых сорта Бе-рель и Салют выступали в роли материнских форм, были обнаружены образцы с высокофертильной (до 97 %) и низкофертильной (до 2-37 %) пыльцой. В то же время в гибридных семьях, где низкофертильный сорт выступал в качестве отцовской формы, у всех сеянцев отмечалась высокая фертильность пыльцы. Это свидетельствует о вероятном наследовании стерильности пыльцевых зерен по материнской линии и о необходимости обязательного контроля качества пыльцы при селекционной работе и подборе опылителей для совместного выращивания в производственных насаждениях жимолости синей.
Изучению самонесовместимости у жимолости синей посвящены работы многих исследователей (35-37). Установлено, что образцы этого вида относятся к самостерильным растениям, при их принудительном самоопылении в пределах клона плоды не завязываются вовсе, завязываются плоды с невыполненными семенами с низкой всхожестью или мелкие бессемянные плоды. Самонесовместимость у представителей L. caerulea проявляется в остановке роста пыльцевых трубок в нижней половине столбика пестика (38). Учитывая результаты, приведенные в других работах (39, 40), можно предположить гаметофитный тип реакции несовместимости у L. caerulea . Самонесовместимость прямо влияет на продуктивность, поскольку служит основой генетического контроля полового размножения растений (41, 42). Односортные насаждения, а также неблагоприятные погодные условия в период цветения и созревания плодов, затрудняющие перекрестное опыление с помощью насекомых, обусловливают практически полную бесплодность. В связи с этим необходимо направленно изучать степень самостерильности и при интродукции отбирать формы с наибольшим проявлением самофертильности в генотипе для дальнейшего закрепления этого признака в потомстве с помощью селекции.
Сортообразцы L. сaerulea , изученные нами, были практически са-мобесплодны. После принудительного самоопыления цветков наблюдалось образование 0,9-10,5 % плодов (табл. 2). Наибольшей степенью автофертильности характеризовались сорта Камчадалка, Голубое веретено, Парабельская и Памяти Гидзюка, в отдельные годы при самоопылении завязываемость плодов у этих сортов составляла соответственно 20,0; 15,9; 15,6 и 15,2 % (43). Сорта Салют и Берель, полученные на основе образцов с Алтая, а также сорт камчатского происхождения Золушка, имели низкую самофертильность. У большинства сортов отмечалось формирование небольшого числа плодов после самоопыления. За период от цветения до созревания они немного увеличивались в размерах, достигая 1/31/2 массы плодов, полученных от перекрестного опыления. При наступлении фазы массового созревания плоды приобретали типичную синеголубую окраску. Анализ семенной продуктивности показал, что они были в большей степени бессемянными (табл. 3). Завязавшиеся семена при посеве всходов не давали.
Завязываемость плодов при автогамии и свободном перекрестном опылении колебалась в значительных пределах по годам исследований. Это объясняется отрицательным воздействием на оплодотворение неблагоприятных погодных условий (длительное понижение температуры до 3 ° С, дождь со снегом), которое наблюдалось в период цветения в 2001 и 2002 годах. Наиболее стабильным процентом полезной завязи характеризовались сорта Берель и Памяти Гидзюка.
Изучение взаимоопыляемости форм и сортов L. саеrulea , проведенное ранее в различных научно-исследовательских институтах в России и за рубежом, в большинстве случаев показало успешное переопыление форм, относящихся к подсекции Caeruleae и имеющих одинаковый набор хромосом (27, 36, 37, 44). Скрещиваемость между ди- и тетраплоидными видами жимолости из подсекции Caeruleae возможно, однако завязываемость плодов и выход полноценных семян были очень невелики, а жизнеспособность триплоидных (2 n = 27) гибридов оказалась снижена (12, 27). Следовательно, использование в качестве опылителей образцов с разным числом хромосом недопустимо.
2. Завязываемость плодов на растениях жимолости синей ( Lonicera caerulea L.) разных сортов в зависимости от сорта-опылителя ( X ±S x , г. Новосибирск, 1999-2002 годы)
|
Опыляе- |
Сорт-опылитель |
Контроль |
||||||
|
мый сорт |
1 |
2 3 |
4 |
5 |
6 |
7 |
8 |
|
|
Берель |
0,9±0,76 |
39,6±2,89 24,5±5,34 |
29,6±4,70 |
36,2±3,21 |
29,6±5,25 |
26,7±5,09 |
44,6±1,82 а |
59,0±1,55 |
|
1,5 |
67,1 41,5 |
50,2 |
61,7 |
50,5 |
45,6 |
75,7а |
||
|
Голубое |
23,3±4,05 |
9,3±2,01 28,9±8,26 |
39,0±3,86 |
41,0±7,66 |
39,5±8,46 |
11,4±3,64 |
41,6±3,46 а |
52,9±6,47 |
|
веретено |
43,9 |
17,5 49,2 |
65,7 |
69 |
66,5 |
19,2 |
70,0а |
|
|
Золушка |
27,3±14,45 |
14,0±6,81 3,2±0,71 |
42,6±1,67а |
40,2±7,62а |
32,2±6,02 |
24,4±5,85 |
42,6±5,64а |
54,7±6,80 |
|
49,9 |
25,6 5,8 |
77,9а |
73,5а |
58,8 |
44,6 |
77,8а |
||
|
Камчадалка 13,4±6,10 |
23,6±4,65 20,6±3,02 |
8,1±4,93 |
29,9±3,49 а |
28,0±5,73 а |
18,6±6,45 |
16,8±4,33 |
37,7±7,01 |
|
|
35,6 |
62,7 54,6 |
21,5 |
79,4а |
74,4а |
49,5 |
53,5 |
||
|
Памяти |
24,5±6,29 |
38,0±4,16 а 27,8±4,41 |
26,4±7,20 |
10,5±2,99 |
20,0±3,73 |
12,0±3,91 |
15,8±2,52 |
51,8±2,49 |
|
Гидзюка |
47,3 |
73,5а 53,7 |
51,1 |
20,4 |
38,6 |
23,2 |
30,5 |
|
|
Парабель- |
25,4±3,48 |
42,1±7,48 а 45,2±2,62 а |
40,4±4,03 а |
20,6±4,36 |
8,2±3,26 |
13,3±3,77 |
31,3±3,85 |
54,9±5,46 |
|
ская |
46,3 |
76,7а 82,4а |
73,6а |
37,6 |
14,9 |
24,2 |
57,1 |
|
|
Салют |
16,0±6,40 |
27,2±3,02 30,2±4,28 |
31,7±3,90 |
38,4±6,28 а |
27,1±3,83 |
3,4±1,17 |
31,1±7,39 |
53,9±5,84 |
|
29,7 |
50,4 56 |
58,7 |
71,1а |
50,2 |
6,3 |
57,6 |
||
|
Томичка |
35,1±3,47 |
53,4±3,23 а 38,0±8,44 |
41,1±8,96 |
38,1±6,53 |
25,5±6,52 |
18,3±5,11 |
6,0±1,33 |
64,0±6,86 |
|
54,9 |
83,4а 59,3 |
64,2 |
59,5 |
39,9 |
28,5 |
9,4 |
||
|
Пр имеч ание . 1 — Берель, 2 — Голубое веретено, 3 |
— Золушка, 4 — Камчадалка, 5 |
— Памяти Гидзюка, |
||||||
|
6 — Парабельская, 7 |
— Салют, 8 — Томичка; контроль — свободное опыление. |
Над чертой — сред- |
||||||
|
няя завязываемость плодов, %; под чертой — к контролю, %; |
а — лучшие по совместимости вариан- |
|||||||
|
ты опыления. |
||||||||
В наших исследованиях при искусственном перекрестном опылении изучаемых сортов L. саеrulea доля завязавшихся плодов, их масса и количество семян сильно варьировали в зависимости от сорта-опылителя (см. табл. 3). Низкой завязываемостью плодов характеризовались варианты, где в качестве опылителей использовали сорта Берель и Салют. Отрицательное воздействие на оплодотворение оказывало низкое качество пыльцы. Относительно низкую завязываемость (25,6-48,6 %) в сочетании с небольшой массой плодов и количеством семян имели комбинации опыления, где в качестве родительских форм использовались близкородственные сорта — Голубое веретено ½ Золушка, Золушка ½ Голубое веретено, Памяти Гид-зюка ½ Парабельская, Памяти Гидзюка ½ Томичка, Парабельская ½ Памяти Гидзюка, Томичка ½ Парабельская и Берель ½ Золушка (см. табл. 2).
Предположительно такая частичная совместимость или полусовместимость у жимолости может быть обусловлена гаметофитным контролем реакции несовместимости пыльцы и тканей пестика. В соответствии с гипотезой E.M. East, A.J. Mangelsdorf (45) и молекулярными исследованиями последних лет (42), растения генотипа S1S2 содержат в диплоидных клетках столбика оба аллеля несовместимости, а в гаплоидных пыльцевых зернах при самоопылении либо аллель S1, либо S2. Как пыльца S1, так и пыльца S2 подавляются в тканях столбика, несущего те же факторы. При переопылении растений с гаметофитным контролем несовместимости в пределах семьи, что возможно в вышеперечисленных случаях, вероятно скрещивание гетерозиготы S1S2 ½ S2S3, в результате чего происходит частичное ослабле- ние барьера несовместимости в связи с присутствием среди пыльцевых зерен как несовместимых с тканями пестика S2, так и не вызывающих эту реакцию S3. В зависимости от того, как распределялись S-аллели S-локуса (локуса несовместимости) в потомстве, мы могли наблюдать межсортовую стерильность, частичную или полную совместимость мужского гаметофита и тканей пестика при переопылении близкородственных сортов.
3. Масса плодов и число семян у растений жимолости синей (Lonicera caerulea L.) разных сортов в зависимости от сорта-опылителя (X±Sx, г. Новосибирск, 1999-2002 годы)
|
Опыляемый сорт |
Сорт-опылитель |
|||||||
|
1 |
2 1 |
3 1 |
4 1 |
5 1 |
6 |
7 1 |
8 |
|
|
Берель |
0,35±0,01 0,61±0,01 0,56±0,06 0,60±0,10 0,64±0,03 0,57±0,04 0,45±0,06 0,62±0,03 |
|||||||
|
0 |
12,0±1,1 |
7,1±2,4 |
6,7±3,04 |
8,0±0,2 |
7,7±3,3 |
4,8±0,9 |
8,9±1,2 |
|
|
Голубое вере- |
0,44±0,07 0,37±0,02 0,60±0,10 0,67±0,06 0,73±0,09 0,67±0,03 0,40±0,03 0,73±0,10 |
|||||||
|
тено |
4,1±0,9 |
1,1±0,2 |
4,1±3,2 |
8,7±6,2 |
14,8±0,1 |
6,4±2,2 |
1,4±0,07 |
17,4±4,3 |
|
Золушка |
0,50±0,06 0,50±0,01 0,47±0,08 0,70±0,10 0,76±0,19 0,60±0,10 0,43±0,30 0,80±0,03 |
|||||||
|
1,9±0,5 |
1,8±0,5 |
0,9±0,05 |
5,8±2,1 |
5,9±1,5 |
5,9±1,6 |
1,8±0,5 |
9,9±3,5 |
|
|
Камчадалка |
0,44±0,01 0,70±0,10 0,60±0,10 0,24±0,06 0,68±0,14 0,52±0,08 0,43±0,01 0,60±0,03 |
|||||||
|
5,2±3,7 |
9,5±1,8 |
4,6±1,2 |
0,3±0,3 |
11,9±0,3 |
7,6±1,5 |
4,8±2,4 |
3,3±1,8 |
|
|
Памяти Гид- |
0,50±0,10 0,72±0,09 0,58±0,09 0,46±0,03 0,25±0,07 0,37±0,06 0,32±0,07 0,43±0,10 |
|||||||
|
зюка |
4,7±2,4 |
13,2±3,7 |
8,2±3,0 |
7,3±2,7 |
1,2±1,1 |
4,6±2,7 |
2,4±0,9 |
3,7±1,2 |
|
Парабельская |
0,54±0,06 0,64±0,08 0,78±0,08 0,63±0,05 0,41±0,04 0,31±0,02 0,30±0,02 0,65±0,08 |
|||||||
|
6,9±2,3 |
18,3±1,6 |
15,2±3,7 |
10,4±2,7 |
5,4±2,3 |
1,5±0,6 |
3,2±1,6 |
9,4±2,1 |
|
|
Салют |
0,36±0,08 0,51±0,07 0,49±0,05 0,55±0,09 0,51±0,09 0,42±0,04 0,30±0,04 0,56±0,10 |
|||||||
|
4,0±2,8 |
6,0±1,0 |
5,2±0,3 |
6,9±2,0 |
5,1±1,4 |
3,2±1,4 |
0,8±0,5 |
7,6±0,6 |
|
|
Томичка |
0,42±0,01 0,53±0,04 0,57±0,09 0,54±0,09 0,46±0,09 0,36±0,09 0,31±0,07 0,21±0,04 |
|||||||
|
8,5±0,6 |
13,0±2,1 |
12,3±0,1 |
7,5±3,6 |
10,2±4,6 |
3,7±1,7 |
6,6±5,0 |
0,8±0,8 |
|
Контроль
0,63±0,02
10,2±1,6
0,92±0,03
15,8±1,5
0,84±0,05
9,8±3,1
0,74±0,06
9,7±1,6
0,81±0,05
15,1±0,8
0,67±0,03
13,3±1,6
0,57±0,03
6,6±1,1
0,59±0,01
12,9±2,8
Примечание. 1 — Берель, 2 — Голубое веретено, 3 — Золушка, 4 — Камчадалка, 5 — Памяти Гидзюка, 6 — Парабельская, 7 — Салют, 8 — Томичка; контроль — свободное опыление. Над чертой — средняя завязываемость плодов, %; под чертой — к контролю, %; а — лучшие по совместимости варианты опыления.
А Б В
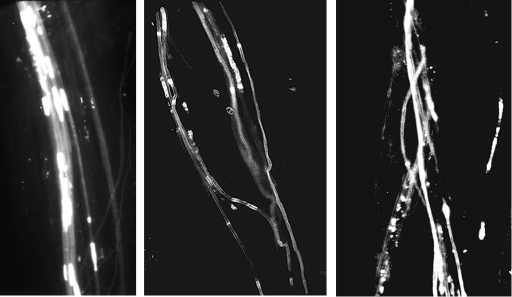
Рост пыльцевых трубок в столбике пестика у растений жимолости синей ( Lonicera caerulea L.): а — при свободном опылении (сорт Золушка; увеличение ½365), б — при опылении близкородственных сортов (Золушка ½ Голубое веретено; увеличение ½277), в — при самоопылении (сорт Томичка; увеличение ½295) (микроскоп МЛ-2Б, «ЛОМО», Россия; г. Новосибирск, 2000 год).
Наблюдения за ростом пыльцевых трубок в тканях пестика показали, что при переопылении сортов, не имеющих общего генетического происхождения, как и при свободном перекрестном опылении, для роста пыльцевых трубок характерна массовость и четкая направленность к семяпочкам. Прямые, ярко флуоресцирующие на фоне тканей пестика пыльцевые трубки росли ближе к центру столбика плотным пучком (рис., А). Каллоз- ные пробки были правильной прямоугольной формы и одинакового размера располагались равномерно по всей длине трубки. В вариантах скрещивания близкородственных сортов так же, как и при самоопылении, наблюдалось отсутствие четкой направленности роста пыльцевых трубок к основанию пестика (см. рис., Б). Отмечено их изгибание, скручивание и слабое свечение уже после прохождения 1/2-2/3 столбика. Каллозные пробки червеобразной или булавовидной формы располагались не регу- лярно и ярко флуоресцировали на едва различимых фоне (в некоторых случаях) пыльцевых трубок. Пыльцевые трубки во всех наблюдаемых вариантах автогамии имели размытое очертание, каллозные пробки были многочисленными и неправильной формы (см. рис. 1, В), что подтверждает данные исследований, проведенных М.Н. Плехановой (38).
Мы распределили сорта по группам (табл. 4). Лучшими опылителями были сорта и отборные формы, которые имели отдаленное генетическое происхождение и характеризовались высокой фертильностью пыльцы. В группу плохих опылителей попали в первую очередь сорта с низкофертильной пыльцой и близкородственные формы.
4. Взаимоопыляемость сортов жимолости синей ( Lonicera caerulea L.) разного эколого-географического происхождения по результатам опытов, проведенных в условиях лесостепной зоны юга Западной Сибири (г. Новосибирск, 19992002 годы)
|
Опыляемые сорта |
Лучшие опылители |
^ Допустимые опылители |
Плохие опылители |
|
Берель |
Томичка |
Голубое веретено Камчадалка Парабельская Памяти Гидзюка |
Салют Золушка |
|
Голубое веретено |
Томичка |
Камчадалка Памяти Гидзюка Парабельская |
Берель Салют Золушка |
|
Золушка |
Памяти Гидзюка Томичка Камчадалка |
Парабельская |
Берель Салют Голубое веретено |
|
Камчадалка |
Памяти Гидзюка Парабельская |
Голубое веретено Золушка Томичка |
Берель Салют |
|
Памяти Гидзюка |
Голубое веретено |
Золушка Камчадалка |
Берель Салют Парабельская Томичка |
|
Парабельская |
Голубое веретено Золушка Камчадалка |
Томичка |
Берель Салют Памяти Гидзюка |
|
Салют |
Памяти Гидзюка |
Голубое веретено Золушка Томичка Камчадалка |
Берель |
|
Томичка |
Голубое веретено Камчадалка |
Золушка Памяти Гидзюка |
Берель Салют Парабельская |
Согласно оценке генетического происхождения, сорта Голубое веретено, Синяя птица, Лазурная, Золушка, Герда, Берель получены от исходной формы Старт ( L. caerulea subsp. kamtschatica ) (43); сорта Томичка, Парабельская, Памяти Гидзюка, Васюганская, Нарымская — производные родительской формы Дельфин ( L. caerulea subsp. venulosa ) (46). Результаты проведенных исследований позволяют предположить более полноценное перекрестное опыление и реализацию потенциальной продуктивности в насаждениях жимолости при использовании в совместных посадках сортов из разных селекционных семей.
Это правило может распространяться и на вновь созданные сорта. Например, в последние годы селекционерами были созданы и получили широкое распространение сорта Бакчарский Великан, Чулымская, Гордость Бакчара, Сильгинка, Бакчарская Юбилейная и др. (46). Поскольку они имеют близкородственное происхождение (получены на основе последующих поколений формы Дельфин), в качестве опылителей для них необходимо использовать сорта из другой семьи.
При закладке плантаций с участием высокопродуктивных сортов Салют, Бархат, Берель и гибридов с их участием, характеризующихся плохим качеством пыльцы, необходимо включение в посадки дополнительных сортов-опылителей.
Таким образом, фертильность пыльцы у сортов жимолости синей ( Lonicera caerulea L.) разного эколого-географического происхождения варьирует в значительных пределах. Низкая фертильность и очень слабая пыльцепродуктивность характерны для ччасти представителей L. caerulea subsp. altaica из Рудного Алтая. Использование в качестве опылителей сортов с высоким содержанием стерильной пыльцы приводит к снижению завязываемости плодов и уменьшению их массы. Все изученные сорта L. сaerulea были самобесплодны. Свободное опыление в условиях лесостепной зоны юга Западной Сибири обеспечило в среднем за годы исследований завязываемость 38-64 % плодов. При скрещивании близкородственных сортов жимолости синей выявлено снижение завязываемости плодов более чем на 50 % по сравнению со свободным опылением, уменьшение массы плодов и семенной продуктивности. В вариантах скрещивания близкородственных сортов жимолости синей и при автогамии наблюдались сходные аномалии роста пыльцевых трубок в тканях пестика, связанные, по-видимому, с гаметофитным контролем самонесовместимости. Подобраны сорта для обеспечения более продуктивного опыления при совместном выращивании.
Список литературы Особенности репродуктивной биологии жимолости синей Lonicera caerulea L
- Svarcova I., Heinrich J., Valentova K. Berry fruits as a source of biologically active compounds: the case of Lonicera caerulea. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub., 2007, 151(2): 163-174 ( ) DOI: 10.5507/bp.2007.031
- Palikova I., Heinrich J., Bednar P., Marhol P., Kren V., Cvak L., Valentova K., Ruzicka F., Hola V., Kolar M., Simanek V., Ulrichova J. Constituents and antimicrobal propeties of blue honeysuckle: A novel sourse for phenolic antioxidants. J. Agric. Food Chem., 2008, 56: 11883-11889 ( ) DOI: 10.1021/jf8026233
- Стрельцина С.А., Сорокин А.А., Плеханова М.Н., Лобанова Е.В. Состав биологически активных фенольных соединений сортов жимолости в условиях северо-западной зоны плодоводства РФ. Аграрная Россия, 2006, 6: 67-72.
- Боярских И.Г., Юшкова Ю.В., Черняк Е.И., Морозов С.В. Содержание биологически активных фенольных соединений в плодах Lonicera caerulea L. различного происхождения в условиях лесостепи Приобья. Вестник Алтайского государственного аграрного университета, 2011, 3: 39-46.
- Боярских И.Г., Васильев В.Г., Кукушкина Т.А. Содержание флавоноидов и гидроксикоричных кислот в Lonicera caerulea (Caprifoliaceae) в популяциях Горного Алтая. Растительные ресурсы, 2014, 1: 105-121.
- Bąkowska-Barczak A.M., Marianchuk M., Kolodziejczyk P. Survey of bioactive components in Western Canadian berries. Can. J. Physiol. Pharmacol., 2007, 85: 1139-1152 ( ) DOI: 10.1139/Y07-102
- Gruia M.I., Oprea E., Gruia I., Negoita V., Farcasanu I.C. The antioxidant response induced by Lonicera caerulaea berry extracts in animals bearing experimental solid tumors. Molecules, 2008, 13(5): 1195-1206 ( ) DOI: 10.3390/molecules13051195
- Celli G.B., Ghanem A., Su Ling Brooks M. Haskap berries (Lonicera caerulea L.) -a critical review of antioxidant capacity and health-related studies for potential value-added products. Food Bioprocess Technol., 2014, 7: 1541-1554 ( ) DOI: 10.1007/s11947-014-1301-2
- Jurikova T., Rop O., Mlcek J., Sochor J., Balla S., Szekeres L., Hegedusova A., Hubalek J., Adam V., Kizek R. Phenolic profile of edible honeysuckle berries (genus Lonicera) and their biological effects. Molecules, 2012, 17: 61-79 ( ) DOI: 10.3390/molecules17010061
- Bors B., Thomson J., Sawchuk E., Reimer P., Sawatzky R., Sander T. Haskap breeding and production final report. ADF 2008-0042, Saskatchewan Agriculture. Regina, 2012.
- Lefol E. Haskap market development -the Japanese opportunity, 2007. Режим доступа: http://www.agriculture.gov.sk.ca/apps/adf/ADFAdminReport/20070012.pdf. Без даты.
- Miyashita T., Hoshino Y. Interploid and intraploid hybridizations to produce polyploid Haskap (Lonicera caerulea var. emphyllocalyx) plants. Euphytica, 2015, 201(1): 15-27 ( ) DOI: 10.1007/s10681-014-1159-4
- Thompson M.M., Chaovanalikit A. Preliminary observation on adaptation and nutracelutical values of blue honeysuckle (Lonicera caerulea) in Oregon, USA. Acta Hortic., 2003, 626: 65-72 ( ) DOI: 10.17660/ActaHortic.2003.626.8
- Dziedzic E. Propagation of blue honeysuckle (Lonicera caerulea var. kamtschatica Pojark.) in in vitro culture. Journal of Fruit and Ornamental Plant Research, 2008, 16: 93-100.
- Małodobry M., Bieniasz M., Dziedzic E. Evaluation of the yield and some components in the fruit of blue honeysuckle (Lonicera caerulea var. edulis Turcz. Freyn.). Folia Horticulturae, 2010 22(1): 45-50 ( ) DOI: 10.2478/fhort-2013-0150
- Smolik M., Ochmian I., Grajkowski J. Genetic variability of Polish and Russian accessions of cultivated blue honeysuckle (Lonicera caerulea). Russian Journal of Genetics, 2010, 46(8): 960-966 ( ) DOI: 10.1134/S1022795410080077
- Ochmian I., Skupien K., Grajkowski J., Smolik M., Ostrowska K. Chemical composition and physical characteristics of fruits of two cultivars of blue honeysuckle (Lonicera caerulea L.) in relation to their degree of maturity and harvest date. Not. Bot. Horti. Agrobo., 2012, 40: 155-162.
- Antalikova M., Matuskovic J. Continuance phonological phase and content of anthocyanins in fruits of the edible honeysuckle (Lonicera kamtschatica Sevast. Pojark.). Proc. Int. Conf. «Perspectives in European Fruit Growing». Lednice, Czech Republic, 2006: 224-225.
- Truta E., Vochita G., Rosu C.M., Zamfirache M.M., Olteanu Z., Oprica L. Karyotype traits in Romanian selections of edible blue honeysuckle. Turk. J. Biol., 2013, 37(1): 60-68 ( ) DOI: 10.3906/biy-1205-28
- Arus L., Kask K. Edible honeysuckle (Lonicera caerulea var. edulis) -under utilized berry crop in Estonia. NJF Report, 2007, 3: 33-36.
- Naugzemys D., Zilinskait S., Skridaila A., Zvingila D. Phylogenetic analysis of the polymorphic 4½ species complex Lonicera caerulea (Caprifoliaceae) using RAPD markers and noncoding chloroplast DNA sequences. Biologia. Section Botany, 2014, 69: 585-593 ( ) DOI: 10.2478/s11756-014-0345-0
- Huo Jun-wei, Yang Guo-hui, Sui Wei, Yu Ze-yuan. Review of study on gemplasm resources of Blue honeysuckle (Lonicera caerulea L). Acta Horticulturae Sinica, 2005, 32(1): 159-164.
- Брыксин Д.М. Ягодоводство в северном Китае. Сады России, 2013, 12: 45-48.
- Плеханова М.Н. Перспективные сорта и формы жимолости для селекции и производства. Труды по прикладной ботанике, генетике и селекции (СПб), 1992, 146: 120-125.
- Боярских И.Г. Результаты эколого-географического испытания сортообразцов Lonicera caerulea. Сибирский вестник сельскохозяйственной науки, 2006, 5: 32-38.
- Боярских И.Г. Особенности морфоструктуры куста жимолости синей в связи с продуктивностью. Сибирский вестник сельскохозяйственной науки, 2007, 9: 66-73.
- Плеханова М.Н. Жимолость (Lonicera subsect. Caeruleae): систематика, биология, селекция. Автореф. докт. дис. СПб, 1994.
- Программа и методика сортоизучения плодовых, ягодных и орехоплодных культур/Под ред. Е.Н. Седова, Т.П. Огольцовой. Орел, 1999.
- Singh R.J. Plant cytogenetics. CRC Press, Boca Raton, 2003.
- Janssen A.W.B., Hermsen J.G.Th. Estimating pollen fertility in Solanum species and haploids. Euphytica, 1976, 25: 577-586 ( ) DOI: 10.1007/BF00041595
- Glantz S.A. Primer of Biostatistics. McGraw-Hill, NY, 2012.
- Delaplanel K.S., Dag A., Danka R.G., Freitas B.M., Garibaldi L.A., Goodwin R.M., Hormaza J.I. Standard methods for pollination research with Apis mellifera. J. Apicult. Res., 2013, 52(4): 1-28 ( ) DOI: 10.3896/IBRA.1.52.4.12
- Боярских И.Г., Куликова А.И. Жизнеспособность пыльцы и мейоз при микроспорогенезе у Lonicera caerulea L. s. l. в условиях лесостепи Приобья. Вестник Алтайского государственного аграрного университета, 2011, 1(75): 39-44.
- Боярских И.Г., Куликова А.И. Особенности микроспорогенеза образцов Lonicera caerulea разного эколого-географического происхождения. Мат. Всерос. конф. с межд. участием «Биологическое разнообразие растительного мира Урала и сопредельных территорий». Екатеринбург, 2012: 193-194.
- Бочкарникова Н.М. Виды и формы жимолости со съедобными плодами, пригодные для культуры. Труды по прикладной ботанике, генетике и селекции (Л.), 1978, 62(2): 72-80.
- Гидзюк И.К. Жимолость со съедобными плодами. Томск, 1981.
- Bożek M. The effect of pollinating insects of two cultivars of Lonicera caerulea L. J. Apicult. Sci., 2012, 56(2): 5-11 ( ) DOI: 10.2478/v10289-012-0018-6
- Плеханова М.Н. Изучение само-и перекрестного опыления жимолости с помощью люминесцентной микроскопии. Бюллетень ВИР, 1982, 126: 53-58.
- Lewis D. Comparative incompatibility in angiosperms and fungi. Adv. Genet., 1954, 6: 235-285.
- Richards A.J. Plant breeding systems. George Allen and Unwin, London, 1986.
- Суриков И.М. Несовместимость и эмбриональная стерильность растений. М., 1991.
- Genetic control of self-incompatibility and reproductive development in flowering plants/E.G. Williams, A.E. Clarke, R.B. Knox (eds.). Springer Science & Business Media, 1994.
- Боярских И.Г. Биологические особенности представителей Lonicera caerulea L. s. l. Автореф. канд. дис. Новосибирск, 2004.
- Жолобова З.П. Селекция голубой жимолости. В сб.: Проблемы устойчивости садовых растений в Сибири. Новосибирск, 1982: 142-145.
- East E.M., Mangelsdorf A.J. A new interpretation of hereditary behavior of self-sterile plants. PNAS USA, 1925, 11: 166-171.
- Савинкова Н.В., Гагаркин А.В. Жимолость на Бакчарском опорном пункте северного садоводства, этапы работы и некоторые итоги. Тр. Межд. науч.-метод. конф. «Состояние и перспективы развития культуры жимолости в современных условиях». Мичуринск, 2009: 129-138.
