Особенности роста брюквенницы Pieris napi (Lepidoptera, Pieridae) в условиях неморальной зоны России
 в условиях неморальной зоны России")
Автор: Нужнова Ольга Камильевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 6 (119), 2011 года.
Бесплатный доступ
Брюквенница, преимагинальные стадии, рост, неморальная зона
Короткий адрес: https://sciup.org/14749962
IDR: 14749962
Текст статьи Особенности роста брюквенницы Pieris napi (Lepidoptera, Pieridae) в условиях неморальной зоны России
Брюквенница (Pieris napi Linnaeus, 1758) - полиморфный вид, представлен достаточно сложным комплексом форм, таксономический статус которых окончательно не определен [8]. Ряд авторов, точку зрения которых мы разделяем, рассматривают P. napi в широком смысле - как надвид, ареал распространения которого на территории России охватывает европейскую часть, Кавказ, Урал, Сибирь, Дальний Восток [2], [6], [7]. В зависимости от физико-географических условий обитания брюквенница имеет моно-вольтинный, бивольтинный или поливольтин-ный жизненный цикл с зимовкой на стадии куколки. Самка откладывает яйца по одному на нижнюю сторону листа или стебель растений из семейств крестоцветные (Brassicaceae Burnett) или резедовые (Resedaceae S. F. Gray); фактическая плодовитость составляет до 450 яиц. Гусеницы появляются через 4-6 дней и развиваются 2-3 недели, в течение которых линяют 4 раза и проходят 5 личиночных стадий. В условиях неморальной зоны жизненный цикл брюк-венницы поливольтинный, лёт имаго наблюдается с марта по октябрь, зимовка куколок длится 4-5 месяцев [2], [7], [13].
Современные исследования, посвященные различным аспектам биологии брюквенницы, проводятся по нескольким направлениям. Одно из них связано с изучением особенностей роста гусениц брюквенницы на разных видах кормовых растений, при этом определяется эффективность питания, темпы роста, продолжительность периода линьки и уровень смертности личинок [2], [13], [17]. В последние годы появились многочисленные публикации по адаптивной значимости монандрии и полиандрии самок брюквен-ницы в различных климатических условиях, влиянию размеров особей обоих полов на репродуктивный успех популяции [9], [10], [15]. Изучается морфологическая изменчивость имаго данного вида, при этом рассматривается ва- риабельность размеров и формы крыльев, меланиновых элементов крылового рисунка [8]. Несмотря на достаточно большое число работ по вопросам биологии и экологии брюквенницы, обширность ареала вида обусловливает неравномерную изученность отдельных популяций и недостаток данных о ходе роста и развития в различных экологических условиях.
Цель исследования - анализ особенностей роста брюквенницы летней генерации в условиях неморальной зоны России.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводили в полевой сезон 2010 года в неморальной зоне России (Карачаево-Черкесская Республика, с. Курджиново, 43°59‘ с. ш., 40°56‘ в. д.) - зоне смешанных и широколиственных лесов с умеренно-теплым климатом. В рассматриваемом районе Карачаево-Черкесии среднегодовая температура воздуха составляет +6,5 °C, осадков выпадает 700-800 мм в год [1], средняя температура июня в 2010 году составила +19,6 °С, июля - +21,4 °С.
Объектом исследования явились особи брюквенницы на разных стадиях жизненного цикла: гусеницы, куколки и имаго. Эксперимент проводился в полевых условиях, особи на всех стадиях развития содержались на разнотравно-злаковом лугу, под кроной дерева при небольшом затенении, на высоте 900 м над уровнем моря. Для проведения исследований были собраны побеги гулявника Лёзеля (Sisymbrium loeseli L.) - одного из кормовых растений гусениц брюквенницы. Данный вид является широко распространенным представителем семейства крестоцветные на территории Европейской России и встречается повсеместно на Кавказе [4]. Побеги поместили в прозрачные пластиковые сосуды объемом 0,5 л с отверстиями в крышке для поступления воздуха. В утренние часы 13.06.2010 провели отлов самок брюквенницы (N = 40) и каждую особь поместили в отдельный сосуд с побегами гулявника. Вечером того же дня бабочек выпустили, а листья с отложенными яйцами собрали. Близкие по габитусу растения гулявника Лёзеля (N = 40) с разнотравно-злакового луга пересадили в прозрачные пластиковые контейнеры с почвой объемом 1 л. На каждое растение перенесли яйца, отложенные одной самкой за день.
В первые четверо суток после появления гу сениц их не взвешивали и не пересаживали, поскольку при этом можно повредить личинок. На 5-й день каждую гусеницу переместили в индивидуальный прозрачный пластиковый сосуд объемом 0,5 л с отверстиями в крышке для поступления воздуха и с указанием порядкового номера. Сосуды содержали свежие побеги гулявника Лёзеля, основание которых было обернуто во влажную марлю. По мере того как листья высыхали или использовались гусеницами в пищу, растения меняли на свежие, причем корм предлагали в избытке, чтобы исключить возможность влияния недостатка пищи на рост личинок [2]. На 5,7, 9 и 11-е сутки с начала личиночной стадии в 12 ч. дня гусениц взвешивали на лабораторных весах ВМ 313 II класса точности.
Сформировавшихся куколок переместили из сосудов в пронумерованные прозрачные пластиковые контейнеры объемом 0,04 л, заполненные мелким гравием. Взвешивание куколок проводили в первые и четвертые сутки после окукливания [13]. Имаго, вышедших из куколок, взвешивали и определяли их пол. Пол гусениц и куколок определен по имаго благодаря использованию непрерывной маркировки сосудов, в которых выращивались особи на всех стадиях жизненного цикла.
Результаты представлены в виде средних величин и стандартных отклонений (M ± S). Смертность особей на разных стадиях жизненного цикла рассчитывали по формуле:
qx
dx nx
где qx - смертность особей, nx - число живых особей в момент учета (x), dx - число особей, погибших в период между двумя последовательными учетами (в интервале от x до x + 1) [3].
Статистическую значимость различий массы тела особей брюквенницы сравнивали с помощью t-критерия Стьюдента после проверки данных на нормальность распределения частот [4]. Зависимость массы гусениц от времени их развития аппроксимировали степенным уравнением регрессии, которое оптимально для описания роста личинок бабочек [14]:
У ! bxa, где у - масса тела, x - день развития гусеницы, a и b - коэффициенты. Для оценки статистической значимости полученных уравнений исполь- зован дисперсионный анализ линейной регрессии. Сравнение аллометрических коэффициентов - показателей степени в уравнениях регрессии для личинок самцов и самок проведено с помощью t-критерия Стьюдента [5].
Для гусениц обоих полов рассчитывали скорость роста по формуле:
m
GR ! Ip, где GR - скорость роста, mp - масса особи при окукливании (г), t - продолжительность развития личинки от момента выхода из яйца до окукливания (сутки) [15]. Средние скорости роста гусениц самцов и самок сравнивали с использованием t-критерия Стьюдента. Корреляционный анализ между показателями массы тела особей брюквен-ницы на разных стадиях жизненного цикла проводили с использованием коэффициента линейной корреляции Пирсона. Количество самцов и самок, вылетевших в определенный день наблюдения, сравнивали по х2-критерию [5].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Эксперимент с момента отлова самок брюк-венницы до вылета имаго следующей генерации продолжался в течение месяца с 13.06 по 13.07.2010. В первый день эксперимента самками брюквенницы (N = 40) отложено 362 яйца (табл. 1). Каждая особь откладывала от 5 до 12 яиц, в среднем 9 ± 2 яйца. Через 6 дней (19.06.2010) из отложенных яиц появились гусеницы, которые развивались 11 суток, за этот период они 4 раза линяли и прошли 5 стадий. При первом подсчете личинок, на 5-е сутки с момента появления, их количество составило 345. Смертность гусениц с 5-х по 7-е сутки достигла 0,041 и к концу личиночной стадии (на 11-е сутки) сократилась до 0,019.
Таблица 1
Изменение количества особей брюквенницы в ходе жизненного цикла в условиях неморальной зоны в 2010 году
|
Стадия и возраст, х |
n x |
d x |
q x |
|
Гусеницы (5-й день) |
345 |
14 |
0,041 |
|
Гусеницы (7-й день) |
331 |
11 |
0,033 |
|
Гусеницы (9-й день) |
320 |
9 |
0,028 |
|
Гусеницы (11-й день) |
311 |
6 |
0,019 |
|
Куколки (1-й день) |
305 |
3 |
0,010 |
|
Куколки (4-й день) |
302 |
1 |
0,003 |
|
Имаго (1-й день) |
301 |
0 |
0 |
Примечание. nx - число живых особей в момент учета (x); dx - число особей, погибших в период между двумя последовательными учетами (в интервале от x до x + 1); qx - смертность особей.
Окукливание гусениц произошло 01.07.2010, всего окуклилось 305 личинок брюквенницы (qx = 0,010), до окончания стадии дожили 302 куколки (qx = 0,003). Стадия куколки у особей обо- их полов продолжалась 8-12 суток. Вылет имаго происходил ежедневно в течение 5 суток (09.0713.07.2010) в утренние часы. Появилась 301 бабочка: 156 особей составили самцы, 145 - самки.
Смертность особей была наиболее высокой в начале личиночной стадии, к стадии куколки она значительно уменьшилась. Минимальная смертность выявлена с момента учета куколок на 4-е сутки их развития до стадии имаго (табл. 1). Причинами гибели особей могли быть бактериальные или вирусные заболевания.
Исследование динамики роста гусениц брюквенницы началось на 5-е сутки с момента их появления. В этот день масса личинок самцов и самок составила 0,005 ± 0,001 г (табл. 2). На 11-е сутки, к окончанию личиночной стадии, масса гусениц самцов превосходила их массу при первичном взвешивании в 36 раз, самок - в 33 раза. Показатели массы личинок брюквенницы в динамике аппроксимировали при помощи уравнения степенной функции. Полученные уравнения регрессии для гусениц обоих полов статистически значимы (табл. 3). Кривые роста личинок самцов и самок (рис. 1, 2) свидетельствуют о том, что для гусениц брюквенницы в условиях неморальной зоны характерен быстрый рост. Сравнение аллометрических коэффициентов в уравнениях выявило, что коэффициент а больше у личинок самцов, чем у самок, при этом различия степенных коэффициентов статистически значимы (t = 2,416; а = 0,016). Таким образом, темпы роста самцов выше, чем самок.
Таблица 2
Динамика массы тела самцов (N = 156) и самок (N = 145) брюквенницы на разных стадиях жизненного цикла в 2010 году
|
Стадия развития |
Пол |
Показатель |
||||||
|
M |
S |
Xmin |
Xmax |
CV |
t |
α |
||
|
Гусеницы (5-е сутки) |
самцы |
0,005 |
0,001 |
0,003 |
0,007 |
14,96 |
1,210 |
0,227 |
|
самки |
0,005 |
0,001 |
0,003 |
0,007 |
14,14 |
|||
|
Гусеницы (7-е сутки) |
самцы |
0,026 |
0,004 |
0,016 |
0,035 |
13,89 |
16,461 |
< 0,001 |
|
самки |
0,021 |
0,002 |
0,013 |
0,027 |
11,75 |
|||
|
Гусеницы (9-е сутки) |
самцы |
0,072 |
0,008 |
0,048 |
0,088 |
11,56 |
10,487 |
< 0,001 |
|
самки |
0,063 |
0,006 |
0,045 |
0,075 |
9,33 |
|||
|
Гусеницы (11-е сутки) |
самцы |
0,178 |
0,011 |
0,149 |
0,207 |
6,13 |
13,520 |
< 0,001 |
|
самки |
0,163 |
0,009 |
0,140 |
0,191 |
5,29 |
|||
|
Куколки (1-е сутки) |
самцы |
0,157 |
0,011 |
0,128 |
0,188 |
6,94 |
11,689 |
< 0,001 |
|
самки |
0,143 |
0,009 |
0,122 |
0,174 |
6,23 |
|||
|
Куколки (4-е сутки) |
самцы |
0,154 |
0,011 |
0,125 |
0,184 |
7,06 |
11,823 |
< 0,001 |
|
самки |
0,140 |
0,009 |
0,119 |
0,165 |
6,26 |
|||
|
Имаго |
самцы |
0,068 |
0,006 |
0,053 |
0,082 |
9,21 |
3,531 |
< 0,001 |
|
самки |
0,066 |
0,006 |
0,051 |
0,080 |
8,93 |
|||
Примечание. M - средняя арифметическая; S - стандартное отклонение; x min - минимальное значение массы тела, г; хтах - максимальное значение массы тела, г; CV - коэффициент вариации, %; t - значения t-критерия Стьюдента; а - уровень значимости.
Таблица 3
Коэффициенты и оценка значимости регрессионных уравнений зависимости массы гусениц от дня их развития в условиях неморальной зоны в 2010 году
|
Пол особей |
a |
b |
R2 |
F |
α |
|
Самцы |
4,4868 |
3,8376 • 10-6 |
0,9899 |
61129,5 |
< 0,001 |
|
Самки |
4,4272 |
3,8304 • 10-6 |
0,9920 |
71785,5 |
< 0,001 |
Примечание. а и b - коэффициенты в степенном уравнении регрессии; R2 - коэффициент детерминации; F - значения критерия Фишера; а - уровень значимости.
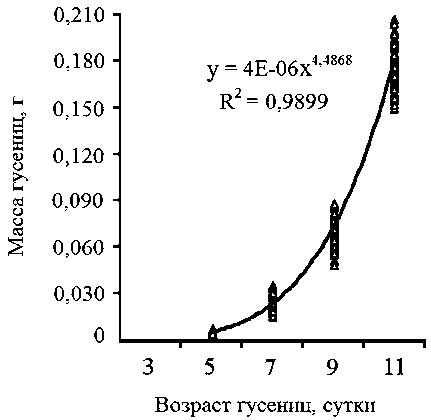
Рис. 1. Динамика массы гусениц самцов брюквенницы в условиях неморальной зоны России в 2010 году
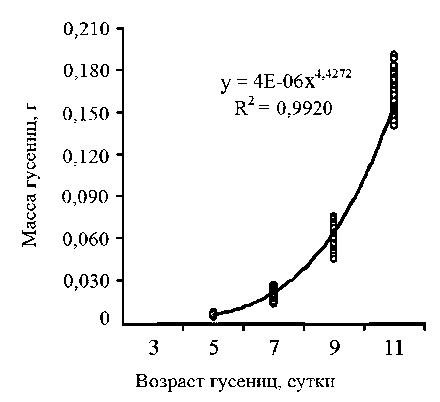
Рис. 2. Динамика массы гусениц самок брюквенницы в условиях неморальной зоны России в 2010 году
Расчет средней скорости роста брюквенницы на личиночной стадии показал, что она достигала 0,014 ± 0,001 г в сутки у гусениц самцов и 0,013 ± 0,001 г в сутки у гусениц самок, данные различия в скорости роста являются статистически значимыми (t = 11,689; а < 0,001).
Масса куколок также различалась у особей обоих полов (табл. 2). Куколки самцов брюквенницы весили на 0,014 г больше, чем куколки самок как в первые, так и на четвертые сутки. Эти различия аналогичны значениям, полученным для конечной массы гусениц, согласно которым личинки самцов весили больше самок на 0,015 г (табл. 2).
Вылет имаго наблюдался в течение 5 дней. Наибольшее количество особей появилось в первые 3 суток, при этом 09.07-10.07.2010 преобладал вылет самцов, в последующие 3 дня - вылет самок, статистическая значимость этих различий подтверждена с использованием х2-крите-рия (рис. 3). Как и на предыдущих стадиях жизненного цикла, у имаго обоих полов выявлены различия массы тела, при этом самцы весили больше самок (табл. 2).
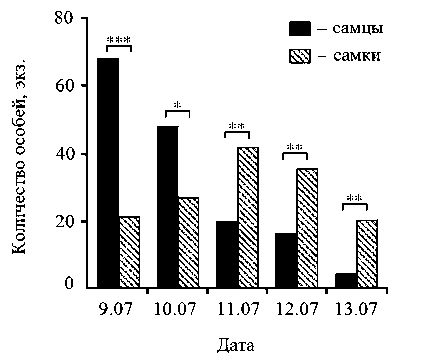
Рис. 3. Динамика появления имаго брюквенницы из куколок в 2010 году: *** - а < 0,001; ** - а < 0,01;
* - а < 0,05 (х2-критерий)
При проведении корреляционного анализа выявлена положительная корреляция массы гусениц 5-го возраста и куколок (г = 0,986; а < 0,001), а также массы куколок и имаго (г = 0,900; а < 0,001). Полученные данные о взаимосвязи веса особей брюквенницы на разных стадиях жизненного цикла согласуются с результатами исследований Ш. Биссундат и К. Виклунд [10], которые также обнаружили корреляцию между массой куколок и имаго.
Таким образом, в ходе эксперимента выявлено, что в условиях неморальной зоны продолжительность развития гусениц самцов и самок летней генерации брюквенницы не различалась и составила 11 суток; стадия куколки у особей обоих полов длилась от 8 до 12 суток; имаго появлялись ежедневно в течение 5 дней. Согласно литературным данным, в бореальной зоне (окрестности г. Стокгольма, Швеция) различия в продолжительности личиночной стадии у самцов и самок брюквенницы варьировали в зависимости от среднесуточной температуры воздуха и составляли менее 12 ч. (при 20 и 23 °С) или более суток (при 17 °С) [17]. Исследователи полага ют, что продолжительность преимагинальных стадий у данного вида в целом и особей каждого пола в частности в значительной степени определяется температурой и фотопериодом. Выявлено, что для летних генераций брюквенницы, развивающихся без диапаузы, характерны незначительные различия в продолжительности личиночной стадии у особей обоих полов, при этом более раннего вылета самцов по сравнению с самками не наблюдается. Напротив, в случае диапаузы у гусениц самок в осенний период развитие менее длительное, чем у гусениц самцов, однако весной на стадии куколки самцы развиваются быстрее и вылетают на 1,5 дня раньше самок [12], [17].
Проведенные исследования позволили установить, что в условиях неморальной зоны средняя скорость роста личинок самцов брюквенни-цы выше, чем самок, на 8 %. По данным Й. Бергстрома, в условиях бореальной зоны скорость роста гусениц обоих полов данного вида различалась на 5 % и при этом также была выше у самцов [9]. Ряд исследователей придерживаются мнения, что именно различия в темпах роста личинок разных полов при одинаковой продолжительности их развития объясняют существование полового диморфизма размеров тела у брюквенницы, а также репницы Pieris rapae (Linnaeus, 1758) и желтушки луговой Colias hyale (Linnaeus, 1758) [11], [12], [16], [17]. Нами выявлено, что в неморальной зоне самцы брюквенницы имеют большую массу тела на стадиях гусеницы, куколки и имаго, чем самки. Рядом авторов получены аналогичные результаты для данного вида на стадии ку кол ки в арктической и бореальной зонах (Финляндия, Швеция) и имаго в бореальной зоне (Швеция) [9], [16], [17]. Брюквен-ница - один из относительно немногих видов, у которого самцы, как правило, крупнее самок. Исследователи считают, что это явление обусловлено полиандрией самок, и объясняют его исходя из теории конкуренции спермы, указывая на связь репродуктивного успеха с размерами самцов брюквенницы [8], [9], [17]. Предполагается, что отбор благоприятствует появлению у данного вида более крупных самцов, поскольку это связано с целым рядом их преимуществ. Так, более крупные самцы при спаривании передают самке больше спермы и питательных веществ, которые используются для развития оплодотворенных яиц, повышают плодовитость и продолжительность жизни самок, увеличивают временной интервал перед повторным спариванием самок и уменьшают количество их спариваний в течение жизни [10], [16].
Исследования динамики массы тела особей на разных стадиях жизненного цикла выявили ее уменьшение в ходе развития «гусеница - куколка - имаго». Потери от конечной массы гусениц составили у куколок 12 % для особей обоих полов, у имаго - 62 % для самцов и 59 % для самок. На стадии имаго самцы весили на 56 %, самки - на 54 % меньше, чем куколки. Потери массы тела у особей обоих полов брюквенницы на разных стадиях жизненного цикла различались не более чем на 3 %. У чешуекрылых потери массы тела в процессе метаморфоза неодинаковы и достигают, например, 46 % при окукливании личинок бражника каролинского Manduca sexta (Linnaeus, 1763) [11]. Вес тела чешуекрылых при метаморфозе уменьшается, поскольку процессы дифференцировки в ходе формирования новых органов и их систем требуют больших энергетических затрат.
ЗАКЛЮЧЕНИЕ
Исследования, проведенные в условиях неморальной зоны России, показали, что длитель- ность стадий развития самцов и самок летней генерации брюквенницы не различается. Наиболее высокая смертность отмечена у гусениц, уменьшается к стадии куколки и снижается до минимальных значений к моменту вылета имаго. Самцы брюквенницы имеют более высокую скорость личиночного роста и бόльшую массу тела на всех изученных стадиях жизненного цикла, чем самки. При метаморфозе происходят потери массы тела, различающиеся у особей обоих полов не более чем на 3 %. Масса и размеры тела ‒ важные факторы репродуктивного успеха самцов из различных популяций брюквенницы, позволяющие оплодотворить большее число яиц у большего количества самок [10].
Список литературы Особенности роста брюквенницы Pieris napi (Lepidoptera, Pieridae) в условиях неморальной зоны России
- Агроклиматический справочник по Ставропольскому краю. Ставрополь, 1958. 234 с.
- Богачева И. А. Факторы, ограничивающие распространение насекомых-филлофагов на Север: случай с брюквенницей и капустницей//Экология. 1997. № 4. С. 293-296.
- Гиляров А. М. Популяционная экология: Учеб. пособие. М.: Изд-во МГУ, 1990. 191 с.
- Дорофеев В. И. Крестоцветные (Cruciferae Juss.) Российского Кавказа//Turczaninowia. 2003. Т. 6. № 3. С. 5-137.
- Ивантер Э. В., Коросов А. В. Введение в количественную биологию. Петрозаводск: Изд-во ПетрГУ, 2003. 304 с.
- Львовский А. Л., Моргун Д. В. Булавоусые чешуекрылые Восточной Европы. М.: Товарищество научных изданий КМК, 2007. 443 с.
- Некрутенко Ю. П. Дневные бабочки Кавказа: Определитель. Семейства Papilionidae, Pieridae, Satyridae, Danaidae. Киев: Наукова думка, 1990. 216 с.
- Шкурихин А. О., Ослина Т. С., Захарова Е. Ю. Анализ изменчивости размеров и формы крыльев Pieris napi L. и P. rapae L. (Lepidoptera: Pieridae) на Среднем и Южном Урале//Экология: от Арктики до Антарктиды: Материалы конф. молодых ученых, 16-20 апреля 2007 г. Екатеринбург: Академкнига, 2007. С. 368-372.
- Bergström J. The evolution of mating rates in Pieris napi. Stockholm: Jannes Snabbtryck Kuvertproffset HB, 2004. 36 p.
- Bissoondath C. J., Wiklund C. Effect of male body size on sperm precedence in the polyandrous butterfl y Pieris napi L. (Lepidoptera: Pieridae)//Behavioral Ecology. 1997. Vol. 8. № 5. P. 518-523.
- Davidowitz G., Nijhout H. F. The physiological basis of reaction norms: the interaction among growth rate, the duration of growth and body size//Integrative and Comparative Biology. 2004. Vol. 44. № 6. P. 443-449.
- Nylin S., Gotthard K. Plasticity in life-history traits//Annual Review of Entomology. 1998. Vol. 43. № 1. P. 63-83.
- Ohata M., Furumoto A., Ohsaki N. Local adaptations of larvae of the butterfl y Pieris napi to physical and physiological traits of two Arabis plants (Cruciferae)//Ecological Research. 2010. Vol. 25. № 1. P. 33-39.
- Tammaru T., Esperk T. Growth allometry of immature insects: larvae do not grow exponentially//Functional Ecology. 2007. Vol. 21. № 6. P. 1099-1105.
- Välimäki P. Reproductive tactics in butterfl ies -the adaptive signifi cance of monandry versus polyandry in Pieris napi. Oulu: Oulun yliopisto, 2007. 66 p.
- Wiklund C., Forsberg J. Sexual size dimorphism in relation to female polygamy and protandry in butterfl ies: A comparative study of Swedish Pieridae and Satyridae//Oikos. 1991. Vol. 60. № 3. P. 373-381.
- Wiklund C., Nylin S., Forsberg J. Sex-related variation in growth rate as a result of selection for large size and protandry in a bivoltine butterfl y (Pieris napi L.)//Oikos. 1991. Vol. 60. № 2. P. 241-250.
