Особенности семенного размножения черники (Vaccinium myrtillus L.) и голубики (V. uliginosum L.) в Пермском крае
 и голубики (V. uliginosum L.) в Пермском крае")
Автор: Данилова М.А., Кетова М.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2018 года.
Бесплатный доступ
Изучены возрастные спектры популяций, семенная продуктивность и всхожесть семян у Vaccinium myrtillus L. и V. uliginosum L. в 6 популяциях на территории Пермского края. Все популяции многочисленные, плодоношение ежегодное, возрастные спектры представлены различными онтогенетическими состояниями, их соотношение указывает на нормальное развитие популяций. Полученные данные показывают, что характеристики семенной продуктивности (количество цветков и плодов на 1 парциальный куст, количество семязачатков в завязи и количество семян в плоде) и всхожести у данных видов сильно вариабельны и в незначительной степени зависят от возрастного спектра популяции. В целом семенное размножение черники и голубики в изученных популяциях ограничено из-за низкой семенной продуктивности и низкой всхожести семян либо продуцирования невсхожих семян.
Семенная продуктивность, всхожесть семян, возрастной спектр, популяция
Короткий адрес: https://sciup.org/147227007
IDR: 147227007 | УДК: 581.9
Seed reproduction features of Vaccinium myrtillus L. and V. uliginosum L. in Perm region
The age spectra of populations, seed productivity and seed germination in Vaccinium myrtillus L. and V. uliginosum L. were studied in six populations in the Perm region. All populations are numerous, fruiting annual, age spectra are represented by different ontogenetic states, their ratio indicates normal development of populations. The data obtained show that the characteristics of seed productivity (number of flowers and fruits per 1 partial bush, number of ovules in the ovary and number of seeds in the fruit) and germination in these species are highly variable and to a small extent depend on the age spectrum of the population. In general, the seed reproduction of V. myrtillus and V. uliginosum in the studied populations is limited due to low seed production and low germination of seeds or the production of non-viable seeds.
Текст научной статьи Особенности семенного размножения черники (Vaccinium myrtillus L.) и голубики (V. uliginosum L.) в Пермском крае
Черника обыкновенная и голубика топяная являются типичными представителями рода Вакциниум ( Vaccinium L.), широко распространенными в Пермском крае [Овеснов, 1997]. У данных видов затруднено семенное размножение; их популяции состоят преимущественно из парциальных образований (кустов и побегов), соединенных корневищами [Серебряков, 1962; Мазуренко, 1982; Шутов, Смирнов, 1991].
Комбинация семенного и вегетативного размножения определяет индивидуальный характер адаптационных способностей каждой популяции. Высокая семенная продуктивность не является гарантией успешного семенного размножения представителей рода Vaccinium из-за непостоянной и в целом невысокой всхожести семян [Лузянина, 2001; Nuortila, 2007; Кутлунина, Зимницкая, Жеребцова, 2008].
У всех представителей рода Vaccinium естественные уровни перекрестного опыления очень высоки, а количество и качество пыльцы не являются лимитирующими факторами, вызывающими снижение реальной семенной продуктивности [Kloet, Lyrene, 1987]. Отмечено 16 видов насекомых, опыляющих чернику, в том числе Andrena lapponica Zitt, Vecpula norvegica (Fab.), Vespula
sylvestris (Scop.), Bombus lapponicus (Fab.), Bombus lucorum (L.), Bombus hortorum (L.) [Ritchie, 1956]. Некоторые виды рода Vaccinium L. проявляют высокую способность к самоопылению, однако семенная продуктивность в этом случае была низкой. Механизмов, препятствующих самоопылению, у черники и брусники не обнаружено [Ritchie, 1956; Nuortila, 2007].
Вегетативное размножение и образование клонов очень распространены для многих видов рода Vaccinium , поэтому даже механизм перекрестного опыления не исключает высокое генетическое сходство обоих родителей. Уровень имбридинга в популяциях брусники, черники, голубики и клюквы может быть очень высок. Исследования эмбриолетальности V. uliginosum L. показали наличие большого количества неразвившихся семяпочек (до 92%) у представителей этого вида [Лузянина, 2001]. Процент развития семян при ксеногамии у V. uliginosum не превышал 45%, у V. myrtillus L. и V. vitis-idaea L. – 17%, что характерно для многих самостерильных растений. Тем не менее этого достаточно для развития полноценных плодов. Для черники и голубики характерна высокая эмбриолетальность (в среднем 55–92%), которая является суммарным результатом аномалий, мегаспорогенеза, опыления, оплодотворения, эмбриогенеза и эн-доспермогенеза. У многих видов данного рода затруднено прорастание семян, что может быть обусловлено различными причинами [Серебряков, 1962; Шутов, Смирнов, 1991; David, Davi, Sean, 2000]. Кроме того, вегетативное размножение ведет к накоплению неблагоприятных мутаций, в том числе тех, которые приводят к стерильности растений, исключая дальнейшее семенное размножение [Кутлу-нина, Зимницкая, Жеребцова, 2008].
Материал и методы исследования
Материал
Было изучено шесть популяций Vaccinium myrtillus L. и V. uliginosum L. на территории Юсь-винского, Краснокамского и Пермского р-нов Пермского края. В Пермском р-не основной древесной породой в месте исследования являлась Pinus sylvestris L., в травяно-кустарничковом ярусе преобладала черника. В Краснокамском р-не был выбран участок заболоченного леса с преобладанием P. sylvestris L. и Betula alba L. В Юсьвин-ском р-не также был выбран участок заболоченного леса с преобладанием Picea fennica (Regel) Kom. и B. alba L. И в Юсьвинском, и в Краснокамском р-нах в травяно-кустарничковом ярусе преобладали черника и голубика. Для различения популяций использованы обозначения: V. myrtillus 1 – популяция черники в Пермском р-не, территория ООПТ «Сосновый бор», V. myrtillus 2 – попу- ляция черники в Пермском р-не, территория ООПТ «Черняевский лес», V. myrtillus 3 – популяция черники в Краснокамском р-не, V. myrtillus 4 – популяция черники в Юсьвинском р-не, V. uligino-sum 5 – популяция голубики в Краснокамском рне, V. uliginosum 6 – популяция голубики в Юсь-винском р-не.
Методы исследования
В каждой популяции закладывались пробные площадки размером 1 м2 регулярным способом в 10-кратной повторности.
Определение онтогенетических состояний парциальных кустов черники и голубики проводилось на основании комплекса качественных морфологических признаков [Полянская, Жукова, Шестакова, 2000].
По количественному составу возрастных групп и их соотношению устанавливали тип популяции. Для оценки возрастной структуры рассчитаны индексы возрастности ∆ [Уранов, 1975] и эффективности ω [Животовский, 2001].
Методика выделения и окрашивания семязачатков модифицирована применительно к специфике объекта. Окрашивание проводилось пропионовым кармином. Завязи нагревали в пропионовом кармине на водяной бане в течение 5–10 мин. Далее из завязей извлекали семязачатки, переносили на предметное стекло и накрывали предметным стеклом [Справочник ..., 2004]. Эта методика позволяет выделить целый семязачаток и определить этап его развития, но не позволяет выявить нарушения в визуально нормально развитых зародышевых мешках, делающих их стерильными.
Семена проращивались в лабораторных условиях. Перед проращиванием семена подвергли стратификации при температуре 4…5ºС в течение 6 месяцев [ГОСТ…, 1998]
Показатели семенной продуктивности определялись по общепринятой методике [Вайнагий, 1974].
Статистическая обработка данных проведена по общепринятой методике в MS EXCEL 2007 c использованием стандартных показателей [Зайцев, 1990].
Результаты и их обсуждение
При изучении популяций V. myrtillus и V. uligi-nosum было обнаружено 7 онтогенетических состояний парциальных кустов: ювенильное (j), взрослое вегетативное (v), молодое генеративное (g1), средневозрастное генеративное (g2), старое генеративное (g3), субсенильное (ss) и сенильное (s). Мы предположили, что семенная продуктивность может быть связана с онтогенетическим состоянием парциального куста, следовательно, показатели семенной продуктивности популяций, обладающих различными онтогенетическими спектрами, будут различны. В популяциях V. uligino- sum наблюдается наличие одновершинных возрастных спектров с максимумом на средневозрастном генеративном состоянии (рис. 1).
Наличие одной вершины возрастного спектра означает, что онтогенез парциальных кустов идет по нормальному пути с последовательным прохождением всех возрастных состояний. В большинстве популяций V. myrtillus также наблюдаются одновершинные онтогенетические спектры. В спектрах популяций черники вершины спектров приходятся на молодое генеративное состояние.
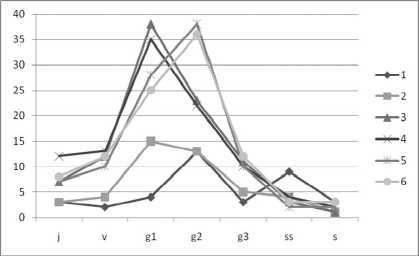
Рис. 1 . Онтогенетические спектры изученных популяций:
1–6 – номера популяций черники и голубики (описание в тексте)
Исключение составляет популяция с территории ООПТ «Сосновый бор», возрастной спектр которой имеет две вершины. При таком распределении более точную типизацию популяций дает классификация «дельта-омега», основанная на соотношении коэффициентов возрастности и эффективности ценопопуляций (табл. 1).
Таблица 1
Возрастная и экологическая характеристика изученных популяций V. myrtillus L. и
-
V. uliginosum L.
Показатель
Популяция
1
2
3
4
5
6
Возрастность популяции, дельта (∆)
0.58
0.43
0.36
0.36
0.40
0.41
Энергетическая эффективность популяции (ω)
0.61
0.61
0.71
0.66
0.75
0.73
Интерпретация коэффициентов согласно классификации «дельта-омега» позволяет отнести популяцию V. myrtillus 1 к стареющим. Популяции V. myrtillus 2 и V. myrtillus 4 относятся к переходным, но из-за достаточно большого количества генеративных особей V. myrtillus 2 популяция приближается к зрелым, а V. myrtillus 4 занимает положение, близкое к зреющим. Популяция V. myrtillus 3 характеризуется переходом от зреющей популяции к зрелой, а популяции V. uliginosum 5 и V. uliginosum 6 относятся к зрелым.
Семенная продуктивность, согласно трактовке И.В. Вайнагий, складывается из следующих показателей:
-
1. Коэффициент плодоцветения (КП) – отношение количества образовавшихся плодов к общему количеству цветков на растении;
-
2. Потенциальная семенная продуктивность (ПСП) – общее количество семяпочек в плоде;
-
3. Реальная семенная продуктивность (РСП) – количество нормально развитых полноценных (выполненных) семян в плоде;
-
4. Процент продуктивности (РСП/ПСП) – отношение реальной семенной продуктивности к потенциальной.
Наименее надежным признаком для описания семенной продуктивности представителей рода Vaccin-ium L. является коэффициент плодоцветения. Значительное количество плодов может быть утрачено на разных этапах созревания из-за механических причин, а также при поедании их животными или сборе ягод людьми. Часть завязей не развивается в плоды. Случайный характер колебаний не позволяет использовать эту характеристику при сравнении популяций между собой. Данные по всем популяциям показывают существенную потерю плодов до фазы созревания (табл. 2). Минимальный коэффициент плодоцве-тения (0.22) отмечен для популяции V. myrtillus 4, максимальный коэффициент плодоцветения выявлен для популяции V.myrtillus 3 (0.53). Для голубики характерны более высокие коэффициенты плодоцветения, чем для черники (0.65 и 0.61).
Таблица 2
Коэффициенты плодоцветения в изученных популяциях V. myrtillus L. и V. uliginosum L.
|
Популяция |
Число цветков на 1 п. к. |
Число плодов на 1 п.к. |
Коэффициент плодоцветения |
|
V. myrtillus 1 |
3.22 ± 0.36 |
1.15 ± 0.54 |
0.36 |
|
V. myrtillus 2 |
4.08 ± 0.53 |
1.44 ± 0.48 |
0.35 |
|
V. myrtillus 3 |
15.33±3.36 |
8.53±2.68 |
0.53 |
|
V. myrtillus 4 |
22.93±4.93 |
5.25±2.72 |
0.22 |
|
V. uliginosum 5 |
23.48±2.70 |
15.18±2.32 |
0.65 |
|
V. uliginosum 6 |
41.88±6.01 |
25.95±4.80 |
0.61 |
Почти все популяции достоверно отличаются по количеству цветков на 1 парциальный куст. Значения t (коэффициент Стъюдента) для популяций V. myrtillus 1 и V. myrtillus 2 – 1.34 (недостоверное значение). Для популяций V. myrtillus 3 и V. myrtillus 4 – 3.11. Для популяций V. uliginosum 5 и V. uligino-sum 6 – 5.63. Кроме того, следует отметить крайне низкие значения этого показателя для популяций черники из рекреационно-парковой зоны г. Перми, которые могут свидетельствовать о неблагоприятном воздействии антропогенной нагрузки на состояние этих популяций.
Потенциальная семенная продуктивность (ПСП) во всех изученных популяциях была высокой (табл. 3).
По количеству семяпочек на 1 завязь популяции V. myrtillus 1 и V. myrtillus 2 демонстрируют значи- тельное сходство. Различие популяций V. myrtillus 3 и V. myrtillus 4 между собой по данному признаку является недостоверным из-за крайне высокой внут-рипопуляционной изменчивости данного признака, о чем можно судить по коэффициенту вариации. Достоверного отличия в ПСП популяций V. uliginosum 5 и V. uliginosum 6 также не обнаружено.
Таблица 3
Потенциальная семенная продуктивность в изученных популяциях V. myrtillus L. и V. uliginosum L.
|
Популяция |
Число семяпочек в 1 завязи |
Коэффициент вариации (cv), % |
|
V. myrtillus 1 |
84.72±15.38 |
17.03 |
|
V. myrtillus 2 |
84.52±13.53 |
24.02 |
|
V. myrtillus 3 |
51.95±8.28 |
49.93 |
|
V. myrtillus 4 |
41.95±9.50 |
70.90 |
|
V. uliginosum 5 |
50.00±5.06 |
31.69 |
|
V. uliginosum 6 |
56.88±6.01 |
33.09 |
Реальная семенная продуктивность (РСП) была значительно ниже ПСП во всех популяциях (табл. 4). Часть семяпочек не развивается в семена; причины этого могут быть различны: аномалии развития зародышевого мешка, отсутствие двойного оплодотворения, механические повреждения и пр.
Таблица 4
Семенная продуктивность в изученных популяциях V. myrtillus L. и V. uliginosum L.
|
Популяция |
Реальная семенная продуктивность |
Коэффициент вариации, % |
Коэффициент продуктивности |
|
V. myrtillus 1 |
44.00±1.90 |
20.75 |
0.52 |
|
V. myrtillus 2 |
36.67±2.87 |
37.50 |
0.43 |
|
V. myrtillus 3 |
29.4±6.35 |
67.62 |
0.52 |
|
V. myrtillus 4 |
22.25±5.49 |
77.24 |
0.40 |
|
V. uliginosum 5 |
34.00±3.22 |
29.67 |
0.70 |
|
V. uliginosum 6 |
41.85±4.30 |
32.15 |
0.73 |
Коэффициент продуктивности наиболее высок в популяциях голубики. Во всех изученных популяциях черники он достаточно стабилен. В целом снижение семенной продуктивности на разных этапах развития во всех популяциях идет стремительно (рис. 2).
Фенотипическая изменчивость всех изученных признаков складывается из двух компонент: средовой изменчивости и генотипической изменчивости или коэффициента наследуемости. Генотипическую изменчивость в природной популяции черники оценить сложно, поскольку для этого потребовалось бы точно определить границы каждого клона. В литературе отмечены случаи отмирания корневища, вследствие чего наблюдалась партикуляция клона, в этом случае точно идентифицировать его границы без молекулярно-генетического анализа невозможно. В пределах своей возрастной группы парциальные кусты морфо- логически сходны между собой, онтогенетическое со- стояние в пределах клона может быть любым.
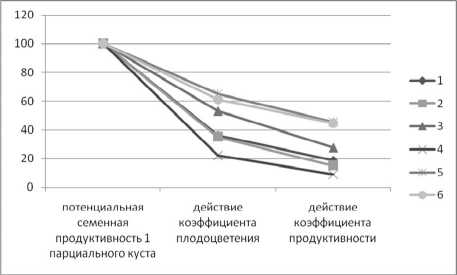
Рис. 2 . Снижение потенциальной семенной продуктивности 1 парциального куста: 1–6 – номера популяций черники и голубики (описание в тексте)
Средовая изменчивость изученных признаков черники может зависеть от множества факторов, среди которых мы выделяем такие как суммарная среднесуточная температура в период вегетации, освещенность, присутствие и активность опылителей, онтогенетическое состояние парциального куста. Собранные нами данные позволяют проследить изменчивость признака в зависимости от онтогенетического состояния.
Был проведен однофакторный дисперсионный анализ выбранных признаков в зависимости от онтогенетического состояния для популяций V. myrtillus L. Нулевая гипотеза заключалась в том, что распределение значений признака является случайным. Анализ проводился при заданном уровне значимости p = 0.05.
Результат анализа подтвердил, что число цветков на 1 парциальный куст в общей выборке демонстрирует зависимость от онтогенетического состояния (F = 2.68) в пределах одной популяции, однако популяции могут достоверно различаться по этому признаку. Результаты изучения зависимости числа плодов на 1 парциальный куст от онтогенетического состояния были недостоверными и исключены из анализа. Число семяпочек на одну завязь не зависит от онтогенетического состояния особей (F = 1.07) в пределах одной популяции и достоверно различается между популяциями. Таким образом, потенциальная семенная продуктивность популяции не зависит от ее возрас-тности, но онтогенетическое состояние необходимо учитывать при определении общего потенциала семенного размножения популяции.
Потенциал семенного размножения не может быть раскрыт без определения всхожести полученных семян. Прорастание семян черники и голубики в природе затруднено, но в лабораторных условиях семена обычно прорастают хорошо [Baskin, Milberg, Andersson, 2000]. Данный признак в изученных популяциях оказался наиболее неоднородным (табл. 5).
Семена из популяций V. myrtillus 1, V. myrtillus 2, V. myrtillus 4 и V. uliginosum 6 были не всхожими. Хранение семян, подготовка их к проращиванию и проращивание проводились в идентичных условиях, таким образом, все отличия связаны исключительно с особенностями изученных популяций и могут свидетельствовать как о аномалиях развития, так и о неблагоприятном воздействии окружающей среды. Популяции V. myrtillus 1 и V. myrtillus 2 испытывали значительную антропогенную нагрузку и были генетически сходны между собой [Данилова, Фатыхова, 2018], но находились на расстоянии друг от друга. Популяции V. myrtillus 4 и V. uliginosum 6 произрастали на одной территории.
Таблица 5
Всхожесть семян в изученных популяциях V. myrtillus L. и V. uliginosum L.
|
Популяция |
Коэффициент всхожести |
Период массового прорастания |
|
V. myrtillus 1 |
0.00 |
- |
|
V. myrtillus 2 |
0.00 |
- |
|
V. myrtillus 3 |
0.69 |
28–32 дни |
|
V. myrtillus 4 |
0.00 |
- |
|
V. uliginosum 5 |
0.22 |
3–5 дни |
|
V. uliginosum 6 |
0.00 |
- |
В тех популяциях, где семена проросли, всхожесть их может быть оценена как средняя ( V. myrtillus 3) и низкая ( V. uliginosum 5).
Выводы
Полученные нами данные показывают, что характер успешности семенного размножения в различных популяций V. myrtillus и V. uliginosum индивидуален. При достаточно высокой потенциальной семенной продуктивности реальная семенная продуктивность значительно ниже. Количество цветков и плодов существенно снижается при интенсивной антропогенной нагрузке. Распространены нарушения всхожести семян, вплоть до их полной нежизнеспособности. Учитывая многократно описанные в литературе проблемы с естественным прорастанием семян представителей рода Vaccinium в природе, можно сделать вывод о том, что семенное размножение часто оказывается неэффективным и доминантное положение в ассоциациях они занимают исключительно благодаря успешному вегетативному размножению.
В популяциях V. myrtillus 1, V. myrtillus 2, V. myrtillus 4 и V. uliginosum 6 семенное размножение отсутствует, несмотря на достаточную семенную продуктивность. Популяция V. uliginosum 5 характеризуется самым низким показателем потенциальной семенной продуктивности (50.00) и низкой всхожестью семян (0.22), ее потенциал семенного размножения тоже невысок. Наиболее успешна популяция V. myrtillus 3.
Список литературы Особенности семенного размножения черники (Vaccinium myrtillus L.) и голубики (V. uliginosum L.) в Пермском крае
- Вайнагий И.В. О методике изучения семенной продуктивности растений // Ботанический журнал. 1974. Т. 59, № 6. С. 826-831
- ГОСТ 13056.6-97 «Семена деревьев и кустарников. Метод определения всхожести» Межгосударственный совет по стандартизации, метрологии и сертификации. Минск: Изд-во стандартов, 1998. 31 с
- Данилова М.А., Фатыхова В.О. Генетическая дифференциация между ценопопуляциями черники (Vaccinium myrtillus L.) в рекреационно-парковой зоне г. Перми // Современные научные исследования и разработки. 2018. № 4(21)-2. C. 169-172
- Животовский Л.А. Онтогенетические состояния, эффективная плотность и классификация популяции растений // Экология. 2001. № 1. С. 3-7
- Зайцев Г.Н. Математика в экспериментальной ботанике. М.: Наука, 1990. 296 с
- Кутлунина Н.А., Зимницкая С.А., Жеребцова М.И. Процессы стерилизации в популяциях вегетативно подвижных растений // Фундаментальные и прикладные проблемы ботаники в начале XXI века: материалы Всерос. конф. Петрозаводск, 2008. Ч. 1. С. 275-277
- Лузянина О.В. Проявление несовместимости у брусничных // Генетические источники лекарственных и ароматических растений. М., 2001. С. 293-297
- Мазная Е.А. Влияние промышленных выбросов на состояние и структуру ценопопуляций Vaccin-ium myrtillus L. и Vaccinium vitis-idaea L. (Кольский п-ов) // Растительные ресурсы. 2001. Т. 37, вып. 3. С. 1-12
- Мазуренко М.Т. Вересковые кустарнички Дальнего Востока (Структура и морфогенез). М.: Наука, 1982. 184 с.
- Овеснов С.А. Конспект флоры Пермской области. Пермь: Изд-во Перм. ун-та, 1997. 252 с
- Полянская Т.А., Жукова Л.А., Шестакова Э.В. Онтогенез черники обыкновенной (Vaccinium myrtillus L.) // Онтогенетический атлас лекарственных растений: учеб. пособие. Йошкар-Ола, 2000. С. 51-60
- Серебряков И.Г. Экологическая морфология растений. Жизненные формы покрытосеменных и хвойных. М.: Высш. шк., 1962. 378 с
- Справочник по ботанической микротехнике. Основы и методы. М.: Изд-во МГУ, 2004. 312 с
- Уранов А.А. Возрастной спектр фитоценопопуля-ций как функция времени и энергетических волновых процессов // Биологические науки. 1975. № 2. С. 7-34
- Шутов В.В., Смирнов А.Н. Опыт анализа возрастного состава побегов ценопопуляций Vaccinium uliginosum L. в Костромской области // Растительные ресурсы. 1991. Т. 27, вып. 1. С. 43-52
- Эмсис И.В. Экспериментальный подход к изучению толерантности травяно-мохового покрова леса к рекреационным нагрузкам // Растительный мир охраняемых территорий. Рига, 1978. С. 150-156
- Baskin C., Milberg P., Andersson L. Germination studies of three dwarf shrubs (Vaccinium, Ericaceae) of Northern Hemisphere coniferous forests // Canadian Journal of Botany. 2000. Vol. 78. P. 1552-1560
- David W., Davi S., Sean S. Studies on the paradox of seedling rarity in Vaccinium myrtillus L. in NE Scotland // Botanical Journal of Scotland. 2000. Vol. 52, № 1. Р. 17-30
- Kloet V., Lyrene M. Self-incompatibility in diploid, tetraploid and hexaploid Vaccinium corymbosum // Canadian Journal of Botany. 1987. Vol. 65. P. 660-665
- Nuortila C. Constraints on sexual reproduction and seed set in Vaccinium and Campanula. Oulu: Oulu University Press, 2007. 60 p
- Ritchie J.C. Biological flora of the British Isles: Vaccinium myrtillus L. // Journal of Ecology. 1956. Vol. 44, № 1. P. 291-299
