Особенности семенной продуктивности Cephalanthera longifolia (L.) Fritsch (Orchidaceae juss.) на трансформированных территориях в Калужской области
 Fritsch (Orchidaceae juss.) на трансформированных территориях в Калужской области")
Автор: Хомутовский Максим Игоревич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 4-4 т.17, 2015 года.
Бесплатный доступ
В статье представлены результаты изучения семенной продуктивности редкого вида Cephalanthera longifolia на северной окраине г. Калуги. Процент плодообразования в ценопопуляции оказался не высоким и в среднем составил 18,3 %, а доля абортивных семян и семян без зародышей в плодах не превысила 3,5 %.
Орхидные, плод, семена, калужская область
Короткий адрес: https://sciup.org/148203935
IDR: 148203935 | УДК: [582.594:577.45]:504.6(470.318)
Features seed efficiency of Cephalanthera longifolia (L.) Fritsch (Orchidaceae juss.) on the disturbed area in the Kaluga region
Article represents the results of studying of seed efficiency of Cephalanthera longifolia in the northern outskirts of Kaluga City (Russia). The percentage of fructification in coenopopulation was not high, average mean was 18,3 %.
Текст научной статьи Особенности семенной продуктивности Cephalanthera longifolia (L.) Fritsch (Orchidaceae juss.) на трансформированных территориях в Калужской области
Работа выполнена при поддержке гранта РФФИ №14-44-03088 р_центр_а,
«Роль техногенных ландшафтов карьеров в сохранении и адвентизации флоры Калужской области».
Cephalanthera longifolia (L.) Fritsch – редкий реликтовый вид с дизъюнктивным ареалом [1]. Он встречается почти по всей Европе (за исключением самых северных районов) от Британских островов и Скандинавии до Средиземноморья, Крыма и Северного Кавказа. Произрастает также в Северной Африке и значительной части Азии. В Восточной Европе за пределами России отмечен в странах Балтии, в Белоруссии, Украине и Молдавии, а также в Закавказье [2]. На территории России находится на северо-восточной границе европейской части ареала и представлен тремя отдельными фрагментами. Наиболее крупный из них находится в Центральной России (Брянская, Калужская, Московская, Орловская, Смоленская, Саратовская, Тверская, Тульская и Курская области), второй – на Северном Кавказе (Республики Адыгея, Дагестан, Кабардино-Балкария, Северная Осетия – Алания, Карачаево-Черкесская, Чеченская и Ингушетия, Ставропольский и Краснодарский края) и третий – на юге Урала (территория Республики Башкортостан, Пермского края и Свердловской области) [1]. Уральские местонахождения вида удалены на 1500 км к востоку от основного ареала. Здесь он растет в светлых березовых (гораздо реже в липовых и хвойношироколиственных) лесах по западному макросклону Среднего и Южного Урала, по склонам речных долин, особенно часто вдоль рек Сылвы и Уфы [3]. В пределах ареала встречается очень редко, популяции обычно очень малочисленны, из многих местонахождений он уже исчез [1].
Вид включен в Приложение II к Международной Конвенции СИТЕС, занесен в Красные книги 18 субъектов РФ [1, 4].
Хомутовский Максим Игоревич, кандидат биологических наук, старший научный сотрудник.
В Тверской области C. longifolia находится на северной границе своего ареала, однако в настоящее время его гербарные сборы с данной территории отсутствуют [5].
Первое упоминание вида на территории Калужской области относится еще к концу XIX в. [6]. В настоящий момент известно несколько точек [7], в трех из них популяции немногочисленны и представлены лишь единичными экземплярами. На северной окраине г. Калуги у дачного массива и ст. Садовой располагается карьер Муратовского щебзавода. Именно здесь были обнаружены наиболее крупные ценопопуляции C. longifolia [8,9]. Вид охраняется на территории национального парка «Угра» [10].
C. longifolia – короткокорневищный травянистый многолетник [11]. На голом, прямом или немного извилистом надземном побеге высотой 25–50 см насчитывается от 5 до 11 линейно-ланцетных листьев 7–16 см длиной и 0,9–3 см шириной. Соцветие прямое, негустое, в котором насчитывается от 3 до 10 (редко до 20) направленных вверх белых, не содержащих нектара, цветков, которые распускаются в конце мая – начале июня [2, 3]. Общая продолжительность онтогенеза составляет 30–40 лет [12]. Вид встречается в широколиственных, реже – смешанных, мелколиственных или хвойных лесах, а также в зарослях кустарников и по опушкам [2]. На Северо-Западном Кавказе растет в сухих тенистых лиственных, очень редко в хвойных лесах, в нижнем горном поясе на склонах южной, юго-западной и юго-восточной экспозиции с крутизной 5-15°. Распространен преимущественно в дубово-грабовых лесах с низким проективным покрытием травянистого яруса (до 30%), но способен поселятся и на вторичных местообитаниях [13]. Предпочитает почвы, бога- тые кальцием [3], изредка растет на глинистых или торфянистых почвах [2].
Вид выращивался в Ботаническом саду Тверского государственного университета [14], однако согласно последней сводке он в настоящий момент отсутствует в коллекции сада [15]. По одним данным C. longifolia сохраняется в культуре лишь короткое время (от 2 до 5 сезонов) [3], по другим – ежегодно цветет и плодоносит, но не размножается [16].
Изучением биологии C. longifolia занимались в Предуралье [17, 18], Крыму [19-22], Прибалтике [23], на Кавказе [12, 13]. Наблюдения также проводят на территории Московской области [24]. Несмотря на это данных, собранных в его естественных условиях, очень мало, тогда как данные о эколого-биологических особенностях C. longifolia на трансформированных территориях и вовсе отсутствуют. В связи с этим целью настоящих исследований стала оценка семенной продуктивности особей C. longifolia, произрастающих на отвалах одного из известняковых карьеров в Калужской области.
МАТЕРИАЛЫ И МЕТОДЫ
Наблюдения и сбор материала проводили летом 2014 г. на отвалах карьера Муратовского щебзавода на северной окраине г. Калуга около ж/д ст. Садовая. В качестве элементарной единицы исследуемого вида принят парциальный побег («условная» особь).
Подсчитывали число цветков (ЧЦ) и плодов (ЧП) на 1 побеге, а также плодозавязывание (ЧП/ ЧЦ×100%). Для определения урожайности подсчет плодов проводили на всех генеративных побегах в популяции. Число семян в плоде определяли, согласно методикам В.В. Назарова [25, 26]. Были вычислены такие показатели: условно-реальная семенная продуктивность (УРСП) – общее число семян в плоде; реальная семенная продуктивность (РСП) – число полноценных семян в плоде; число неполноценных семян, к которым относили семена без зародышей и абортивные семена в плоде. Также определяли РСП в расчете на генеративную побег (РСП плода × среднее число плодов на побеге) и урожай семян, под которым понимали число полноценных семян, продуцируемых растением на единицу площади [27]. В нашем случае мы рассчитывали урожай семян на 1 кв.м (среднее число генеративных побегов на 1 кв.м × РСП генеративного побега). Кроме этого вычисляли общую семенную продуктивность в ценопопуляции (РСП генеративного побега × общее число генеративных побегов). В качестве меры изменчивости признаков использовали коэффициент вариации (CV, %). Уровни варьирования признаков приняты по Г. Н. Зайцеву [28]: СV>20 % – высокий, СV= 11-20 % – средний, СV<10 % – низкий.
Так как C. longifolia является очень редким видом, для изучения семенной продуктивности вида собрали только 30 плодов со зрелыми семенами до начала фазы диссеминации (вскрытия плодов). Перед помещением плода в бумажный пакет измеряли его длину и ширину с помощью штангенциркуля. Затем хранили их в течение двух недель при температуре +24ºС, после чего закладывали в холодильник на постоянное хранение (+4ºС) [29]. Зрелые семена изучали с помощью светового микроскопа MICMED-5, фотоаппарата Canon Power Shot A95 (программы Image J) и сканирующего электронного микроскопа LEO 143OVP (программы LEO SRV-32). Вычисляли среднюю длину и ширину семенной кожуры (тесты) и зародыша, отношение длины к ширине, доля неполноценных семян (ЧНС/УРСП×100%). Объем семенной кожуры и зародыша, соотношение этих показателей и объем воздушного пространства в семени вычисляли по следующим формулам:
VS (объем семени) = 2[(w/2)2×(½l)×(1,047)], где w – ширина семенной кожуры, l – длина семенной кожуры, 1,047 – π/3;
VE (объем зародыша) = 4/3×π×(½l)×(½w)2, где w – ширина зародыша, l – длина зародыша;
AS (объем воздушного пространства в семени) = ( VS-VE )/ VS ×100% [30, 31]. Для определения морфологических параметров брали по 50 зрелых хорошо сформированных семян из каждой коробочки (в целом было измерено 1500 семян). Данные обрабатывали статистическими методами с использованием пакета Microsoft Excel. Тип семян определяли по методике R. Dressler [32].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Ценопопуляция вида произрастает на уплощенном участке вершины отвалов известняка и занимает площадь около 56 кв.м. Отвалы зарастают ивами (преобладала Salix caprea L.), Populus tremula L., Pinus sylvestris L., Hippophaё rhamnoides L. В популяции с общей численностью 175 особей преобладают взрослые вегетативные особи. Плотность в среднем составила 6,2 особей/кв.м. Общее проективное покрытие травянистого яруса, в котором отмечали такие виды как: Fragaria vesca L., Pyrola rotundifolia L., Orthilia secunda (L.) House, Medicago lupulina L., Taraxacum offi cinale Wigg. , Leucanthemum vulgare Lam., Solidago virgaurea L., варьировало от 5 до 30%.
Пыльцеголовник длиннолистный размножается как семенным, так и вегетативным путем. Вегетативное размножение осуществляется двумя путями – делением корневища и образованием растений из придаточных почек на корнях [33]. Роль вегетативного размножения в поддержании популяций пыльцеголовника значительна и его интенсивность зависит от степени микотрофии растений [23].
Тип опыления C. longifolia – исключительно аллогамный [20]. Цветки не содержат нектара и их относят к типу колокольчато-воронковидных [34]. Один из листочков околоцветника – губа выполняет функцию посадочной площадки для насекомых-опылителей. Она разделена перетяжкой на две части: заднюю лопасть – гипохилий, и переднюю – эпихилий, который имеет гребневидные выросты, окрашенные в ярко-желтый цвет, имитирующие пыльники. Вероятно, они выполняют роль дополнительных аттрактантов для пчел [35]. Перетяжка губы является еще одной особенностью цветка Cephalanthera . Ее строение позволяет эпихилию отгибаться вниз под тяжестью тела насекомого, что значительно расширяет круг опылителей за счет крупных пчел, размеры которых превышают размеры внутренней полости цветка, образованной сомкнутыми лепестками и чашелистиками [36]. C. longifolia привлекает неопытных опылителей своими ярко-белыми цветками. Отмечено, что привлекательность цветков безнектарных орхидей для пчел резко увеличивается при наличии в местах их произрастания, так называемых, модельных растений, цветки которых внешне похожи на цветки орхидей, но в отличие от них вознаграждают опылителей пыльцой и нектаром. В Крыму для C. longifolia модельными растениями выступают обильно цветущие в местах его произрастания Polygonatum orientale Desf., P. odoratum (Mill.) Druce, Symphytum tauricum Willd. и Convallaria majalis L. Предполагают, что в опылении принимают участие шмель Bombus haematurus Kriechbaumer и земляная пчела Andrena symphyti Schmied [36]. В Израиле таким модельным растением выступает Cistus salviifolius L. [35].
Плодообразование вида колеблется очень резко – от 0 до 82%, в среднем – 26% [23]. В Крыму процент цветков, из которых завязались плоды, составил 11,0% [20]. В Израиле при совместном произрастании с C. salviifolius плодообразование повышалось до 68% [35]. В данной ценопопуляции оно не превысило 50,00% и составило в среднем всего лишь 18,33% (табл. 1).
Из 55 генеративных особей изученной це-нопопуляции только на 41 сформировались плоды. Из них по одному плоду завязалось на 36% особей, по два – у 27%. Максимальное число плодов (7) было отмечено только у одной особи. Если рассматривать положение их на побеге, то у большинства плоды завязались только из цветков, располагавшихся в нижней части соцветия, лишь у 20% особей плоды были отмечены в его срединной части. На вершине побега плоды не завязались практически ни у кого. Исключение составила, только одна особь с 6 плодами, у которой они были отмечены как в нижней и срединной, так и в верхней части соцветия.
Предполагают, что уровень плодоношения сильно зависит от освещенности местообитаний. Так, отмечено, что C. longifolia в светлых мелколиственных лесах Среднего Урала вызревшие плоды дают от 21,6% до 37,4% цветков, а под пологом тенистых липовых и темнохвойно-липовых лесов – лишь 1,9–4,2% [3]. В Предуралье в смешанном лесу с относительной освещенностью более 10% плодообразование составило 16,1%, а в местообитании, где освещенность достигала 60–80%, оно увеличивалось до 26,4% [18]. Большинство особей изученной ценопопуляции произрастали в световом окне, часть под невысокими соснами и относительная освещенность варьировала от 40% до 95%. Кроме того, генеративные побеги, на которых не завязались плоды были отмечены как на участках с полной освещенностью, так и с затенением. А большинство особей, на которых было отмечено по 3–4 плода, располагались в условиях некоторого затенения. На наш взгляд снижение плодообразования связано не столько с уровнем освещенности местообитания, сколько с численностью опылителей в нем, а также наличием модельных (кормовых) растений.
Плод у C. longifolia – сухая коробочка, которая при созревании вскрывается 6 продольными щелями и имеет обратноузкояйцевидную форму. Их длина у изученных особей варьировала от 21 до 33 мм и составила в среднем 28,40±0,66 мм. Ширина плода находилась в диапазоне от 5 до 7 мм, а среднее значение составило 6,34±0,11 мм. Следов повреждения плодов насекомыми-фитофагами не отмечалось. Процент неполноценных
Таблица 1. Характеристика плодов и семенная продуктивность Cephalanthera longifolia
|
Показатель |
min-max |
M±m |
CV, % |
|
Число цветков на 1 генеративном побеге |
3–21 |
8,84± 0,54 |
45,57 |
|
Число плодов на 1 генеративном побеге |
0–7 |
1,75± 0,23 |
97,51 |
|
Плодообразование, % |
0,00–50,00 |
18,33±2,01 |
81, 32 |
|
Длина плода (L f ), мм |
21–33 |
28,40±0,66 |
12,77 |
|
Ширина плода (W F ), мм |
5–7 |
6,34±0,11 |
9,13 |
|
L F / W F |
3,83–5,64 |
4,48 |
9,24 |
|
УРСП плода, шт |
3052–11921 |
6681,33±450,67 |
36,95 |
|
РСП плода, шт |
3025–11773 |
6606,37±444,96 |
36,89 |
Примечание к табл. 1 : min-max – минимальное и максимальное значение; M – среднее арифметическое значение, m – стандартная ошибка среднего арифметического значения, CV – коэффициент вариации
семян в плодах варьировал незначительно – от 0,35% до 3,26% и в среднем составил 1,12±0,11%. В Пермском крае этот показатель оказался также низким – 0,20% [17].
Исследования в Крыму показали, что число семязачатков в завязи у C. longifolia , которое отражает потенциальную семенную продуктивность, варьировало от 3521 до 12793 шт., тогда как число полноценных семян (РСП), сформировавшихся в плоде, в среднем оказалось невысоким – 5993,0 шт. [20]. В Предуралье этот показатель находился в диапазоне от 3361 до 6261 шт., а среднее значение составило 4283,2 шт. [17]. В изученных нами плодах значения РСП были в более широких пределах и в среднем показатель оказался выше, чем в Крыму и Предуралье (табл. 1). РСП генеративной особи составила 11531 шт., что почти в два раза выше, чем было отмечено в Крыму, где этот показатель составил – 6592 шт. [20]. Это связано с тем, что, кроме невысокой РСП плода, в период наблюдений в среднем число вызревших плодов на побеге оказалось также ниже (1,1±0,30 шт.) [20], чем в изученной нами ценопопуляции (табл. 1).
Число генеративных особей в период наблюдений в среднем было 2,8 шт./кв.м. В связи с этим урожай семян составил 32287 шт./кв.м. Общая же семенная продуктивность в ценопопуляции оказалась – 634212 шт.
Основная причина снижения семенной продуктивности у C. longifolia связана с системой скрещивания (исключительно аллогамной) вида, характеризующейся факультативной мимикрией. Отсутствие в местообитании C. longifolia модель-но (кормового) вида существенно снижает шансы успешного опыления орхидеи. Также отмечено, что у C. longifolia по сравнению с другими видами рода Cephalanthera (C. rubra (L.) Rich., C. damasonium (Mill.) Druce) отмечен наименьший вклад пластических веществ в органы репродукции и всю надземную часть [22]. Вид, вероятно, выработал стратегию экономии ресурсов. Так у раскрывшихся цветков C. longifolia плацента в завязи дифференцирована лишь на небольшие меристематические бугорки, а семязачатки формируются только через несколько недель после опыления. Это позволяет виду не тратить пластические вещества на производство семязачатков, которые в последствии не участвуют в образовании семян. Кроме того, затраты на формирование неопыленных цветков частично компенсируются за счет фотосинтеза. Производство семян также не только не сопровождается ослаблением особи, а, наоборот, большинство особей увеличивают свою фитомассу и неплодоносящие или слабо плодоносящие особи цветут регулярно в течении многих лет [21]. Снижение семенной продуктивности у вида идет на уровне особи. Об этом говорят данные, полученные в Крыму. Так коэффициент продуктивности особи у C. longifolia составил всего 0,088, тогда как коэффициент продуктивности цветка оказался достаточно высоким – 0,768 [20].
Согласно типификации семян орхидных [32] семена C. longifolia были отнесены к Limodorum-типу и имели веретеновидную или обратноузкояйцевидную форму. Однослойная семенная кожура (теста) оказалась почти прозрачной и сквозь нее хорошо виден зародыш овальной формы, имеющий насыщенную темно-коричневую окраску. Размеры тесты полноценных семян составили в среднем 1052,83±13,10 мкм в длину и 199,90±1,99 мкм в ширину (табл. 2). Таким образом, семена C. longifolia в Калужской области оказались немного короче и шире, чем в Пермском крае, где длина семени в среднем была 1,40±0,04 мм, а ширина – 0,18±0,01 мм [17]. Теста семян из Крыма также имела большие размеры по сравнению с изученными нами: ее длина в среднем была 1,312±0,042 мм, а ширина – 0,203±0,005 мм [20]. Зародыш, как и у всех орхидных, не дифференцирован, его длина варьировала от 162,96 мкм до 238,54 мкм и составила в среднем 201,13±1,62 мкм, а ширина – от 108,33 мкм до 160,76 мкм (126,42±1,06 мкм в среднем). Эти данные в целом совпадают с данными, полученными В.В. Назаровым [20] в Крыму, где длина и ширина зрелого зародыша семени составили 0,204±0,006 мм и 0,131±0,003 мм соответственно. Абортивные семена, как правило, имели неправильную форму и меньшие размеры (рис.).
Благодаря своим небольшим размерам семена хорошо разносятся ветром. Легкость им придает полость, заполненная воздухом. Она образуется в результате разрушения внутреннего интегумента, нуцеллуса и эндосперма [37-40]. Объем воздушного пространства варьировал от 73,0% до 91,2% и в среднем оказался достаточно высоким (табл. 2).
Из изученных показателей C. longifolia наиболее изменчивыми оказались показатели семенной продуктивности (табл. 1), а наименее – объем воздушного пространства в семени и размеры зародыша семени (табл. 2).
Таким образом, несмотря на низкий процент плодообразования в ценопопуляции у C. longifolia , проведенные исследования позволили выявить реальную семенную продуктивность, которая оказалась выше, чем в других частях ареала. Наблюдения не показали зависимости уровня плодоношения от освещенности местообитаний. Доля абортивных семян и семян без зародышей оказалась ниже 3,5 %. В 2014 г. общая семенная продуктивность в ценопопуляции составила более 0,5 млн. полноценных семян. Изучение морфологии семян показало относительную географическую изменчивость их линейных размеров. Чтобы спрогнозировать перспективы развития ценопопуляции C. longifolia необходимо провести дополнительную работу. В частности, антэкологические исследования позволят выявить основные факторы, влияющие на показатели продуктивности особей в данном местообитании.
Таблица 2. Морфометрическая характеристика семян Cephalanthera longifolia
|
Показатель |
min-max |
M±m |
CV, % |
|
Длина семени (L s ), мкм |
574,29–1391,24 |
1052,83±13,10 |
12,44 |
|
Ширина семени (W s ), мкм |
154,88–243,70 |
199,90±1,99 |
9,97 |
|
L S / W S |
3,10–6,95 |
5,30 |
14,03 |
|
Длина зародыша (L e ), мкм |
162,96–238,54 |
201,13±1,62 |
8,05 |
|
Ширина зародыша (W e ), мкм |
108,33–160,76 |
126,42±1,06 |
8,41 |
|
L E / W E |
1,30–2,04 |
1,60 |
8,31 |
|
Объем семени (V s ), мкм3х 10 3 |
5153–21593 |
11191±292 |
26,10 |
|
Объем зародыша (V e ), мкм3 х 10 3 |
1041–2836 |
1705±37 |
22,11 |
|
V S / V E |
3,70–11,36 |
6,68 |
24,05 |
|
Объем воздушного пространства в семени (A s ), % |
73,00–91,19 |
84,13±0,40 |
4,74 |
Примечание к табл. 2 : min-max, M, m, CV – см. примечание к табл. 1.
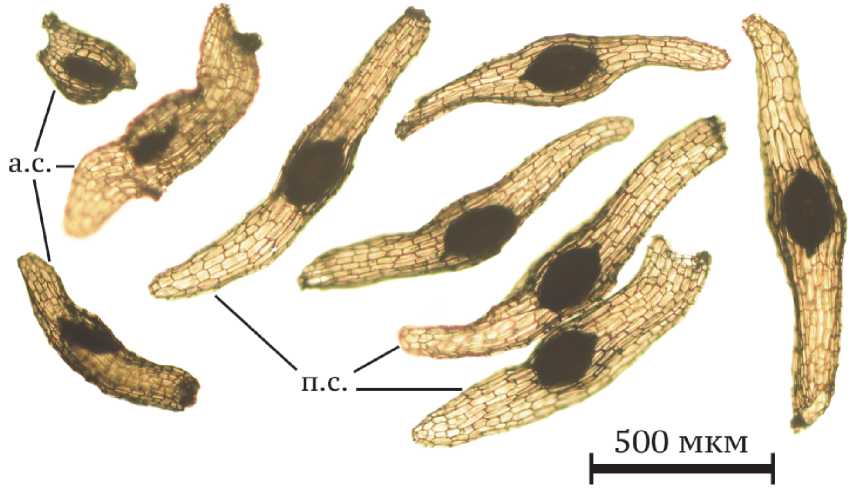
Рис. Внешний вид семян Cephalanthera longifolia :
а.с. – абортивные семена, п.с. – полноценные семена.
Список литературы Особенности семенной продуктивности Cephalanthera longifolia (L.) Fritsch (Orchidaceae juss.) на трансформированных территориях в Калужской области
- Красная книга Российской Федерации (растения и грибы). М.: Товарищество научных изданий КМК, 2008. 855 с.
- Вахрамеева М.Г., Варлыгина Т.И., Татаренко И.В. Орхидные России (биология, экология и охрана). М.: Товарищество научных изданий КМК, 2014. 437 с.
- Мамаев С.А., Князев М.С., Куликов П.В., Филиппов Е.Г. Орхидные Урала: систематика, биология, охрана. Екатеринбург: УрО РАН, 2004. 124 с.
- Красная книга Тульской области (растения и грибы): официальное издание/Под ред. А. В. Щербакова. Тула: Гриф и К, 2010. 393 с.
- Красная книга Тверской области. Тверь: Вече Твери, АНТЭК, 2002. 256 с.
- Литвинов Д.И. Список растений, дикорастущих в Калужской губернии, с указанием полезных и вредных. Калуга, 1895. 112 с.
- Калужская флора: аннотированный список сосудистых растений Калужской области/Н.М. Решетникова, С.Р. Майоров, А.К. Скворцов, А.В. Крылов, Н.В. Воронкина, М.И. Попченко, А.А. Шмытов. М.: Т-во научных изданий КМК, 2010. 548 c.
- Воронкина Н.В., Крылов А.В. Характеристика вида и популяции пыльцеголовника длиннолистного Cephalanthera longifolia (Huds.) Fritsch//Вопросы археологии, истории, культуры и природы Верхнего Поочья: Материалы X Региональной науч. конф. 25-27 марта 2003 г. Калуга: Гриф, 2003. С. 679-686.
- Решетникова Н.М., Крылов А.В. Орхидеи отвалов известняковых карьеров Калужской области//Вестн. ТвГУ. Сер. Биология и экология. 2007. Вып. 4, № 8 (36). С. 111-115.
- Красная книга Калужской области. Калуга: Золотая Аллея, 2006. 608 с.
- Татаренко И.В. Орхидные России: жизненные формы, биология, вопросы охраны. М., Аргус, 1996. 206 с.
- Перебора Е.А. Экология орхидных Северо-Западного Кавказа: монография; под общ. ред. И.С. Белюченко. Краснодар: КубГАУ, 2011. 441 с.
- Перебора Е.А. Орхидные Северо-Западного Кавказа . М., Наука, 2002. 254 с.
- Растения Красной книги России в коллекции ботанических садов и дендрариев. М., 2005. 142 с.
- Наумцев Ю.В., Лебедев А.Н. Редкие виды орхидных Тверской области в Ботаническом саду Тверского госуниверситета//Вестник ННГУ. №3(3). 2014. C. 97-100.
- Коновалова Т.Ю., Шевырева Н.А. Опыт выращивания видов орхидных флоры Московской области в открытом грунте//Восстановление и мониторинг природной флоры. М.: Товарищество научных изданий КМК. 2010. С. 32-39.
- Шибанова Н.Л., Долгих Я.В. Морфометрическая характеристика семян и реальная семенная продуктивнось редких видов орхидных Предуралья//Вестник Пермского университета. Сер. Биология. 2010. Вып. 2. С. 4-6.
- Шибанова Н.Л., Долгих Я.В. Влияние условий освещения на завязывания плодов у 9 видов орхидных Предуралья//Вестник Пермского университета. Сер. Биология. 2011. Вып. 2. С. 4-6.
- Лагутова О.И. Об опылении некоторых видов орхидных Южного берега Крыма//Матер. 8-й межд. конф. «Изучение онтогенеза в ботанических учреждениях Европы и Азии». Киев, 1995. С. 80.
- Назаров В.В. Репродуктивная биология орхидных Крыма: Дис. … канд. биол. наук. СПб., 1995. 294 с.
- Кучер Е.Н., Назаров В.В., Вахрушева Л.П. Затраты на производство цветков и плодов у некоторых видов орхидных Крыма//Бюл. Ботанического сада им. И.С. Косенко: Краснодар, 1998. С. 94-98.
- Кучер Е.Н. Репродуктивное усилие и репродуктивные тактики видов рода Cephalanthera Rich. в Крыму//Ученые записки Таврического национального университета им. В.И. Вернадского. Сер. «Биология, химия». 2014. Т. 27 (66). № 1. С. 93-101.
- Вахрамеева М.Г., Варлыгина Т.И., Куликов П.В. Пыльцеголовник длиннолистный//Биологическая флора Московской области. Вып. 12. М.: «Аргус», 1996. С. 48-59.
- Красная книга Московской области. 2-е изд., доп. и перераб./Отв. ред.: Т.И. Варлыгина, В.А. Зубакин, Н.А. Соболев. М.: Т-во научных изданий КМК, 2008. 828 с.
- Назаров В.В. Определение реальной семенной продуктивности у Dactylorhiza romana и D. incarnata (Orchidaceae Juss.)//Ботан. журн. 1988. Т. 73. № 2. С. 231-233.
- Назаров В.В. Методика подсчета мелких семян и семяпочек (на примере сем. Orchidaceae)//Ботан. журн. 1989. Т. 74. № 8. С. 1194-1196.
- Работнов Т.А. Методы изучения семенного размножения травянистых растений в сообществах//Полевая геоботаника. М.; Л.; 1960. Т. 2. С. 20-40.
- Зайцев Г.Н. Методика биометрических расчетов//Математическая статистика в экспериментальной ботанике. М., 1973. 256 с.
- Буров А.В., Широков А.И., Коломейцева Г.Л. О создании банка семян редких видов орхидных умеренной зоны//Биологический вестник, Т. 8, № 1, 2004, С. 8-11.
- Arditti J., Michaud J.D., Haeley P.L. Morphometry of orchid seeds. I. Paphiopedium and native California and related species of Cypripedium//Amer. J. Bot. 1979. Vol. 66, № 10. P. 1128-1137.
- Healey P.L., Michaud J.D., Arditti J. Morphometry of orchids seeds. III. Native of California and related species of Goodyera, Piperia, Platanthera, and Spiranthes//Amer. J. Bot. 1980. Vol. 67. P. 508-518.
- Dressier R.L. Phylogeny and classification of the orchid family. Portland, Oregon: Dioscorides press, 1993. 314 p.
- Irmisch T. Beiträge zur biologie und morphologie der Orchideen. Leipzig: Ambrosius Abel, 1853. 82 s.
- Фегри К., ван дер Пэйл Л. Основы экологии опыления. М.: Мир, 1982. 377 с.
- Dafni A., Ivri Y. The flower biology of Cephalanthera longifolia (Orchidaceae) -pollen imitation and facultative floral mimicry//Plant Syst. Evol. 1981. Vol. 137. P. 229-240.
- Корженевский В.В., Иванов С.П., Фатерыга А.В., Квитницкая А.А. Виды рода Cephalanthera во флоре Крыма и их антэкологические особенности//Охрана и культивирование орхидей (Материалы IX Международной конференции, Санкт-Петербург, 26-30 сентября 2011 г.). Москва: Товарищество научных изданий КМК, 2011. С. 204-210.
- Cocucci, A.E., Jensen W.A. Orchid embryology: the mature megagametophyte of Epidendrum scutella//Kurtziana. 1969. Vol. 5. P. 23-38.
- Лодкина М.М. Семейство Orchidaceae//Сравнительная анатомия семян. Однодольные/ред. А.Л. Тахтаджян. Л.: Наука, 1985. Т. 1. С. 142-150.
- Шамров И.И. Морфогенез семязачатка и семени у Listera ovata (Orchidaceae)//Ботан. журн. 2001. Т. 86, № 1. С. 3-13.
- Шамров И.И. Семязачаток цветковых растений: строение, функции, происхождение/Под ред. Т.Б. Батыгиной. М.: Товарищество научных изданий КМК, 2008. 350 с.
