Особенности сезонной динамики фотосинтетических пигментов в листьях некоторых древесных интродуцентов в лесостепи Среднего Поволжья

Автор: Кузнецов Р.В., Осипова Е.А., Помогайбин Е.А.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экологическая физиология и биохимия
Статья в выпуске: 1-4 т.11, 2009 года.
Бесплатный доступ
В статье представлены данные относительно сезонной динамики фотосинтетических пигментов в листьях древесных растений - представителей родовых комплексов боярышник, клен, орех в насаждениях дендрария ботанического сада Самарского государственного университета.
Интродуценты, фотосинтетические пигменты, сезонная динамика, листья, адаптация
Короткий адрес: https://sciup.org/148198419
IDR: 148198419 | УДК: 581.5
Concerning the seasonal dynamics of photosynthetic pigments in some tree species introduced in forest-steppe Middle Povolzhye
Some new data concerning the seasonal dynamics of photosynthetic pigments are given for some representatives of species complexes Acer, Crataegus, Juglans growing in dendrarium of Botanical Garden of Samara State University.
Текст научной статьи Особенности сезонной динамики фотосинтетических пигментов в листьях некоторых древесных интродуцентов в лесостепи Среднего Поволжья
В ходе интродукции растения, переносимые за пределы своих природных ареалов, оказываются в новых природных условиях. При переносе в более неблагоприятные для своего роста местности или условия растения не приспосабливаются, а, защищаясь с разной степенью успеха, борются за свое существование. При несоответствии условий среды комплексу наследственных свойств данного организма он в зависимости от степени этого несоответствия и от своей видовой и онтогенетической жизнеспособности, в том числе потенциала адаптации, живет в ухудшенных условиях различный период времени, но в итоге гибнет или вырождается в ближайших поколениях. Не только сколько-нибудь значительные географические перемещения, но и менее существенные попытки вывести интроду-центы из состояния оптимума (или зоны комфорта), по-видимому, не проходят для них бесследно. Связь всех частей организма между собой и с условиями среды приводит его к необходимости компенсировать недостающий экологический фактор соответствующим изменением некоторых своих функций, например повышением интенсивности дыхания, сокращением силы роста и другими процессами автооптимизации. Борьбу за сохранение существования по мере ухудшения условий организм ведет различными средствами адаптации до некоторой критической точки, или линии пессиму-ма, за которой он уже не живет, а начинает гибнуть, исчерпав видовую и онтогенетическую амплитуду возможной изменчивости своих функций [3].. Любые вредные воздействия на организм, в том числе и термические, компенсируются путем мобилизации одних и тех же его жизненных сил или жизненного потенциала, который расходуется при этом, по-видимому, без существенного восполнения, несмотря на внешнее, подчас достаточно полное восстановление нормы физиологических или морфологических признаков.
При выводе растений из их естественного ареала при нарастающем ухудшении условий обитания, по мере удаления особи от зоны оптимума вида, адаптационные изменения физиологических функций и морфологических признаков, оставаясь обратимыми, количественно нарастают вплоть до линии пессимума, которая географически ограничивает допустимую для жизнедеятельности видовую амплитуду обитания. За пределами критической линии пессимума адаптационные изменения становятся патологическими и необратимыми, и в зависимости от удаления от нее особь погибает с большей или меньшей скоростью. В первую очередь при интродукционных перемещениях наступают адаптационные изменения физиологических функций. При дальнейшем, более сильном ухудшении условий среды начинаются изменения вегетативной сферы особи (уменьшение роста, увеличение ветвления и густоты куста, переход древесной или кустарниковой формы в близкую к травянистым растениям). Адаптационные изменения как обратимые, так и необратимые большей частью серьезно не затрагивают органов репродуктивной сферы, что и понятно в отношении сохранения видовой индивидуальности.
Листовой аппарат цветковых растений представляет собой эффективную систему фотосинтезирующих органов, обнаруживающих достаточно широкие адаптационные возможности. Он демонстрирует проявление структурно-функционального единства, является примером реализации принципа избыточности за счет наличия множества дублирующих идентичных структур [1]. На фоне видоспецифичных особенностей, структура листьев обнаруживает зависимость от комплекса биотопиче-ских условий, среди которых важнейшая роль принадлежит освещенности. Это приводит к формированию даже в кроне одного и того же древесного растения листьев «световой» и «теневой» структуры [4]. Особенности тканевого строения и биохимические показатели листовых пластинок зависят также от гидротермических условий местообитания, от особенностей почвенного субстрата [6, 7]. Эти адаптационные возможности позволяют устанавливать соответствие между экологическими условиями и уровнем метаболических процессов в листьях.
Как известно, в условиях умеренной зоны сезонная динамика ряда биохимических показателей листовой массы характеризуется чертами сходства у различных древесных видов и согласуется с особенностями развития и функционирования листьев в течение вегетационного периода [2]. Так, в годы со «средними» летними условиями содержание фотосинтетических пигментов в листьях древесных растений достигает максимума в июне-июле, к августу несколько снижается, к сентябрю в зависимости от пункта наблюдений в городских насаждениях находится на уровне одной четвертой – одной шестой от сезонного максимума. Однако засушливые условия в сочетании с высокими температурами, возникая в разные сроки внутри вегетационного периода, могут смещать и сроки достижения сезонных максимумов, и продолжительность их поддержания в листьях, и скорость убывания пигментов в стареющих листьях.
В данной работе мы хотели бы представить данные относительно особенностей сезонной динамики фотосинтетических пигментов в листьях древесных растений-представителей родовых комплексов боярышник, клен и орех, изученных нами на примере насаждений дендрария ботанического сада Самарского государственного университета.
МЕТОДИКА РАБОТЫ
Изучение сезонной динамики пигментов для представителей различных родовых комплексов проводили в 2004 – 2008 гг. (для кленов – 20042006, для боярышников – 2004-2005, для орехов – 2008 г.) в дендрарии ботанического сада Самарского государственного университета. Пробы листьев древесных растений-интродуцентов, находящихся в генеративной стадии развития, отбирали на высоте 1,5 м по периметру кроны, высушивали в условиях лаборатории и после измельчения фитомассы в лабораторной мельнице проводили спектрофотометрическое определение содержания фотосинтетических пигментов в ацетоновых экстрактах [5]. Данные обрабатывали с использованием пакета программ Excel.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В данном сообщении мы используем результаты оценки количества пигментов, проведенной не только у разных видов древесных интродуцентов, но и в разные годы наблюдений. Кратко укажем, что вегетационный период 2004 г. в целом характеризовался высоким количеством осадков за исключением жаркого и засушливого августа; 2005 г. – условия влагодефицита отмечались в мае и особен- но июле-августе; 2008 – некоторый дефицит влаги в мае и августе.
Для растений – представителей родовых комплексов боярышники и клен перечень анализируемых видов включает местные и интродуцированные (боярышник кровяно-красный – аборигенный, интродуценты - боярышники Максимовича, пери-стонадрезанный, полумягкий, хлорокарпа (желтоплодный), даурский; клены остролистный и татарский – аборигенные, интродуценты – клены ясенелистный, полевой, завитой, Семенова, серебристый, голый). В целом и уровни содержания фотосинтетических пигментов, и общий характер их динамики у местных видов и интродуцентов внутри одного родового комплекса были сходны. Реакция на наступление засушливых условий обнаружила некоторые видоспецифические различия.
Так, у аборигенного боярышника кровянокрасного и наиболее адаптировавшегося к местным условиям боярышника полумягкого дефицит осадков в августе привел к ускоренному старению листьев, на фоне быстрого накопления суммы активных температур ускорилась подготовка к завершению вегетационного периода. Сходную динамику обнаружил и боярышник даурский. У боярышников перисторассеченного и хлорокарпа изобилие осадков в июле спровоцировало рост количества пигментов, чего у других видов не наблюдалось.
У всех видов рода клен в той или иной степени проявилась способность к синтезу дополнительного количества пигментов в конце вегетационного периода после снятия неблагоприятного воздействия (засуха в августе) и восстановления высокой влагообеспеченности. В виде тенденций видоспецифические различия продемонстрировали сами количественные показатели и выраженность июльского максимума.
Что касается видов рода орех, в природной дендрофлоре региона они отсутствуют. Среди изучавшихся растений были представлены виды и формы (орехи айлантолистный, сердцевидный, черный (2 представителя разного происхождения), маньчжурский, серый, скальный, грецкий, грецкий форма скороплодная). При различной структуре листовых пластинок, что проявляется как в их площади, так и в характере структуры (толщина, плотность жилкования, мера склерофильности) были существенно выражены видоспецифические различия по уровню содержания пигментов. Сезонная динамика также не совсем совпадала, что может означать неодинаковый уровень соответствия местных условий экологическому оптимуму видов. Лишь у некоторых растений наблюдался
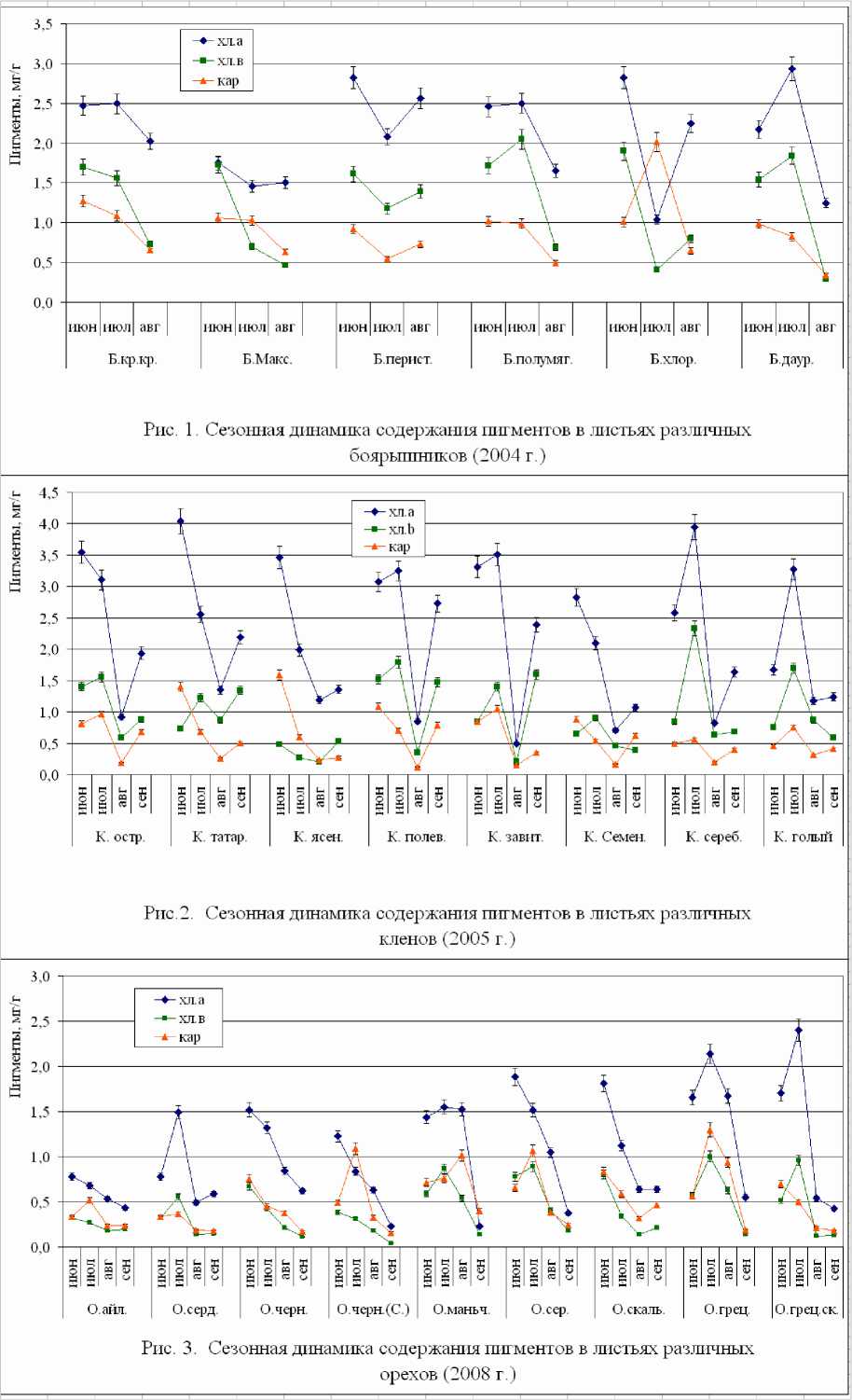
дополнительный синтез пигментов к сентябрю, после засушливых августовских условий. Наоборот, у большинства видов было отмечено высокое (зачастую превышающее количество хлорофилла В) содержание каротиноидов в зрелых листьях, что может быть отнесено к особенностям данного рода вообще.
ЗАКЛЮЧЕНИЕ
Таким образом, при интродукции в лесостепи Среднего Поволжья (г. Самара) древесные виды-интродуценты, представители родов боярышник, клен и орех демонстрируют на фоне закономерной сезонной динамики фотосинтетических пигментов различия, которые предположительно связаны с их неодинаковыми адаптационными возможностями в районе интродукции.
Список литературы Особенности сезонной динамики фотосинтетических пигментов в листьях некоторых древесных интродуцентов в лесостепи Среднего Поволжья
- Гродзинский Д.М. Надежность растительных систем. Киев: Наукова думка, 1983. 368 с.
- Кавеленова Л.М. Проблемы организации системы фитомониторинга городской среды в условиях лесостепи. Самара; Самарский университет, 2006. 200 с.
- Зайцев Г.Н. Оптимум и норма в интродукции растений. М.: Наука, 1983. 269 С
- Лотова Л.И. Ботаника: Морфология и анатомия высших растений. М.: КомКнига, 2007. 512 с.
- Методы биохимического исследования растений/Под ред. А.И.Ермакова. М.: Агропромиздат, 1987. 480 с.
- Bussotti F., Borhgini F., Celesti C., Leonzio C., Bruschi P. Leaf morphology and macronutrients in broadleaved trees in central Italy//Trees. 2000. V. 14. No. 7. P. 361-368.
- Kolb T.E., Hart S.C. Boxelder water sources and physiology at perennial and ephemeral stream sites in Arizona//Tree Physiology. 1997. V. 17. P.151-160.
