Особенности строения и развития женских генеративных структур у линий кукурузы с наследуемым и индуцированным типами партеногенеза

Автор: Апанасова Наталия Владимировна, Гуторова Ольга Валентиновна, Юдакова Ольга Ивановна, Смолькина Юлия Валерьевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-2 т.19, 2017 года.
Бесплатный доступ
В статье приводятся результаты цитоэмбриологического анализа линий кукурузы c разными типами цитоплазмы и наследуемым партеногенезом (АТ-3 (N), АТ-3 (М), АТ-3 (С), АТ-3 (Т), АТ-3 (В)), а также гаплоиндуцирующих линий с ненаследуемым типом партеногенеза (ЗМС-П1, ЗМС-П2, ЗМС-П3). Установлено, что для партеногенетических форм кукурузы с N, М, Т, B и С типами цитоплазмы наряду с развитием партеногенетических зародышей (2,3-5,5%) характерно формирование в зародышевых мешках дополнительных яйцеклеток (0,3-10%). У линий с М, B и С типами цитоплазмами встречались мегагаметофиты с двумя партеногенетическими проэмбрио (0,8; 0,7 и 0,5% соответственно). Такие гаметофитные аномалии могут быть использованы как диагностические признаки при поиске и отборе новых партеногенетических форм у кукурузы. У линий с высокой гаплоиндуцирующей способностью ЗМС-П1, ЗМС-П2, ЗМС-П3 партеногенетического развития яйцеклеток в неоплодотворенных зародышевых мешках не зарегистрировано. У линий ЗМС-П1 и ЗМС-П3 мегагаметофиты аномального строения (с дополнительными яйцеклетками и полярными ядрами) были единичными, а у линии ЗМС-П2 полностью отсутствовали.
Гаплоидия, апомиксис, партеногенез, матроклинные гаплоиды, гаплоиндукция in vivo, зародышевый мешок, проэмбрио, кукуруза
Короткий адрес: https://sciup.org/148205120
IDR: 148205120 | УДК: 581.331.1:633.15
Peculiarities of structure and development of female generative structures in corn lines with heritable and nonheritable types of parthenogenesis
The cytoembryological analysis data of corn lines with the inherited parthenogenesis with different types of cytoplasm (AT-3 (N), AT-3 (M), AT-3 (S), AT-3 (T), AT-3 (V)) and the haploid inducer lines with non-inherited parthenogenesis (ZMS-P1, ZMS-P2, ZMS-P3) are given in the article. It is found that the AT-3 (N), AT-3 (M), AT-3 (S), AT-3 (T), AT-3 (V) corn lines are characterized both parthenogenetic embryos development (2.3-5%) and the formation of additional egg cells in the embryo sacs (0.3-10%). These gametophyte anomalies can be used as diagnostic features in the search and selection of new parthenogenetic forms in corn. In the ZMS-P1, ZMS-P2, ZMS-P3 lines in the unfertilized embryo sacs the egg cells parthenogenetic development was not detected. In the plant of the ZMS-P1 and ZMS-P3 lines the gametophytic anomalies (additional egg cells and additional polar nuclei) were recorded in the single embryo sacs. In the ZMS-P2 line anomalies were absent.
Текст научной статьи Особенности строения и развития женских генеративных структур у линий кукурузы с наследуемым и индуцированным типами партеногенеза
У такой важной сельскохозяйственной культуры как кукуруза существуют формы как с наследуемым, так и ненаследуемый типами партеногенеза. Наследуемый тип гаплоидии представляет интерес для синтеза нередуцированных апомиктов на основе объединения признаков партеногенеза, не-редукции и нормального развития эндосперма [1]. Ненаследуемый (индуцированный) тип гаплоидии в настоящее время широко используется для ускоренного создания гомозиготных линий кукурузы, и на их основе высокогетерозисных гибридов [7, 8]. Создано немало гаплоиндуцирующих линий с высокой частотой гаплоиндукции, пригодных для коммерческого использования. К ним относятся полученные от линии-основателя Stock 6 линии: КМС и ЗМС [9], WS14 [10], MHI [11], RWS [12], UH400 [13], HZI1, CAUHOI [14], PHI [15], KEMS [16] и др. Технология гаплоиндукции включена в различные коммерческие селекционные программы для кукурузы в Европе, Северной Америке, в Китае [14], Пакистане [17], Вьетнаме и других странах [18, 19]. Несмотря на большое количество полученных гапло-индукторов создание новых линий кукурузы с гап-лоиндуцирующей способностью остается актуальным. Целесообразность таких работ обусловлена необходимостью адаптации к разным климатическим условиям, повышения частоты гаплоиндукции и введения новых маркерных признаков, позволяющих быстро отбирать гаплоиды среди гибридного потомства. Эффективное использование в селекционно-генетических программах линий кукурузы с различными типами партеногенеза невозможно без учета их эмбриологических особенностей.
Цель работы: изучение особенностей строения и развития женских генеративных структур у линий кукурузы с наследуемым и индуцированным типами партеногенеза.
Материал и методика исследования. Материалом исследования послужили линии кукурузы c разными типами цитоплазмы и наследуемым партеногенезом (АТ-3 (N) с нормальным исходными типом цитоплазмы; АТ-3 (М) с молдавским типом цитоплазматической мужской стерильности; АТ-3 (С) с парагвайским типом ЦМС; АТ-3 (Т) с техасским типом ЦМС; АТ-3 (В) с боливийским типом ЦМС), а также линии с высокой гаплоиндуцирую-щей способностью (ЗМС-П1, ЗМС-П2, ЗМС-П3). В качестве контроля использовали линию ЗМgl, не обладающую гаплоиндуцирующей способностью и не склонную к наследуемому партеногенезу.
Растения высаживали в поле квадратногнездовым методом, 16 делянок по 4 ряда. Початки изолировали до появления пестичных нитей пергаментными пакетами. Фиксацию ацетоалкоголем проводили на 7 сутки после появления первых пестичных нитей. Препараты женских гаметофитов приготавливали с использованием метода ферментативной мацерации семязачатков с последующей диссекцией [20]. Анализ препаратов проводили с использованием микроскопа AxioStar (С. Zeiss, Германия).
Результаты исследования.
Строение и развитие женских генеративных структур линии кукурузы, несклонной к партеногенезу (контроль). Все исследованные зрелые зародышевые мешки линии ЗМgl имели типичное для злаков строение. Они были восьмиядерные, семиклеточные, Polygonum-типа. Яйцевой аппарат трехклеточный, состоящий из одной яйцеклетки и двух синер-гид. Центральная клетка с двумя полярными ядрами, несливающимися до оплодотворения. В процессе созревания зародышевого мешка антиподы претерпевают несколько митотических делений, в результате чего формируется антиподальный комплекс, состоящий в среднем из 15-20 одноядерных антипод. Гаметофитных аномалий и партеногенетического развития яйцеклеток у изученных растений линии ЗМgl не обнаружено (см. табл. 1).
Структура женских гаметофитов неопыленных завязей изученных линий кукурузы
|
Линия |
Всего исследовано зародышевых мешков |
Количество зародышевых мешков, % |
||||||
|
нормального строения |
с допол-нитель-ными яйцеклетками |
с дополнительными полярными ядрами |
с автономным проэм-брио |
недиф-ферен-циро-ванные |
||||
|
всего |
с одним |
с двумя |
||||||
|
ЗМgl (контроль) |
207 |
100,0 |
0,0 |
0,0 |
0,0 |
0,0 |
0,0 |
0,0 |
|
Линии с наследуемым типом партеногенеза |
||||||||
|
АТ-3 (N) |
300 |
96,1 |
0,3 |
0,0 |
3,3 |
3,3 |
0,0 |
0,3 |
|
АТ-3 (М) |
300 |
86,6 |
8,3 |
0,0 |
3,1 |
2,3 |
0,8 |
0,0 |
|
АТ-3 (В) |
300 |
86,7 |
10,0 |
0,0 |
3,0 |
2,3 |
0,7 |
0,3 |
|
АТ-3 (С) |
200 |
88,0 |
6,0 |
0,0 |
6,0 |
5,5 |
0,5 |
0,0 |
|
АТ-3 (Т) |
200 |
94,0 |
2,5 |
0,0 |
3,5 |
3,5 |
0,0 |
0,0 |
|
Линии с индуцированным типом партеногенеза (гаплоиндукторы) |
||||||||
|
ЗМС-П1 |
256 |
99,6 |
0,4 |
0,0 |
0,0 |
0,0 |
0,0 |
0,0 |
|
ЗМС-П2 |
172 |
100,0 |
0,0 |
0,0 |
0,0 |
0,0 |
0,0 |
0,0 |
|
ЗМС-П3 |
207 |
99,5 |
0,0 |
0,5 |
0,0 |
0,0 |
0,0 |
0,0 |
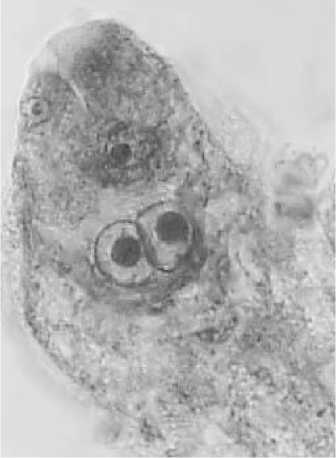
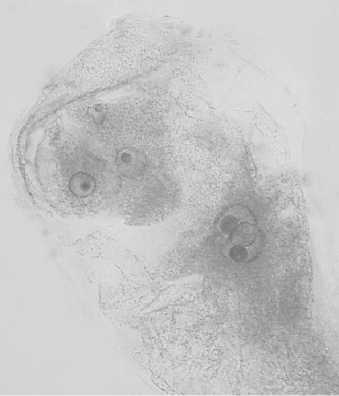
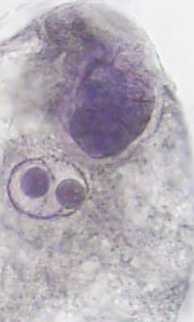
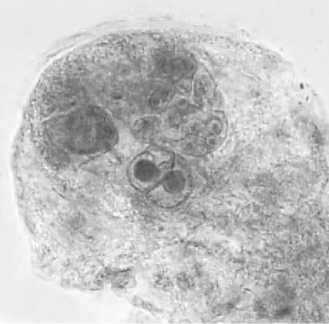
Строение и развитие женских генеративных структур линий кукурузы с наследуемым типом партеногенеза. У растений линий кукурузы АТ-3 (N), АТ-3 (М), АТ-3 (С), АТ-3 (Т) и АТ-3 (В) в неопылен-ных завязях зародышевые мешки с партеногенетическим проэмбрио встречались с частотой от 3,0 до 6,0% (см. табл. 1, рис. 1в). Наряду с мегагаметофитами нормального строения (рис. 1а) были обнаружены зародышевые мешки с дополнительными яйцеклетками (одной, иногда двумя) (рис. 1б). Дополнительные женские гаметы формировались за счет нарушения процессов поляризации и дифференцировки элементов зародышевого мешка. Такие нарушения чаще всего приводили к образованию трехклеточного яйцевого аппарата, состоящего из двух яйцеклеток и одной синергиды (рис. 1б) или в некоторых случаях из трех яйцеклеток. Реже наблюдалось формирование четырехклеточного яйце- вого аппарата, состоящего из двух синергид и двух яйцеклеток. Дополнительные женские гаметы могли развиваться партеногенетически, о чем свидетельствует наличие в неопыленных завязях зародышевых мешков с двумя проэмбрио (рис. 1г). Такие мегагаметофиты были зарегистрированы у линий с цитоплазмами М, B и С типами с частотой 0,8; 0,7 и 0,5%, соответственно (см. табл. 1).
Строение и развитие женских генеративных структур линий кукурузы с индуцированным типом партеногенеза. Практически все исследованные зрелые зародышевые мешки линий ЗМС-П1, ЗМС-П2, ЗМС-П3 имели типичное строение (см. табл. 1). Лишь в одном зародышевом мешке линии ЗМС-П1 было обнаружено две яйцеклетки, и в одном мегагаметофите линии ЗМС-П3 – три полярных ядра. Из более чем 600 проанализированных зародышевых мешков ни в одном из них не обнаружено партено- генетического развития яйцеклеток, тогда как при самоопылении данных линий гаплоиды встречались с частотой около 8% [19].
Это свидетельствует о том, что:
-
1) гаплоиндуцирующие линии не склонны к образованию гаплоидов за счет наследуемого партеногенеза;
а
б
в
г
Рис.1. Зародышевые мешки кукурузы линии АТ-3:
1 - типичного строения; 2 - с двумя яйцеклетками в яйцевом аппарате; 3 - с партеногенетическим проэмбрио; 4 - с двумя партеногенетическими проэмбрио
-
2) развитие гаплоидов у них индуцируется после опыления собственной пыльцой. Способность гап-лоиндуцирующих линии индуцировать образование гаплоидов при самоопылении дает возможность при создании новых гаплоиндуцирующих линий проводить отбор на гаплоиндукцию в самоопылен-ном потомстве.
Выводы: проведенное исследование дает основание констатировать, что развитие женского гаметофита у линий кукурузы, не склонных к партеногенезу, и гаплоиндуцирующих линий ЗМС-П1, ЗМС-П2, ЗМС-П3 происходит, как правило, без каких-либо отклонений от нормы. При задержке опыления 7 сут индукции яйцеклеток к партеногенезу не наблюдается. У линий с высокой гаплоиндуци-рующей способностью партеногенетические зародыши развиваются только после опыления собственной пыльцой. У линий кукурузы с наследуемым типом партеногенеза АТ-3 (М), АТ-3(С), АТ-3 (Т), АТ-3 (В) при задержке опыления 7 сут зародыши без оплодотворения развиваются в среднем с частотой около 3%. Данные линии характеризуются также повышенной частотой формирования зародышевых мешков с дополнительными яйцеклетками от 0,3% у АТ-3(М) до 10,0% у АТ-3(В). Развитие мегагаметофитов со структурными отклонениями - закономерное эмбриологическое явление, сопутствующее апомиксису у дикорастущих злаков [21]. С наибольшей частотой у них встречается полигаметия -формирование в мегагаметофите дополнительных яйцеклеток за счет трансдетерминации синергид или антипод. Повышенная частота полигаметии у апомиктичных дикорастущих злаков и партеногенетических форм кукурузы свидетельствует о том, что образование дополнительных яйцеклеток является признаком, сопряженным с наиболее важным элементом апомиксиса партеногенезом. Высокая частота встречаемости дополнительных женских гамет дает возможность использовать эту эмбриологическую особенность в качестве диагностического признака при выявлении склонности к наследуемым формам партеногенеза на эмбриологическом уровне и при проведении искусственного отбора на наследуемый партеногенез у кукурузы. Линии АТ-3 (М), АТ-3(С), АТ-3 (Т), АТ-3 (В) были получены от одной исходной линии АТ-3 путем перевода ее на разные формы цитоплазмы с ЦМС. Развитие партеногенетических проэмбрио и формирование дополнительных гамет у этих линий свидетельствует о том, что данные признаки являются ядерными, на характер проявления которых цитоплазма не оказывает влияние.
Работа выполнена при частичной финансовой поддержке Минобрнауки России в рамках базовой части государственного задания в сфере научной деятельности по заданию №6.8789.2017/БЧ.
Список литературы Особенности строения и развития женских генеративных структур у линий кукурузы с наследуемым и индуцированным типами партеногенеза
- Тырнов, В.С. Гаплоидия у растений: терминология и классификация: Учеб. пособие для студентов биол. фак. -Саратов: Изд-во Сарат. ун-та, 2005. 44 с.
- Тырнов, В.С. Гаплоидия и апомиксис//Репродуктивная биология, генетика и селекция. -Саратов: Изд-во Сарат. ун-та, 2002. С. 32-46.
- Тырнов, В.С. Прикладные аспекты гаметофитного апомиксиса/Эмбриология цветковых растений: терминология и концепции. -СПб: Мир и семья, 2000. Т. 3. С. 203-206.
- Чалык, С.Т. Методы гаплоидии в генетике и селекции кукурузы. -Кишинев, 2003. 179 с.
- Dwivedi, S.I. Haploid: constraints and opportunities in plant breeding/S.I. Dwivedi, A.B. Britt, L. Tripathi et al.//Biotechnology Advances. 2015. V. 33. P. 812-829.
- Dunwell, J.M. Haploids in flowering plant: origins and exploitation//Plant Biotechnol. J. 2010. V. 8. P. 377-424.
- Gallais, A. The use of doubled haploids in recurrent selection and hybrid development in maize/A. Gallais, J. Bordes//Crop Sci. 2007. V. P. 190-201.
- Coe, E.H. A line of maize with high haploid frequency//Am. Nat., 1959. V. 93. P. 381-382.
- Тырнов, В.С. Индукция высокой частоты возникновения матроклинных гаплоидов у кукурузы/В.С. Тырнов, В.Н. Завалишина//Докл. АН СССР. 1984. Т. 276. № 3. C. 735-738.
- Lasharmes, P. Genetic control of maternal haploidy in maize (Zea mays L.) and selection of haploid inducing lines/P. Lasharmes, M. Bekert//Theor. Appl. Genet. 1988. V. 76. P. 404-410.
- Eder, J. In vivo haploid induction in maize/J. Eder, S. Chalyk//Theor Appl Genet, 2002. V. 104. P. 703-708.
- Rober, F.K. Fortpflanzungsbiologische und genetische Untersuchungen mit RLFP-makern zur in vivo -Haploideninduktion bei Mais. Verlag Grauer. Stutgart, Gemany, 1999. 320 p.
- Chang, M.T. Doubled haploids / M.T. Chang, E.N. Coe // In: Al Kriz, Ba Larkins (edds) // Biotechnology in Agriculture and forestry, 2009. V.63. Molecular Genetic Approaches to Maize Improvement. - Springer Verlag, Berlin, Heidelberg. P. 127-142.
- Prasanna, B.M. Dounled haploid technology in maize breeding: Theory and practice/B.M. Prasanna, V. Chaikam, G. Mahuku//CIMMYT, Mexico, DF, 2012. 53 p.
- Rotarenco,V. New inducer of maternal haploids in maize/V. Rotarenco, G. Dicu, D. State, S. Fuia//Maize Genet. Coop. Newsletter. 2010. V. 84.
- Шацкая, О.А. Создание гаплоиндукторов кукурузы: три цикла отбора на высокую частоту индукции матроклинных гаплоидов//Сельскохозяйственная биология. 2010. № 5. С. 79-86.
- Khakwwani, K. Development of Maize Haploid Inducer Lines and Doubled Haploid Lines in Pakistan/K. Khakwwani, M.R. Dogar, M.A.A. Hussain et al.//British Biotechnology Journal. 2015. V. 8(3). P.1-7.
- Hu, H. The genetic basis of haploid induction in maize identified with a novel genome-wide association method/H. Hu, T.A. Schrag, R. Peis et al.//Genetics. 2016. V. 202. P. 1267-1276.
- Гуторова, О.В. Создание генетически маркированных линий кукурузы с наследуемым и индуцированным типами партеногенеза/О.В. Гуторова, Н.В. Апанасова, О.И. Юдакова//Известия Самарского научного центра Российской академии наук. 2016. Т. 18, №2(2). С. 341-344.
- Юдакова, О.И. Методы исследования репродуктивных структур и органов растений: учеб.-метод. пособие для студентов биол. фак./О.И. Юдакова, О.В. Гуторова, Ю.А. Беляченко. -Саратов: Изд-во Сарат. ун-та, 2012. 44 с.
- Шишкинская, Н.А. Популяционная эмбриология и апомиксис у злаков/Н.А. Шишкинская, О.И. Юдакова, В.С. Тырнов. -Саратов: Изд-во Сарат. ун-та, 2004. 148 с.
