Особенности твердосемянности Cuscuta japonica Choisy, C. campestris Yunck. (Cuscutaceae Dum.)
")
Автор: Леусова Н.Ю.
Журнал: Международный журнал гуманитарных и естественных наук @intjournal
Рубрика: Биологические науки
Статья в выпуске: 12-1 (51), 2020 года.
Бесплатный доступ
Рассмотрены особенности твердосемянности двух видов повилик Cuscuta japonica Choisy и C. campestris Yunck. (Cuscutaceae Dum.), влияние эколого-географических условий на твердосемянность. Показано, что в лабораторных условиях для преодоления покоя семян данных видов, необходима химическая стратификация в концентрированной серной кислотой (H2SO4 конц.). Растения-хозяева не влияют на прорастание семян повилики японской, следовательно, утверждение о влиянии растений-хозяев на прорастание не является всеобщим, по крайней мере, для данного вида.
Повилика, прорастание семян, покой семян, твердосемянность, скарификация
Короткий адрес: https://sciup.org/170190708
IDR: 170190708 | DOI: 10.24411/2500-1000-2020-11595
The features of hard coat of Cuscuta japonica Choisy and C. Campestris Yunck. (Cuscutaceae Dum.)
The features of hard coat of two species of dodder - Cuscuta japonica Choisy and C. campestris Yunck. (Cuscutaceae Dum.), The influence of ecological and geographical conditions on hard coat for example, the Amur region. It is shown that under laboratory conditions to overcome the dormancy of these species, requires chemical stratification in concentrated sulfuric acid (H2SO4 conc.). Host-plants do not affect the Japanese dodder seed germination, therefore, the statement about the impact of the plant-hosts on the germination is not universal, at least for the species.
Текст научной статьи Особенности твердосемянности Cuscuta japonica Choisy, C. campestris Yunck. (Cuscutaceae Dum.)
Введение. Любое антропогенное воздействие постепенно становится основным фактором, определяющим динамику современного распространения организмов. Вторжение или вымирание видов, иногда даже одного может приводить к радикальной перестройке сообществ [1]. Паразитические растения представляют особую группу, но биология их изучена недостаточно. И не всегда бывает понятно, почему одни виды, становятся космополитами, другие же, наоборот, имеют ограниченное распространение. Исключительно паразитический образ жизни повилик (Cuscuta L.) создает огромные трудности в борьбе с ними. В полной мере это касается Cuscuta japonica Choisy и C. campestris Yunck. (Cuscutaceae Dum.), имеющих широкий круг хозяев [2]. Одной из особенностей повилик как паразитов является большое количество семян, которое исчисляется тысячами на одно растение. Семена созревают постепенно – период покоя может длиться 4-5 лет, а, по некоторым сведениям, до 20 лет [3]. Поэтому изучение природы покоя семян и условий его преодоления имеет первостепенное значение. Для многих видов рода Cuscuta наблюдается одно из проявлений покоя – состояние твердосемянности (полная водонепроницаемость кожуры, реже – околоплодника), что влияет на динамику прорастания, а также делает семена устойчивыми к соляризации почв. В естественных экосистемах деятельность входящих в них видов, как правило, направлена на поддержание этих экосистем как среды собственного обитания.
В «Справочнике по проращиванию покоящихся семян» [4] есть сведения по прорастанию некоторых видов повилик, однако данные по С. japonica Choisy и C. campestris Yunck отсутствуют. Как показывают наши предварительные эксперименты, низкая всхожесть семян повилик также обусловлена твердосемянностью [5, 6]. По данным J.M. Hutchison и F.M. Ashton [7] всхожесть недозревших семян повилики полевой составляет 80%. В процессе дозревания происходит естественное высушивание семян, уменьшается проницаемость и повышается прочность семенной оболочки, в результате чего у семян наступает вторичный покой и их всхожесть снижается до 2%. Согласно данным J.H. Dawson [8], более 90% зрелых семян имеют твердую оболочку. Такие семена длительно сохраняют свою жизнеспособность и имеют растянутый период прорастания. Скарификация твердых семян наждачной бумагой, накалыванием или обработкой серной кислотой нарушает покой и повышает всхожесть семян.
Какие факторы играют первостепенную роль и в значительной степени влияют на снятие покоя семян повилики в естественных условиях? – вопрос остается до настоящего времени дискуссионным. Немаловажную роль оказывает, например, глубина залегания семян в почве, выгорание растительных сообществ (тепловая обработка), зимний период, поедание животными и др. Открытым остается и значение химических сигналов во взаимодействии хозяин-повилика (паразит), которое показано для различных паразитических животных и растений. Ответ паразита на химический сигнал хозяина отмечен для некоторых паразитических цветковых растений [9]. Со времени исследования 1913 года считается, что семена повилики ускоренно прорастают, реагируя на выделения растений-хозяев. Для проростков повилики вещества, выделяемые растением-хозяином, необходимы для успешного прикрепления и перехода к паразитическому образу жизни. Это утверждение вошло во все российские и зарубежные учебники в качестве аксиомы. Так, С. Тарр [10] обосновывает, что прорастание, очевидно, происходит под действием корневых выделений некоторых растений, которые, однако, не обязательно являются хозяевами данного паразита. Стимуляция прорастания семян паразита корневыми выделениями растений обеспечивает паразиту определенные преимущества, так как они повышают вероятность успешного заражения. Тем не менее, круг таких исследований ограничен определенными видами корневых паразитов [9, 11].
Целью наших исследований было изучение особенностей прорастания семян аборигенного вида C. japonica и адвентивного C. campestris , в условиях Амурской области.
Материал и методы исследований. Объектом исследований служили зрелые семена Cuscuta japonica и C. campestris собранные в 2015-2017 гг. Для опытов отбирали семена, одинаковые по размеру, без видимых повреждений. Семена в количестве 25 штук помещали в чашки Петри на увлажненную фильтровальную бумагу (субстрат) и проращивали при комнатной температуре. Проросшими считали наклюнувшиеся семена, у которых длина проклюнувшихся осевых органов превышала половину ширины семени.
Химическую скарификацию вызывали обработкой семян концентрированной серной кислотой (H2SO4 конц.) с различ- ным временем выдерживания: 10 мин., 15 мин., 20 мин., 30 мин., 40 мин., 1 час. В последующем, семена промывали проточной водой и в количестве 25 штук помещали в чашки Петри на увлажненную фильтровальную бумагу, проращивание проводили при комнатной температуре. В качестве контроля использовали семена, необработанные кислотой. Химическую обработку проводили в соответствии с требованиями, указанными в «Справочнике по проращиванию покоящихся семян» М.Г. Николаевой с соавторами [1985]. По прорастанию семян после такой обработки оценивали их жизнеспособность. Семена, не прораставшие в течение 4 дней, считали нежизнеспособными. Измерения проводили в 4-кратной биологической повторности.
Результаты исследования. Твердые семена сельскохозяйственных культур обычно перед посевом подвергают различного рода физическим или химическим обработкам. Используют скарификацию, замачивание в концентрированной серной кислоте, ошпаривание кипятком и т.д. [4]. Причины, вызывающие торможение прорастания покоящихся семян, различны, и, следовательно, условия нарушения покоя. М.Г. Николаева с соавторами [4] указывают, что у семян одних и тех же растений твердосемянность проявляется не всегда, и зависит от степени зрелости семян, условий созревания и хранения. Глубина покоя варьирует не только у разных видов, но нередко в пределах одного вида и даже растения. В связи с вышесказанным способ обработки должен устанавливаться опытным путем.
Полученные нами ранее [12] и в настоящее время опытные данные показывают (рис. 1), что семена повилики японской и повилики полевой характеризуются низкой лабораторной всхожестью (3 и 14%, соответственно) при нормальных температурных условиях (22-250C, поскольку оптимум для большинства видов лежит в диапазоне 10-30 0С) и высоким процентом твердых семян (97 и 86, соответственно).
Изучение твердосемянности выявило узкий диапазон варьирования данного признака в зависимости от генотипа. При изучении семян каждого вида установлено, что твердосемянность у исследуемых повилик колебалась соответственно от 9698% для повилики японской и 84-88% для повилики полевой. Нами также отмечено, что при хранении семян в течение 15 лет (семена урожая 2005 года) не теряется их жизнеспособность, а степень твердосе-мянности остается на прежнем уровне. Все это свидетельствует о жестком контроле признака со стороны генотипа и высокой степени приспособленности к паразитическому образу жизни данных видов повиликовых, которая даже в пределах рода может достаточно широко варьировать: от присутствия хлорофилла у одних видов до полного отсутствия какой – либо фотосинтетической активности у других. Кроме того, проведенные исследования позволяют утверждать и отсутствие зависимости числа твердых семян с их формой, окраской рубчика и качеством их оболочки [4], как это показано для других видов растений.
В литературе есть сведения, что незрелые семена повилики китайской (Ciscuta chinensis Lam.) прорастают быстрее в отличие от зрелых [13]. Показано, что семена обладают гетерогенностью и даже у одного растения будут отличаться по степени зрелости. Мы исследовали способность к прорастанию повилики японской у семян разных групп: «зеленые», «незрелые» – сформированы после созревания в коробочках в течение 7 дней; «зрелые» темно-окрашенные – длина в среднем 2 мм). Наши наблюдения показали, что процент прорастания был выше у «незрелых» семян и составлял 80±3% по сравнению со «зрелыми» семенами - 3±1%. Зеленые семена прорастали плохо, однако процент прорастания их был несколько выше, чем у зрелых и составлял 10±1%. Продолжительность жизни проростка также была неодинакова для различных групп. Группа «зеленых» семян характеризовалась очень низкой продолжительностью жизни проростка (3-5 дней), что связано с недостатком питательных веществ и недоразвитием зародыша. Тогда как у зрелых семян про- росток способен выживать без прикрепления к растению-хозяину, используя только собственные резервы, достаточно длительное время в течение 24-27 дней.
После 14 дней высыхания все семена становятся зрелыми и процент прорастания снижается до 3±1%. Кроме того, мы наблюдали за семенами, находящимися во влажной камере при нормальных температурных условиях в течение 2-х лет. Установлено, что процент прорастания с течением времени не увеличивался, а оставался на уровне 3-5%. Это еще раз говорит в пользу гетерогенности семян повилики японской. При этом семена не утрачивают своей жизнеспособности и быстро прорастают после химической скарификации.
Эколого-географические условия формирования вида, как правило, детерминируют твердосемянность, в частности, низкая влажность воздуха ведет к резкому увеличению процента твердых семян [4]. Однако, наши данные для C. japonica равнозначны данным японских исследователей [13], несмотря на то, что условия формирования семян резко различаются по данному фактору. Все это, в очередной раз, свидетельствует об узком диапазоне варьирования признака и строгом генетическом контроле.
В ходе нашего эксперимента определен и целесообразный метод для ускорения прорастания семян повиликовых – обработка семян концентрированной серной кислотой, позволяющей получить проростки данных видов в массе для последующего их изучения, так как исследование их ювенильного этапа затруднено, в связи с твердосемянностью. По сравнению с контролем (3%) процент прорастания C. japonica при обработке H 2 SO 4 конц. (время воздействия 1 час) достигал стабильно 70%. Для прорастания повилики полевой требовалось меньшее время обработки 40-45 минут: процент прорастания при этом уже составлял 83%, а в течение 1 часа – 89-90 % (контроль 14% (рис. 1). Разница в реакции на обработку семян, вероятно, обусловлена их размером ( C. japonica – 2-3 мм, C. campestris 11,5 мм) и видовыми различиями.
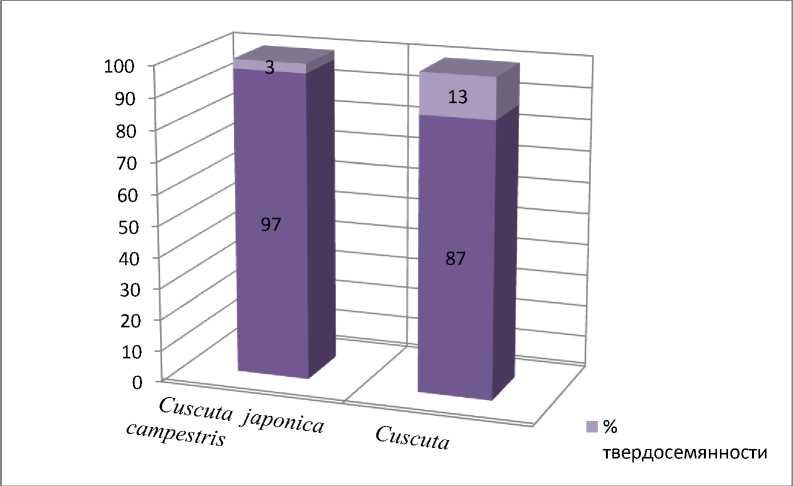
Рис. 1. Особенности прорастания семян Cuscuta japonica Choisy и C. campestris Yunck. (показано среднее значение признака)
Прорастание семян и прикрепление растений – паразитов к хозяину происходит под действием химических сигналов – корневых выделений некоторых растений [9, 10]. Это экспериментально установлено для растений – корневых паразитов. Поэтому считают, что облигатный стеблевой паразит – повилика Cuscuta L. имеет особенности подобные другим паразитическим растениям, и семена повилик ускоренно прорастают, реагируя на выделения растений-хозяев. Стимуляция прорастания семян паразита корневыми выделениями других растений обеспечивает ему определенные преимущества, так как повышают вероятность успешного заражения. Нами экспериментально показано, что присутствие семян и проростков растений-хозяев: Capsicum annuum L., Brassica oleracea L., Helianthus annus L., Cucumis sativus L., Raphanus sativus L., Avena sativa L. не влияет на прорастание семян повилики японской C. japonica. Это может быть обусловлено полной водонепроницаемостью семенной оболочки по-вилик. Таким образом, утверждение о влиянии растений-хозяев на прорастание не является всеобщим, по крайней мере, для повилики японской.
Заключение
В ходе наших исследований определен целесообразный метод для ускорения прорастания семян повиликовых – обработка семян концентрированной серной кислотой для Cuscuta japonica оптимальное время обработки составляет 60 минут, а для C. campestris – 40-45 минут. Более широкий размах признака повилики полевой, позволяет сделать вывод об иной стратегии вида, стремящегося занять свой потенциальный ареал. Возможно, это связано с космополитизмом C. campestris , как более универсального паразита, способного существовать в самых разнообразных условиях обитания. В отличие от нее, повиликой японской (аборигенным видом) уже выработан адаптивный механизм обитания в данных условиях. Семена повилики японской обладают гетерогенностью. Утверждение о влиянии растений-хозяев на прорастание не является всеобщим, по крайней мере, для повилики японской.
Список литературы Особенности твердосемянности Cuscuta japonica Choisy, C. campestris Yunck. (Cuscutaceae Dum.)
- Городков К.Б. Динамика ареала: общий подход. III. динамика ареала под воздействием биотических факторов // Энтомологическое обозрение. - 1997. - Т. 76 (1). - С. 86-110.
- Аистова Е.В., Леусова Н.Ю. Растения-хозяева видов рода Cuscuta l. (Cuscutaceae Dumort.) в Амурской области и в Приморском крае // Turczaninowia. - 2012. - Т. 15, № 2. - С. 96-101.
- Бейлин И.Г. Паразитизм и эпифитотиология. - М.: Наука, 1986. - 351 с.
- Николаева М.Г., Разумов М.В., Гладкова В.Н. Справочник по проращиванию покоящихся семян. - Ленинград: Наука, 1985. - 347 с.
- Леусова Н.Ю., Крылов А.В. Покой семян повилики японской Cuscuta japonica Choisy // VII Дальневосточная молодежная школа-конф. по актуальным вопросам химии и биологии, 15-22 сент., 2003 г., Владивосток: тез. докл. - Владивосток: МЭС, ТИБОХ ДВО РАН. - С. 32-33.
