Особенности видового состава и показатели зараженности гельминтами озерной лягушки Pelophylax ridibundus в условиях трансформации Местообитаний южного Урала

Автор: Зарипова Фалия Фуатовна, Файзулин Александр Ильдусович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 5-5 т.16, 2014 года.
Бесплатный доступ
В статье приведены данные по видовому составу, показателям зараженности гельминтов озерной лягушки Южного Урала (Оренбургской области и Республики Башкортостан) в условиях разнохарактерного антропогенного воздействия. Таксономический состав гельминтов включает для района исследования 16 видов: 13 видов трематод и 3 вида нематод. Выявлено снижение показателей экстенсивности инвазии в условиях наибольшего антропогенного воздействия у 4 видов трематод (на статистически значимом уровне).
Амфибии, гельминты, антропогенные воздействия, оренбургская область, республика башкортостан
Короткий адрес: https://sciup.org/148203450
IDR: 148203450 | УДК: 597.6
Features of species composition and indicators of infection by helminths marsh frog Pelophylax ridibundus in the context of transformation of the habitats of the Southern Urals
The article presents data on species composition, infection rates of helminths Marsh Frog of Southern Urals (Orenburg region and the Republic of Bashkortostan) in a diverse human impact. The taxonomic composition of worms for the study area includes 16 species: 13 species of trematodes and 3 species of nematodes. Showed a reduction in performance in terms of extensiveness of invasion largest anthropogenic impact in 3 species of trematodes (at a statistically significant level).
Текст научной статьи Особенности видового состава и показатели зараженности гельминтами озерной лягушки Pelophylax ridibundus в условиях трансформации Местообитаний южного Урала
В настоящее время антропогенная трансформация существенно влияет как на популяционную структуру, так и на биоценотические связи животных [1, 3, 8–11, 14, 15].
Следствием влияния антропогенного фактора является изменение видового состава животных и структуры их популяций, в том числе бесхвостых амфибий. Наша работа посвящена изучению некоторых популяционных характеристик озерной лягушки Pelophylax ridibundus Pallas, 1771 на территории Южного Урала – востока Оренбургской области и юго-востока Республики Башкортостан.
МАТЕРИАЛ И МЕТОДИКА
Сбор и обработка материала произведена в 2012-2014 гг. в 4-х популяциях озерной лягушки (локалитетах) (табл. 1): 1. «Худолаз» – окрестности г. Сибай, Баймакский район Республики Башкортостан, р. Худолаз, городская плотина (n=15); 2. «Гадельшинские озера» – пруды в 18 км на северо-запад от г. Сибай, Баймакский район Республики Башкортостан (n=20); 3. «Ишкинино» – д. Ишкинино, Гайский район Оренбургской области, р. Сухая Губерля (n=15); 4. «Уртазым» – прибрежное мелководье Ириклинского вдхр. на р. Урал у д. Уртазым Кваркенского района Оренбургской области (n= 20).
Несмотря на кратное повышение ПДК в лока-литете № 2 «Гадельшинские озера» нами данное местообитание рассматривается в качестве контроля, при этом высокие показатели содержания меди и цинка отражают фоновое значение для водоемов территорий Башкирского Зауралья.
В качестве условного контроля нами выбрана популяция «Уртазым» Ириклинского водохрани-
лища. Анализ качества воды показал превышение ПДК по меди – в 4 раза, по ХПК – в 1,5 ПДК, БПК5 – в 1,2 раза, нефтепродукты на уровне ПДК, ниже ПДК азот нитритный.
Методом полных гельминтологических вскрытий изучались паразиты амфибий [13]. Определение гельминтов амфибий проводили по книге К.М. Рыжикова с соавторами [12]. Для анализа зараженности амфибий использовали показатели: экстенсивность (Е, %), интенсивность (I, экз.) инвазии, индекс обилия (М, экз.) паразитов [1]. Статистическая оценка различий по экстенсивности инвазии проведена по принятой методике. В соответствии со значениями экстенсивности инвазии условно выделяются следующие группы паразитов: доминантные (Е>70%), субдоминантные (Е>50%), обычные (Е>30%), редкие (Е>10%) и единичные (Е<10%).
Проведенный таксономический анализ показал, что на территории Оренбургской области популяции озерной лягушки P. ridibundus представлены «восточной» формой по данным анализа митохондриальной ДНК [5, 20].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Общий состав гельминтов озерной лягушки района исследования включает 16 видов. Это трематоды – Gorgodera loossi (Sinitzin, 1905), Gorgoderina vitelliloba (Olsson, 1876), Opisthiogly-phe ranae (Frölich, 1791) Looss, 1899, Pneumonoe-ces variegatus (Rudolphi, 1819), Skrjabinoeces simi-lis (Looss, 1899) Sudarikov, 1950, Pleurogenes claviger (Rudolphi, 1819) Looss, 1896, Pleuro-genoides medians (Olsson, 1876) Travassos, 1921, Pleurogenes intermedius Issaitchikov, 1926, Proso-tocus confusus (Looss, 1894) Looss, 1899, Brandesia turgida (Brandes, 1888) Stossich, 1899, Strigea fal-conis Szidat, 1928, mtc., Strigea strigis (Schrank, 1788) Abildgaard, 1790, mtc., Codonocephalus urni-gerus (Rudolphi, 1819) Diesing, 1850, mtc.; немато- ды – Rhabdias bufonis (Schrank, 1788), Cosmocerca ornata (Dujardin, 1845), Aplectana acuminata (Schrank, 1788).
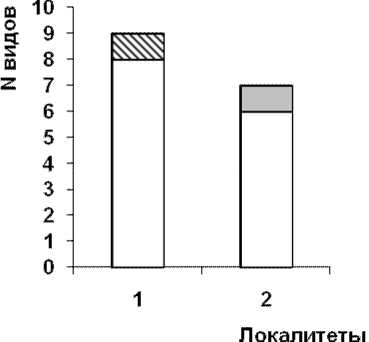
Общими для всех трех локалитетов являются 2 вида O. ranae, P. variegatus . В локалитете №1 доминантные по степени экстенсивности – P. claviger , субдоминантые – G. loossi ,обычные – O. ranae , P. variegatus , редкие – P. medians , P. con-fusus, B . turgida, A. acuminata , единичные – G. vitelliloba . В локалитета №2 доминантные – G. loossi , O. ranae , субдоминантые – P. confusus , обычные – P. variegatus, S. falconis, mtc., редкие – S. similis, P . medians, единичные не выявлены.
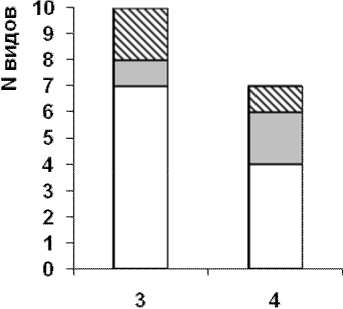
Для локалитета №3 доминантные виды отсутствуют, субдоминантые – O. ranae, обычные – P. variegatus, P. claviger, R. bufonis, A. acuminata, редкие – S. similis, P. intermedius, S. falconis, mtc., единичные – G. loossi, G. vitelliloba. Для локали-тета №4 доминантные – O. ranae, C. ornata, суб-доминантые – не выявлены, обычные – P. con-fusus, редкие – P. claviger, P. variegatus, S. strigis, mtc., Codonocephalus urnigerus, mtc., единичные не выявлены.
Экологический анализ состава гельминтов представлен на рис. 1.
В условиях высокой нагрузки загрязнения местообитания отсутствуют гельминты, передающиеся от амфибий к хищникам-батрахофагам (аллогенные биогельминты) - S. falconis, mtc., Strigea strigis, mtc., Codonocephalus urnigerus, mtc..
Таблица 1. Данные анализа воды из местообитаний амфибий по содержанию эссенциальных и токсических металлов
|
s § s ц н |
сч д s s Н U £ |
Медь (Cu) |
Цинк (Zn) |
Кадмий (Cd) |
Свинец (Pb) |
||||
|
мг/дм3 |
пдк рыбх. |
мг/дм3 |
пдк рыбх. |
мг/дм3 |
пдк рыбх. |
мг/дм3 |
пдк рыбх. |
||
|
1 |
высокая |
0,0213 |
21,30 |
0,3790 |
37,90 |
0,0036 |
0,72 |
0,0001 |
0,02 |
|
2 |
низкая |
0,0110 |
11,00 |
0,0390 |
3,90 |
0,0010 |
0,20 |
0,0020 |
0,33 |
|
3 |
высокая |
0,0090 |
9,00 |
0,0410 |
4,10 |
0,0010 |
0,20 |
0,0140 |
2,33 |
|
4 |
низкая |
0,003 |
3,00 |
0,04 |
4,00 |
- |
Ниже ПДК |
- |
Ниже ПДК |
Таблица 2. Состав гельминтов популяций P. ridibundus Южного Урала
|
Виды гельминтов |
Локалитеты |
|||
|
Республика Башкортостан |
Оренбургская область |
|||
|
1. Худолаз |
2. Гадельшинские озера |
3. Ишкинино |
4. Уртазым |
|
|
Высокая |
Низкая |
Высокая |
Низкая |
|
|
1 |
2 |
3 |
4 |
5 |
|
G. loossi * |
66,67±12,18(1-13) 2,53 |
100,00±0,00(4-96) 20,50 |
6,67±6,45(1) 0,06 |
– |
|
G. vitelliloba* |
6,67±6,45(1) 0,06 |
– |
6,67±6,45(1) 0,06 |
– |
|
O. ranae** |
44,34±11,46(1-16) 1,73 |
95,00±4,87(8-481) 68,00 |
53,33±12,89(21-144) 41,87 |
90,00±6,71(1-25) 9,00 |
|
P. variegatus*** |
40,00±12,66(1-8) 1,47 |
30,00±10,25(1-41) 4,75 |
40,00±12,66(1-9) 1,80 |
25,00±0,31(1-7) 0,75 |
|
S. similis *** |
– |
10,0±6,71(1) 0,10 |
20,00±10,34(2-7) 0,80 |
– |
|
P. claviger ** |
80,00±10,34(1-11) 4,07 |
– |
46,67±12,89(3-9) 2,47 |
10,00±6,71(2-3) 0,25 |
|
P . medians** |
20,00±10,34(1-4) 0,60 |
10,00±2,24(6-7) 0,65 |
– |
– |
|
P. intermedius** |
– |
– |
13,33±8,79(3-5) 0,53 |
– |
|
P. confusus** |
13,33±33,98(1-7) 0,53 |
60,00±13,42(1-12) 2,90 |
– |
40,00±10,96(2-15) 2,30 |
|
B . turgida |
26,67±11,43(1-26) 2,33 |
– |
– |
– |
|
S. falconis, mtc.**** |
– |
45,00±11,13(2-15) 3,35 |
33,33±12,18(4-17) 3,4 |
– |
|
S. strigis, mtc.**** |
– |
– |
– |
15,00±7,99(3-14) 1,05 |
|
C. urnigerus, mtc.**** |
– |
– |
– |
25,00±10,96(12) 0,60 |
|
R. bufonis*** |
– |
– |
40,00±12,66(1-6) 1,13 |
Окончание таблицы 2
|
1 |
2 |
3 |
4 |
5 |
|
C. ornata** |
– |
– |
– |
85,00±7,99(1-36) 8,45 |
|
A. acuminata** |
13,30±8,77(1) 0,13 |
– |
33,33±12,18(3-6) 1,4 |
– |
|
Нематод |
1 |
0 |
2 |
1 |
|
Трематод |
8 |
7 |
8 |
6 |
|
Число видов |
9 |
7 |
10 |
7 |
Примечание: над чертой – экстенсивность заражения (Е±mE, %); в скобках – интенсивность заражения (I, экз.); под чертой – индекс обилия паразита (M, экз.). Локализация: * – мочевой пузырь, ** – кишечник, *** – легкие, **** – кожа
В локалитете № 2 при низкой нагрузке отсутствуют гельминты амфибий – автогенные геогельминты (не циркулирующие по трофическим связям), отмеченные в других локалитетах – R. bufonis , C. ornata , A. acuminata .
С возрастанием степени трансформации местообитания снижается число видов гельминтов, поступающих через объекты питания амфибий (автогенные биогельминты) в ряду 8 – 7 – 6 – 4. Данная группа включает 10 видов – G. loossi, G. vitelliloba, O. ranae, P. variegatus, S. similis, P. claviger, P. medians, P. intermedius, P. confusus, B. turgida.
-
□ автогенные геогельминты
-
□ аллогенные биогельминты
-
□ автогенные биогельминты
Локалитеты
а)
б)
Рис. Экологические формы гельминтов P. ridibundus а) Республика Башкортостан, локалитеты: 1. Худолаз, 2. Гадельшинские озера; б) Оренбургская область: 3. Ишкинино 4.Уртазым
Для 3 видов трематод уровень экстенсивности ниже на статистически значимом уровне в условиях загрязнения местообитания земноводных Республики Башкортостан G. loossi (р<0,05), O. ranae (р<0,001) и P. confusus (<0,01). В Оренбургской области экстенсивность инвазии у 2 видов трематод на статистически значимом уровне O. ranae (р<0,05) ниже, а P. claviger (р<0,05) выше в условиях загрязнения.
В целом фауна гельминтов характерна для Южного Урала, по данным ранее проведенных исследований [4, 6, 7, 18, 19].
По данным [15] в условиях антропогенной трансформации – урбанизации –происходит разрушение исторически сложившихся паразитарных систем в условиях низкой численности или вымирания видов – окончательных хозяев гельминтов (ужей, хищных птиц, псовых, млекопитающих). Другим фактором нарушения устойчивости паразитарных систем является снижение доли в рационе и, соответственно, в трансформированных экосистемах промежуточных и/или дополнительных хозяев (моллюсков, личинок и имаго стрекоз, жуков, ручейников, головастиков и сеголетков амфибий). При этом обычно видовой состав гельминтов сохраняется, но снижаются показатели зараженности гельминтами. По мнению А.А. Бураковой [2] R. bufonis «характеризуется высокой устойчивостью к химическому загрязнению». Причем в целом трематоды в большем числе встречаются в «экологически чистых местностях». При этом нематода О. filiformis от- мечена с высокими показателями зараженности у остромордой лягушки городских территорий [2]. В условиях сильного загрязнения пестицидами отмечается снижение (на статистически значимом уровне) экстенсивности и интенсивности инвазии у 3 видов трематод G. varsoviensis, P. claviger, S. similis и возрастание у трематоды O. ranae при снижении интенсивности инвазии [8]. Для г. Тольятти [13] также отмечается снижение показателей экстенсивности и интенсивности инвазии у 13 видов трематод Gorgodera asiatica, G. pagen-stecheri, G. varsoviensis, Gorgoderina vitelliloba, Pneumonoeces variegatus, Skrjabinoeces similis, Brandesia turgida, Prosotocus confusus, Pleurogenes claviger, Pleurogenoides medians, Opisthioglyphe ranae, ad., Paralepoderma cloacicola, mtc., Strigea strigis, mtc. и 1 вида нематод Cosmocerca ornata, за исключением трематоды P. asper. По данным для района г. Уфа [6] отмечено снижение экстенсивности инвазии 3-х видов трематод Opisthiogly-phe ranae, Pleurogenes claviger, Strigea sphaerula, mtc. Для города Ульяновска [9] с градиентом антропогенного воздействия видовой состав возрастает с 8 и 9 до 13 видов при снижении экстенсивности инвазии.
ВЫВОДЫ
-
1) Видовой состав гельминтов (число видов) не отражает напрямую степень антропогенного воздействия на местообитания амфибий, которое не является однородным и проявляется как в загрязнении водоемов (в районе исследования – тяжелыми металлами), так и в трансформации (создание водохранилищ, прудов).
-
2) В условиях градиента антропогенного воздействия от низкого к высокому, возрастает экстенсивность инвазии гельминта P. variegatus , а вида O. ranae - снижается.
-
3) Большое значение имеют изменения в составе экологических форм гельминтов: в условиях трансформации местообитания отсутствуют гельминты II-й экологической группы, передающиеся от амфибий к хищникам-батрахофагам (аллогенные биогельминты).
Исследование проведено при поддержке гранта РФФИ № 14–04–31315 мол_а.
Список литературы Особенности видового состава и показатели зараженности гельминтами озерной лягушки Pelophylax ridibundus в условиях трансформации Местообитаний южного Урала
- Бреев К.А. Применение негативного биноминального распределения для изучения популяционной экологии паразитов. Методы паразитологических исследований. Л.: Наука, 1972. 70 с.
- Буракова А.В. Особенности заражения гельминтами остромордой лягушки фоновых и урбанизированных территорий//Вестник Оренбургского государственного университета. 2008. № 2. С. 111-116.
- Вершинин В.Л. Экологические особенности популяций амфибий урбанизированных территорий: Автореф. дис. … д-ра биол. наук. Екатеринбург, 1997. 47 с.
- Давлетбакова Г.М., Юмагулова Г.Р. Гельминты бесхвостых амфибий Оренбургской области//Экологический сборник 4. Труды молодых ученых Поволжья/под ред. проф. С.В. Саксонова. Тольятти: Кассандра, 2013 -С. 31-34.
- Ермаков О.А., Файзулин А.И., Закс М.М., Кайбелева Э.И., Зарипова Ф.Ф. Распространение «западной» и «восточной» форм озерной лягушки Pelophylax ridibundus s. l. на территории Самарской и Саратовской областей (по данным анализа митохондриальной и ядерной ДНК)//Известия Самарского научного центра Российской академии наук. 2014. Т. 16, № 5(1). С. 409-412.
- Зарипова Ф.Ф., Юмагулова Г.Р., Файзулин А.И. Гельминтофауна озёрной лягушки (Rana ridibunda Pallas, 1771) урбанизированных территорий республики Башкортостан//Современная герпетология. 2012. Том 12, вып. 3/4. С. 134-142.
- Зарипова Ф.Ф., Юмагулова Г.Р., Файзулин А.И. Гельминты озерной лягушки Rana ridibunda (Pallas, 1771) г. Салават (Республика Башкортостан)//Современные проблемы общей паразитологии. Материалы Международной научной конференции (30 октября -1 ноября 2012 г. Москва): Центр паразитологии Института проблем экологии и эволюции им. А.Н. Северцова РАН, 2012. С. 127-130.
- Лебединский А.А. Гельминтофауна озерных лягушек в условиях антропогенного воздействия и связь инвазии с полиморфизмом//Животные в природных экосистемах. Н. Новгород, 1994. С. 25-32.
- Матвеева Е.А. Эколого-фаунистические особенности гельминтофауны Rana ridibunda Pall. на территории Ульяновской области: автореф. дис. …канд. биол. наук. -Ульяновск, 2009. -24 с.
- Резванцева М.В. Материалы по гельминтофауне озерной лягушки (Rana ridibunda) в окрестностях Тамбова//Вестник Тамбовского ун-та. Сер. Естественные и технические науки. 2008. Т. 13, Вып. 5. С. 330-332.
- Романова Е.М., Индирякова Т.А., Матвеева Е.А. Паразитарные системы как индикатор состояния биоценоза//Вестник Ульяновской государственной сельскохозяйственной академии. 2009. № 2. С. 79-81.
- Рыжиков К.М., Шарпило В.П., Шевченко Н.Н. Гельминты амфибий фауны СССР. М.: Наука, 1980. 279 с.
- Скрябин К.И. Метод полных гельминтологических вскрытий позвоночных, включая человека. М.: Изд-во МГУ, 1928. 45 с.
- Файзулин А.И., Чихляев И.В., Кузовенко А.Е. Амфибии Самарской области. Кассандра, 2013. 140 с.
- Чихляев И.В., Файзулин А.И., Замалетдинов Р.И., Кузовенко А.Е. Трофические связи и гельминтофауна зеленых лягушек Rana esculenta complex (Anura, Amphibia) урбанизированных территорий Волжского бассейна//Працi Українського герпетологiчного товариства, 2009, N. 2. С. 102-109.
- Чихляев И.В. Характеристика жизненных циклов гельминтов земноводных (Amphibia) Среднего Поволжья//Биоразнообразие и экология паразитов: Тр. Центра паразитол. 2010. Т 46. С. 315-324.
- Чихляев И.В., Кириллова Н.Ю., Кириллов А.А. Характеристика жизненных циклов трематод (Trematoda) наземных позвоночных Среднего Поволжья//Известия Самарского научного центра Российской академии наук. 2012. Т. 14. № 5-1. С. 132.
- Чихляев И.В., Файзулин А.И. Использование гельминтологических данных при исследовании формирования ареалов бесхвостных земноводных (Anura, Amphibia) Волжского бассейна//Теоретические проблемы экологии и эволюции. Теория ареалов: виды, сообщества, экосистемы (V Любищевские чтения)/Под ред. чл.-корр. Г.С. Розенберга и проф. С.В. Саксонова. Тольятти: ИЭВБ РАН, 2010. С. 209-214.
- Юмагулова Г.Р. Гельминты амфибий Южного Урала: Автореф. дис. … канд. биол. наук. Уфа, 2000. 19 с.
- Akın C., Bilgin C.C., Beerli P., Westaway R., Ohst T., Litvinchuk S.N., Uzzell T., Bilgin M., Hotz H., Guex G.-D. Phylogeographic patterns of genetic diversity in eastern Mediterranean water frogs have been determined by geological processes and climate change in the Late Cenozoic//J. Biogeogr. 2010. V. 37. P. 2111-2124.
