Особенности влагопотребления и влагообеспеченности растений различных экологических групп

Автор: Муромцев Н.А., Семенов Н.А., Анисимов К.Б.
Журнал: Бюллетень Почвенного института им. В.В. Докучаева @byulleten-esoil
Статья в выпуске: 82, 2016 года.
Бесплатный доступ
Каждая экологическая группа растений (гигрофиты, мезофиты и ксерофиты) характеризуется определенным, свойственным только ей, узким интервалом потенциала почвенной влаги, при котором значения относительной транспирации не опускаются ниже оптимального уровня (1.0-0.9). Эти интервалы учитывают как влияние свойств почв (через потенциал влаги), так и метеорологические условия (через транспирацию) и могут быть использованы в качестве оптимального показателя влагообеспеченности растений. Выявлены особенности потребления почвенной влаги растениями, заключающиеся в том, что объем почвенной влаги и интенсивность ее потребления увеличиваются, а “мертвый” запас уменьшается в направлении от гигрофитов к мезофитам и далее к ксерофитам. Показано, что снижение роста и накопление полезной продукции растениями проявляется уже при уменьшении содержания почвенной влаги до нижней границы оптимального увлажнения. Дальнейшее уменьшение влажности почвы приводит сначала к перманентному, а затем и устойчивому завяданию растений. Установлены интервалы значений потенциала почвенной влаги, соответствующие влажности устойчивого завядания растений разных экологических групп. Салат (гигрофит) погибает при содержании влаги, равном 1.5, овес (мезофит) - при 1.0, а ксерофитные растения - ниже 1.0 влажности завядания. Широко используемый метод “вегетационных миниатюр” является несовершенным и может быть использован лишь для ориентировочных представлений о нижнем пределе доступности почвенной влаги растениям.
Транспирация, испаряемость, потенциал почвенной влаги, градиент потенциала, влажность почвы
Короткий адрес: https://sciup.org/14313652
IDR: 14313652 | УДК: 631.4 | DOI: 10.19047/0136-1694-2016-82-71-87
Peculiarities in water use and supply of different ecological plant groups
Every ecological group of plants including hygrophytes, mesophytes and xerophytes is characterized by a definite narrow interval of the soil water potential, in which the values of relative transpiration cannot be lower than the optimal level (1.0-0.9). These intervals take into account the influence of soil properties (moisture potential) and meteorological conditions (transpiration) and can be used as optimal indices for the water supply of plants. The peculiar soil water uptake by plants has been studied to show that the soil water and its intensive uptake become increasing but its “dead” reserve decreasing from hygrophytes towards mesophytes and then xerophytes. It is worth emphasizing that the decline in the growth and accumulation of useful products by plants occur due to decreasing the content of soil water to the lower boundary of optimal moistening. The further decrease in soil moisture leads to permanent and then ultimate wilting of plants. Under study were also intervals for soil water potential corresponding to the moisture of ultimate wilting of different ecological plant groups. Lettuce (hygrophyte) is becoming dead when the content of soil water is equal to 1.5, oat (mezophyte) - 1.0 and xerophytes - lower than 1.0 of the wilting moisture. The method of “vegetative miniatures” widely adopted now is imperfect and can be performed only for preliminary notions about the lower limit for plant availability of the soil water.
Текст научной статьи Особенности влагопотребления и влагообеспеченности растений различных экологических групп
Растению для поддержания нормальной жизнедеятельности необходим определенный уровень транспирации, назовем ее абсолютной транспирацией ( АТ ), соответствующий испаряемости ( Е 0 ), т.е. АТ = Е 0 . Отсюда, поток влаги из почвы в растение должен удовлетворять потенциально возможной транспирации (Т 0 ), соответствующей (равной) испаряемости, т.е. поток влаги ( I ) =АТ= Т 0. = Е 0 . Однако из-за несоответствия скорости потока влаги через границу почва–корень растения скорости ее потока на границе лист расте-ния–атмосфера в природных условиях поток влаги ( I ) из почвы в растение часто становится меньше относительной транспирации, равной испаряемости (Т 0 = Е 0 ) (Семенов и др., 2005; Муромцев, 2011).
Для достижения такой высокой интенсивности АТ необходимо обеспечить соответствующую плотность потока влаги из почвы в растение. Однако плотность потока влаги в системе почва–растение определяется не только ее содержанием в почве, но и сопротивлением проводящей системы твердая фаза почвы– паренхима–сосуды корня, стебля и листьев. В результате происходит несоответствие между поступлением влаги из почвы в растение и расходованием ее на транспирацию, что приводит к возникновению водного дефицита влаги в листьях, к перманентному их завяданию и снижению темпов наращивания полезной продукции. И это несоответствие существенно зависит от биологофизиологических свойств, которые сильно различаются между растениями, представителями различных экологических групп: гигрофитов, мезофитов и ксерофитов. К первой группе относятся влаголюбивые овощные культуры (салат, капуста, огурцы и другие), к мезофитам – хлебные злаки и многие виды трав, а к ксерофитам – дикорастущие растения, например, житняк, дикая вишня и др. К последним культурам можно отнести и засухоустойчивые виды некоторых культурных злаков, например, просо и пшеницу Саратовская 29, отличающуюся повышенной степенью засухоустойчивости по сравнению с другими сортами.
Для того чтобы иметь реальную и адекватно отражающую состояние влаги в почве и растении информацию необходим динамический критерий влагообеспеченности растений. В качестве такого критерия часто используется относительная транспирация (Т/Т0), представляющая отношение действительной (Т) транспирации к потенциальной (Т0). Она имеет преимущество по сравнению с другими подобными критериями, так как учитывает и изменение метеорологической обстановки, и физиологические особенности растений (Муромцев, 2002).
Все живительные соки природы (вода и растворенные в ней химические вещества) передвигаются как на поверхности и в теле земли, так и в отдельно взятом растении, исключительно благодаря градиенту (перепаду) сил: осмотических, гидравлических, гравитационных, механических и других (Глобус, 1969; Судницын, 1979; Мелиоративная энциклопедия, 2004; Hosseini et al., 2011). Градиенты сил, под действием которых движутся все субстанции (свет, тепло, влага, электромагнитные поля и др.) представляет собой перепад сил на единицу расстояния. Если силы любого вида в двух произвольно взятых точках пространства (дневной поверхности земли, растения и прочего) будут равны, то движения веществ и энергий в этом направлении происходить не будет. Это означает, что растения не смогут потреблять влагу и химические вещества из почвы, и неминуемо погибнут.
Градиент (перепад) потенциала влаги обеспечивает как внутрипочвенный влагообмен, так и обмен влагой и элементами питания растений в системе почва–растение–атмосфера (Муромцев, 2005; Воронин, 1984; Муромцев и др., 2014; Pertovici et al., 2010). Возникновение перепада потенциала происходит как в системе почва–всасывающие корни, так и между потенциалом влаги в корнях и листьях растений.
В соответствии с изложенными представлениями состояние воды в системе почва–растение–атмосфера можно представить в виде непрерывной водной нити (водного тяжа), идущей непосредственно из слоя почвы, прилегающего к корневым волоскам, затем переходящей в корни, от них в стебель и далее в листья. Этот водный тяж находится в непрерывном и постоянном (в вертикальном и горизонтальном направлении) движении. Первопричиной движения является расход влаги из системы на испарение ее через устьица растения в атмосферу и за счет поступления влаги из почвы. Происходит наращивание водной нити снизу при расходе ее сверху. Скорость этого движения определяется интенсивностью
Бюллетень Почвенного института им. В.В. Докучаева. 2016. Вып. 82. транспирации, и существенно зависит от влагопроводности почвы (Муромцев, 2013).
ОБЪЕКТЫ И МЕТОДЫ
Исследования проведены на аллювиальных почвах р. Арчеда (Волгоградская область) и р. Москва (Московская область). В прирусловых частях рек Арчеда и Москва расположены аллювиальные супесчаные почвы, а в центральных частях пойм – их суглинистые аналоги. Морфогенетические и агрофизические свойства этих почв кратко рассмотрены в ранее (Муромцев, 2011). Дополнительно укажем, что величины основных водно-физических свойств аллювиальных суглинистых почв близки между собой. В частности, объемная масса – верхнего слоя 0–30 см составляет 1.1–1.2 г/см3, влажность завядания – 8.0–10.0%, наименьшая влагоемкость 26–28%, содержание гумуса 2.48–2.81%. Аллювиальная супесчаная почва (пойма р. Арчеда) в слое 0–30 см характеризуется величинами объемной массы – 1.45 г/см3, влажности завядания – 2.72%, наименьшей влагоемкости – 11.4% и содержания гумуса – 1.34%.
Исследования выполняли в лабораторных и полевых условиях с использованием больших (около 9 кг почвы) вегетационных сосудов и испарителей ГГИ-500-100 высотой 100 см и диаметром 40 см. В лабораторных условиях в вегетационных сосудах с аллювиальной суглинистой и супесчаной почвами выращивали салат (гигрофит), овес (типичный мезофит), засухоустойчивые сорта пшеницы (Саратовская 29) и проса и типичный ксерофит житняк. Эти же культуры выращивали в испарителях, установленных на лизиметрической площадке в пойме р. Москва. В вегетационных сосудах и испарителях устанавливали тензиометры для измерения капиллярного потенциала влаги. Полный потенциал влаги в листьях и почвах измеряли психрометрическим методом (Муромцев, 2011), а влажность почвы сосудов и испарителей – весовым методом.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Перепад потенциала влаги создается на границах перехода из одной части системы почва–растение–атмосфера в другую и в пределах каждой части. На границе перехода жидкости в пар, т.е. на границе системы лист–воздух, он обусловлен разностью между упругостью водяного пара в тканях листьев и в окружающем слое воздуха. На границе перехода влаги из почвы в корни – разностью между потенциалом в корнях растений и в прилегающем к ним слое почвы. Поскольку прилегающий к корням слой почвы постоянно дренируется (обезвоживается), то между ним и последующими за ним слоями почвы, в свою очередь, образуются перепады потенциала влаги. В самом растении так же, как и в отдельных его частях, перепады потенциала возникают из-за разной обводненности тканей.
Перепады потенциала влаги (движущая сила) в почве и опытных растениях, соотнесенные к потенциалу влаги в двух аллювиальных почвах, приведены в табл. 1.
Таблица 1. Соотношение перепада потенциала (Рл – Рп) влаги в системе почва–растение и потенциала почвенной влаги (Рп) в аллювиальных поч- вах поймы р. Арчеда (Волгоградская область)
|
Культура |
Перепад потенциала влаги, ×102 кПа |
|||
|
аллювиальная суглинистая почва |
аллювиальная супесчаная почва |
|||
|
Р п |
Р л – Р п |
Р п2 |
Р л – Р п |
|
|
Салат |
05 |
05 |
05 |
03 |
|
10 |
06 |
10 |
04 |
|
|
20 |
03 |
20 |
02 |
|
|
30 |
– |
30 |
– |
|
|
40 |
– |
40 |
– |
|
|
Овес |
05 |
10 |
05 |
10 |
|
10 |
12 |
10 |
12 |
|
|
20 |
13 |
20 |
13 |
|
|
30 |
06 |
30 |
12 |
|
|
40 |
05 |
40 |
– |
|
|
Просо |
05 |
10 |
05 |
11 |
|
10 |
18 |
10 |
18 |
|
|
20 |
23 |
20 |
21 |
|
|
30 |
20 |
30 |
20 |
|
|
40 |
17 |
40 |
17 |
|
|
Пшеница |
05 |
–1 |
05 |
11 |
|
10 |
24 |
10 |
27 |
|
|
20 |
27 |
20 |
33 |
|
|
30 |
33 |
30 |
– |
|
|
40 |
–330 |
40 |
– |
|
|
Житняк |
05 |
11 |
05 |
12 |
|
10 |
24 |
10 |
26 |
|
В соответствии с данными табл. 1, максимальные значения перепада потенциала влаги ( Р л – Р п ) в системе лист растения– почва в суглинистой почве составляют (6, 13, 23, 33) × 102 кПа соответственно в салате, овсе, просе и пшенице. При сравнительно высоком содержании влаги в почве, характеризуемом потенциалом влаги до 500 кПа (5 атм), перепады потенциала влаги минимальны: в салате – 5 × 102, овсе – 10 × 102, а в просе, пшенице и житняке – (10–13) × 102 кПа. По мере уменьшения содержания влаги и коэффициента влагопроводности ( К п) перепад потенциала плавно возрастает (по абсолютной величине) в салате и овсе, и довольно резко – в пшенице, просе и житняке, достигая в последних культурах значений в пределах (23–33) × 102 кПа. Характер изменения величин перепада потенциала таков: сначала он, возрастая, достигает максимального значения (различного для разных растений), затем с уменьшением влаги до влажности устойчивого завядания ( ВЗ ) растений (для ксерофитов и засухоустойчивых растений – до максимальной гигроскопической влаги ( МГ ) и ниже) снижается. При этом существенных различий между величинами перепада потенциала ксерофита и ксерофитных зерновых культур не наблюдается.
Конечные значения перепада потенциала, соответствующие состоянию устойчивого завядания растений, сильно различаются в растениях разных экологических групп. В соответствии с характером изменений Р п и перепада потенциала функция (Р л - P п ) f(P п ) имет одним максимумом и два минимумами. Физическая сущность этого явления может быть представлена следующим образом. В условиях высокого содержания влаги в почве около наименьшей влагоемкости ( НВ ) и немного ниже, поток в растение компенсирует ее расход на транспирацию. В это время усилия растений направлены в основном на преодоление сопротивления системы почва–ксилема стебля движению влаги, поскольку силы влагоудержания незначительны, и перепад потенциала минимален, но достаточен для обеспечения такой плотности потока, который компенсировал бы потери влаги на эвапотранспирацию (суммарное испарение).
При дальнейшем уменьшении содержания влаги в почве ее потенциал (Рп) понижается и тем интенсивнее, чем меньше влаги становится в почве. Это приводит к различной интенсивности увеличения потенциала влаги в растениях (Рл): она существенно выше в ксерофитных растениях по сравнению с мезофитами и гигрофитами. В результате этого перепад потенциала возрастает в системе почва–растение до максимума. Наконец, содержание влаги в почве становится настолько малым, что скорости повышения потенциала влаги в почве и растениях выравниваются. В этих условиях возрастание перепада потенциала влаги в системе почва–растение приостанавливается, в дальнейшем происходит его численное уменьшение.
Изложенные представления иллюстрируются рис. 1–3. При непрерывном уменьшении влажности почвы потенциал влаги в почве (рис. 1) и листьях растений (рис. 2) неуклонно повышается, однако Т/Т 0 некоторое время продолжает оставаться на уровне единицы, что свидетельствует о полной компенсации расхода влаги на транспирацию потоком ее из почвы. В дальнейшем, на 12-е сутки в суглинистой почве, относительная транспирация проса становится меньше единицы, и с этого времени она начинает быстро уменьшаться. Для других культур это уменьшение наблюдается значительно раньше, например, в салате в середине вторых суток.
По скорости (или времени) снижения Т/Т 0 до уровня <1.0– 0.9 растения можно расположить в ряд: салат – овес – (пшеница– просо–житняк), свидетельствующий о возрастании засухоустойчивости и о значительно больших возможностях потребления влаги от начала ряда к его концу. С этим вполне согласуется динамики влажности и потенциалов влаги в почве и растениях . В почве легкого гранулометрического состава изменения потенциалов влаги в почве и растениях, коэффициента влагопроводности (на рисунках не представлен) и относительной транспирации более динамичны, а снижение последней (Т/Т 0 ) наблюдается значительно раньше, чем в суглинистой почве, что обусловлено различием гидрофизических свойств этих почв (в частности, влагоемкости), главная роль в которых принадлежит водоудерживающим силам.
Начало снижения Т/Т 0 (рис. 1 и 2) в различных растениях наблюдается при следующих значениях потенциала почвенной влаги ( Р п ) соответственно в суглинистой и супесчаной почвах: 7.5 × 102 и 5.0 × 102 кПа в салате, 14.0 × 102 и 8.5 × 102 кПа в овсе, 17.0 × 102 и 14.0 × 102 кПа в просе, 11.0 × 102 и 9.5 × 102 кПа в пшенице и 16.0 × 102 кПа в житняке. Отсюда, наибольшее значе-
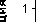
0,8
0,6
0,4
0,2

0 5 10 15 20 25 30 35 40
Потенциал влаги, кПа
Рис. 1. Зависимость относительной транспирации (Т/Т 0 ) различных растений от потенциала влаги в аллювиальной почве поймы р. Арчеда: суглинистая почва: 1 – пшеница, 3 – житняк, 4 – просо, 6 – овес, 8 – салат; супесчаная почва: 2 – пшеница, 5 – просо, 7 – овес, 9 – салат.
1,2
0,8
0,6
0,4
0,2
0 10 20 30 40 50 60 70
Потенциал влаги, кПа
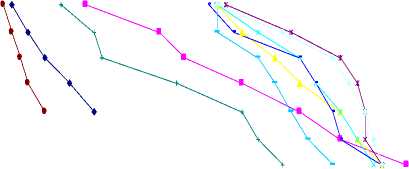
. . р рц 0р р ний от потенциала влаги в листьях; аллювиальная суглинистая почва: 1 – пшеница, 3 – житняк, 4 – просо, 6 – овес, 8 – салат; аллювиальная супесчаная почва: 2 – пшеница, 5 – просо, 7 – овес, 9 – салат, 10 – житняк.
ние Р п , соответствующее началу снижения Т/Т о , наблюдается при выращивании проса и житняка, а наименьшее – салата. Значение потенциала влаги в листьях всех исследованных растений значительно больше по сравнению с потенциалами влаги в почве. Оно
1,2
0,8
0,6
0,4
0,2

♦ 1
■—2
—*—5
—•—6
—I—7
--■--8
-^-9
15 20
Перепад потенциала влаги, кПа
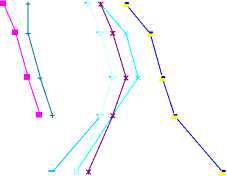
Рис. 3. Зависимость относительной транспирации (Т/Т 0 ) различных растений от перепада потенциала влаги ( Р л – Р п ) в аллювиальных почвах поймы р. Арчеда: суглинистая почва: 1 – пшеница, 3 – житняк, 4 – просо, 6 – овес, 8 – салат; супесчаная почва: 2 – пшеница, 5 – просо, 7 – овес, 9 – салат, 10 – житняк.
составляет для салата, овса, проса, пшеницы и житняка на суглинистой и супесчаной почвах: 12 × 102 и 10 × 102, 28 × 102 и 20 × 102, 38 × 102 и 37 × 102 и 44 × 102 и 43 × 102 кПа. Значения потенциала влаги в листьях овса и ксерофитных растений достигает внушительных величин: (55 – 58) × 102 кПа для растений на суглинистой и (43–55) × 102 кПа – супесчаной почвах.
Конечное значение Т/Т0, характеризующее устойчивое завядание растений ксерофитов, составляет 0.18–0.20, а салата – 0.47. Эти данные свидетельствует о том, что у гигрофитов (салата) влажность завядания наступает не постепенно (плавно), а резко с высокой интенсивностью. С физической точки зрения это можно объяснить тем, что у гигрофитов из-за высокой требовательности к воде не выработались в процессе эволюции соответствующие реакции на экстремальные условия среды, выражающиеся (в данном случае) в резком снижении доступности почвенной влаги. Поэтому потребление влаги этими растениями быстро прекращается при исчерпании потенциальных возможностей, характеризуемых отсутствием перепада потенциала, когда значения потенциала в листьях и почве примерно равны между собой (Рп = Рл). Нагляд- ное представление о влиянии перепада потенциала на относительную транспирацию дает рис. 3.
В соответствии с рис. 3 значение перепада потенциала приближается к максимальной величине в то время, когда Т/Т0 становится меньше единицы (0.87). Эти значения для салата, овса, проса, пшеницы и житняка на суглинистой и супесчаной почвах составляют соответственно 7.7 × 102 и 7.0 × 102, 12.5 × 102 и 12.0 × 102, 16.0 × 102 и 15.0 × 102, 15.5 × 102 и 15.0 × 102 и 17.0 × 102 и 17.0 × 102 кПа. Отсюда, с увеличением степени ксероморфизма в направлении от салата к житняку и засухоустойчивым злакам начало снижения относительной транспирации существенно сдвигается в сторону больших значений перепада потенциала. При этом различие величин перепада потенциала для одного и того же растения, выращенного на суглинистой и супесчаной почвах, незначительно и находится в пределах 1.0–1.5 кПа. Интервал максимальных значений перепада потенциала соответствует интервалу относительной транспирации в пределах 0.8–0.67 (0.90– 0.70).
Существенный интерес представляет зависимость основных энергетических параметров в совокупности (потенциала влаги в почве и листьях, перепада потенциала и относительной транспи-раци 4
0 10 20 30
Влажность почвы,%
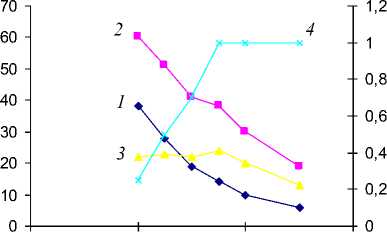
Рис. . р , перепада потенциала ( 3 ) и относительной транспирации ( 4 ) от влажности аллювиальной суглинистой почвы (пойма р. Арчеда) в условиях лабораторного опыта.
Зависимости, приведенные на рис. 4, дают наглядное представление об основных показателях энергетического состояния влаги в системе почва–растение–атмосфера и их соотношениях между собой при непрерывно уменьшающейся влажности почвы. Снижение относительной транспирации до 0.7 наблюдается при уменьшении содержания влаги в почве до 15.0% (60% НВ). Потенциал влаги в это время составил в почве 19.0 × 102, листьях 41 × 102 кПа, а перепад потенциала понизился с максимума 24 (содержание влаги 17.5% или 70% НВ) до 22 кПа. С уменьшением содержания влаги до 40% НВ относительная транспирация уменьшилась до 0.25, потенциалы влаги в почве и листьях проса возросли соответственно до (38 и 60) × 102 кПа, а перепад потенциала остался на прежнем уровне 22 × 102 кПа. Коэффициент вла-гопроводности ( К п ) в начале опыта составлял 1 × 10–10 см3/(с г), а к концу опыта уменьшился на три порядка (на рис. 3 не приведен), до 1 × 10–13 см3/(с г). Аналогичная картина наблюдается в вариантах с другими культурами на аллювиальных суглинистой и супесчаной почвах. Различие заключается лишь в абсолютных значениях Р л –Р п , Р п , К п и Т/Т 0 .
Рассмотренные функциональные зависимости в системе почва–растение–атмосфера, имеющие универсальный характер, проявляются в аналогичном виде и при проведении исследований в испарителях ГГИ-500-100, т.е. в условиях, максимально приближенных к полевым (рис. 5).
В условиях более мягкой природной атмосферной обстановки, чем в лабораторном вегетационном опыте, уменьшение относительной транспирации от 1 до 0.9 и меньше протекает постепенно, плавно, без резких перегибов зависимости Т/Т 0 от влажности. Существенное снижение относительной транспирации (с 1.0 до 0.77) осуществляется при уменьшении относительной влажности почвы до 50% НВ, что предметно свидетельствует о щадящем воздействии факторов природной среды на функционирование растений. Потенциал влаги в почве и листьях проса в это время составляет 10 × 102 и 33 × 102 кПа (перепад 23 × 102 кПа). Уменьшение Т/Т 0 до 0.50 обусловливает повышение потенциала влаги в почве до 20 × 102, в листьях проса – 35 × 102 кПа. Перепад потенциала понизился в это время до 15 × 102 кПа.
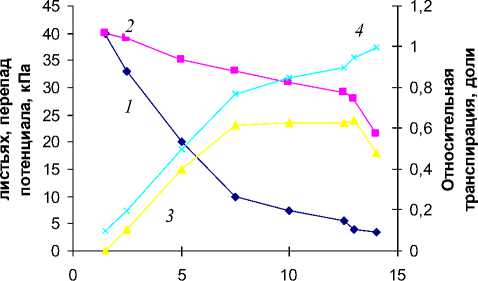
Влажность почвы,%
Рис.
репада потенциала ( 3 ) и относительной транспирации ( 4 ) от влажности аллювиальной супесчаной почвы Москворецкой поймы в испарителях.
Отметим, что с существенным понижением содержания влаги и относительной транспирации до 0.50 коэффициент влагопро-водности уменьшается с 1 × 10–9 до 1 × ×10–12 см3/(с г). С приближением содержания влаги к ВЗ перепад потенциала с оставляет 5 × 102 кПа, К п – 6 × 10–12 см3/(с г), а Т/Т 0 уменьшается до минимального значения 0.2.
Потребление почвенной влаги растениями и неусваиваемый “мертвый” запас влаги (после необратимого завядания и дальнейшей гибели культуры) определяются, как показано выше, соотношением потенциалов влаги в системе почва–растение и способностью растения развивать в листьях и корнях высокое давление клеточного сока (осмотический потенциал, сосущая сила растений). Интересные данные, в некотором роде аналогичные нашим, были получены А.Г. Дояренко (1963). По его данным, “мертвый” запас воды в почве под растениями с сосущей силой 16 атм (16 × 102 кПа) составлял в южном черноземе 14% и светлокаштановой почве 10%. При использовании растений с сосущей силой 32 атм “мертвый” запас снижается в этих почвах до 9.5 и 7.3% соответственно. В дальнейшем подобные исследования выполнены и другими учеными (Судницын, 1979; Роде, 2008; Baver, 1956; Giedrojc, Wilozynski, 1982).
Таблица 2. Соотношение потенциалов влаги в почве и листьях растений и остаточной влажности ( W ост ) в аллювиальных почвах Арчединской поймы
|
Параметр |
Гигрофиты (салат) |
Мезофиты (овес) |
Ксерофиты (просо, пшеница, житник) |
МГ* |
ВЗ = МГ × × 1.34 |
|
% |
zo |
||||
|
Аллювиальная супесчаная почва |
|||||
|
Р л , × 102 кПа |
16–18 |
40–42 |
50–65 |
1.80 |
02.41 |
|
Р п , × 102 кПа |
14–16 |
30–32 |
50–45 |
1.80 |
02.41 |
|
W ост, % |
4.0 |
2.9 |
2.1 |
1.80 |
02.41 |
|
Аллювиальная суглинистая почва |
|||||
|
Р л , × 102 кПа |
18–20 |
45–47 |
50–65 |
7.70 |
10.31 |
|
Р п , × 102 кПа |
16–18 |
35–37 |
45–47 |
7.70 |
10.31 |
|
W ост , % |
15.7 |
10.5 |
9.4 |
7.70 |
10.31 |
* МГ - максимальная гигроскопическая влага.
Под “мертвым” запасом А.Г. Дояренко понимал содержание (остаточной) влаги в почве, которое остается в ней после устойчивого завядания растений. Отметим, что снижение роста и накопление полезной продукции растениями проявляется уже при уменьшении содержания почвенной влаги до нижней границы оптимального увлажнения (около 0.7 НВ). Дальнейшее уменьшение влажности почвы приводит сначала к перманентному, а затем и к устойчивому завяданию растений. Значения влажности устойчивого завядания растений, в виде остаточной влажности, в нашем опыте содержится в табл. 2.
Величина остаточной влажности существенно различается как по почвам, так и по экологическим группам растений. Салат устойчиво завядает при содержании влаги в суглинистой почве, равной 15.7, овес-гигрофит – при 10.5, а ксерофитные растения – при 9.4%. Отсюда, салат погибает при содержании влаги в суглинистой почве, равном 1.5 ВЗ, овес-мезофит – 1.0 ВЗ, а ксерофит-ные растения – ниже 1.0 ВЗ (в интервале содержания влаги между ВЗ и МГ). Потенциалы влаги, соответствующие ВЗ, составляют в почве и растениях соответственно значения: (17 и 19) × 102 кПа в варианте с салатом, (36 и 46) × 102 кПа – овсом и (46 и 57) × 102 кПа в варианте с ксерофитными растениями.
Говоря о “мертвом” запасе почвенной влаги и влажности завядания растений, нельзя обойти проблему методов определения и оценки этой важной для гидрофизики почв и растений категории влаги, которую раньше (Судницын, 1979; Роде 2008) принято было называть константой (одной из почвенных констант).
Основной экспериментальный метод определения ВЗ (метод вегетационных миниатюр), широко используемый и в наши дни, является несовершенным и может быть использован лишь для ориентировочного представления о нижнем пределе доступности почвенной влаги растениям.
Одновременно с методом вегетационных миниатюр оценку нижней границы доступности для растений почвенной влаги (еще с середины ХХ в.) стали практиковать по потенциалу влаги. В качестве оценочного значения обычно используют 15 × 102 кПа или 15 атм (Воронин, 1984; Судницын, 1979; Baver, 1956). Однако значения потенциалов влаги в почве и растениях при влажности, соответствующей ВЗ, возрастают до величин, в 2 раза и более превышающих 15 атм. Значения Р п , Р л и Р л – Р п также существенно различаются по экологическим группам растений, что указывает на недостаточность использования в оценках ВЗ по потенциалу почвенной влаги лишь одного его значения.
Поскольку перепад потенциала влаги в системе почва– растение является движущей силой по переносу влаги и определяет нижнюю границу доступной для растений влаги, то его значения могут быть положены в основу методики оценки ВЗ, наряду с использованием для этих целей потенциалов влаги одновременно в почве и растениях.
Сущность этого предложения заключается в одновременном определении полного потенциала влаги в почве и листьях растений непосредственно в поле или в лабораторных условиях психрометрическим или криоскопическим методами. В тех случаях, когда приборы установлены в лаборатории, расположенной недалеко от сельскохозяйственных полей, пробы почв и растений можно быстро доставлять в специальных эксикаторах-термостатах, выполненных из материалов с низкой теплопроводностью, например, из тефлона. Определение потенциалов следует проводить периодически с некоторым интервалом времени и обязательно при появлении первых признаков завядания: подвядание, скручивание листьев, потеря тургора, ярко-зеленого цвета, появление желтоватого оттенка и некоторых других признаков.
Поэтому дальнейшие исследования этой проблемы должны быть направлены на отработку технологических приемов и техники определения потенциалов влаги в различных природных условиях и на разработку конкретных приемов и градаций энергетических показателей применительно к конкретным условиям среды обитания растений.
ЗАКЛЮЧЕНИЕ
Каждая экологическая группа растений (гигрофиты, мезофиты и ксерофиты) характеризуется определенным узким оптимальным интервалом потенциала почвенной влаги, при котором условия водного питания растений таковы, что значения Т/Т 0 не опускаются ниже оптимального уровня (1.0–0.9). Эти интервалы учитывают как влияние свойств почв (через потенциал влаги), так и атмосферные условия (через транспирацию) и могут быть использованы в качестве оптимального интервала влагообеспечен-ности растений.
По интенсивности снижения относительной транспирации до уровня <1–0.9 растения можно расположить в ряд: салат (гигрофит) – овес (мезофит) – ксерофитные растения (пшеница– просо–житняк), свидетельствующий о увеличении засухоустойчивости и возрастающих возможностях потребления влаги от начала ряда к его концу.
Потенциал почвенной влаги, соответствующий влажности устойчивого завядания растений, различен у растений разных экологических групп. Салат погибает при содержании влаги, равном 1.5 ВЗ, овес-мезофит – 1.0 ВЗ, а ксерофитные растения – ниже 1.0 ВЗ. Потенциалы влаги, соответствующие ВЗ, составляют в почве и растениях соответственно значения: (17 и 19) × 102 кПа в варианте с салатом, (36 и 46) × 102 кПа – овсом и (46–57) × 102кПа в варианте с ксерофитными растениями.
Интервал потребляемой влаги увеличивается, а “мертвый” запас уменьшается в направлении от гигрофитов к мезофитам и далее к ксерофитам. Уменьшение роста и накопление полезной продукции растениями проявляется уже при снижении содержа- ния почвенной влаги до нижней границы оптимального увлажнения. Дальнейшее уменьшение влажности почвы приводит сначала к перманентному, а затем и к устойчивому завяданию растений.
Основной экспериментальный метод определения ВЗ (метод вегетационных миниатюр), используемый в широких масштабах и в наши дни, является несовершенным и может быть использован лишь для ориентировочных представлений о нижнем пределе доступности почвенной влаги растениям.
Поскольку перепад потенциала влаги в системе почва– растение является движущей силой по переносу влаги и определяет нижнюю границу доступной для растений влаги, то его значения могут быть положены в основу методики оценки влажности завядания (наряду с использованием для этих целей потенциалов влаги в почве и растении).
Список литературы Особенности влагопотребления и влагообеспеченности растений различных экологических групп
- Воронин А.Д. Структурно-функциональная гидрофизика почв. М.: Изд-во Моск. ун-та, 1984. 204 с.
- Глобус А.М. Экспериментальная гидрофизика почв. Л.: Гидрометеоиздат, 1968. 272 с.
- Дояренко А.Г. рF почв Юго-востока и сосущая сила саратовских пшениц. Избр. соч. М., 1963. Т. 1. С. 178-186.
- Мелиоративная энциклопедия. М.: Росинформагротех, 2004. Т. 3. С. 10-11.
- Муромцев Н.А. Оценка влагообеспеченности растений//Бюл. Почв. ин-та им. В.В. Докучаева. 2011. Вып. 67. С.20-31.
- Муромцев Н.А. Определение коэффициента влагопроводности в почвенных колонах и лизиметрах//Бюл. Почв. ин-та им. В.В. Докучаева. 2013. Вып. 72. С. 84-95.
- Муромцев Н.А. Формирование и состояние влаги в капиллярной кайме дерново-подзолистой почвы при восходящем потоке из грунтовых вод//Бюл. Почв. ин-та им. В.В. Докучаева. 2005. Вып. 57. С. 50-57.
- Муромцев Н.А. Энергетические аспекты потребления почвенной влаги растениями//Бюл. Почв. ин-та им. В.В. Докучаева. 2002. Вып. 56. С. 9-12.
- Муромцев Н.А., Семенов Н.А., Мажайский Ю.А., Анисимов К.Б. Закономерности накопления, потерь и возврата влаги и химических веществ при внутрипочвенном влагообмене//Бюл. Почв. ин-та им. В.В. Докучаева. 2014. Вып. 76. С.1 11-125.
- Роде А.А. Основы учения о почвенной влаге. Избр. тр. М., 2008. Т. 3. 663 с.
- Семенов Н.А., Муромцев Н.А., Сабитов Г.А., Коротков Б.И. Лизиметрические исследования в луговодстве. М.: Аверс Пресс, 2005. 584 с.
- Судницын И.И. Движение почвенной влаги и водопотребление растений. М.: Изд-во Моск. ун-та, 1979. 254 с.
- Giedrojc В., Wilozynski А. The influence of meteorological fасtors and capillary potential of soil on field consumption by crops//Zesz. Probl. Post. nauk rol. 1982. V. 93. No. 281. P. 85.
- Hosseini S.M., Mir M., Ganjian N., Pisheh Y.P. Estimation of the water retention curve for unsaturated clay//Can. J. Soil Sci. 2011. V. 91. No. 4. P. 543-549.
- Pertovici T., Marinov A.M. A mathematical generalized approach to estimate soil moisture retention characteristics from texture classes//Univ. Politehn. Bucharest. Sci. Bull. D. 2010. V. 72. No. 1. P. 59-66.
